

Экология, 2020, № 1, стр. 16-24
Влияние факторов среды на разнообразие жизненных форм травянистой растительности юга лесостепи
П. А. Шарый a, *, Л. С. Шарая b, Л. В. Сидякина b, С. В. Саксонов b
a Институт физико-химических и биологических проблем почвоведения РАН
142290 Московская обл., Пущино, ул. Институтская, 2, Россия
b Институт экологии Волжского бассейна РАН
445003 Самарская область, Tольятти, ул. Комзина, 10, Россия
* E-mail: p_shary@mail.ru
Поступила в редакцию 06.03.2019
После доработки 20.05.2019
Принята к публикации 29.05.2019
Аннотация
Для понимания закономерностей изменения видового богатства (ВБ) на охраняемых природных территориях необходимо знание влиятельных факторов среды. В Национальном парке “Самарская Лука” для доминирующих жизненных форм (ЖФ), а именно гемикриптофитов и геофитов, построены модели регрессии видового богатства, выявляющие солнечную энергию и рельеф как ведущие факторы. Карты ВБ показывают разделение ЖФ в пространстве, уменьшающее конкуренцию между ними. Обнаружено, что доля ВБ терофитов в редколесье на участке в 3 раза меньше, чем в Среднем Поволжье, что отражает влияние локального видового пула лесного сообщества. Спектр ВБ ЖФ следует геометрической прогрессии и связан с активными геологическими разломами.
Изучение закономерностей пространственного распределения видового богатства (ВБ) сосудистых растений основано прежде всего на выявлении их связей с такими факторами среды, как климат и энергия солнечной радиации. В глобальном масштабе (разрешение ~100 км) энергию оценивают потенциальной эвапотранспирацией [1]. В данном масштабе связи обычно наиболее тесны из-за больших диапазонов изменения как ВБ, так и климатических условий [2], что позволяет объяснять более 80% дисперсии ВБ растений [1, 3] и строить глобальные карты их ВБ [4, 5]. Подобные карты рассчитывают и для отдельных групп растений, как, например, папоротникообразные [6].
Аналогичные факторы среды используют и в региональном масштабе [7, 8], где разрешение ~10 км. Из-за меньшего диапазона изменений климата связи с факторами среды в этом масштабе обычно слабее, объясняя, например, 50% дисперсии ВБ в Австрийских Альпах [9]. При раздельном изучении групп растений в горах и предгорьях Калифорнии [7] данный показатель менялся от 62% (кустарники) до 88% (многолетние травы).
При разрешении 10–100 м в локальном масштабе климат не учитывают из-за его малых изменений на участке исследования, но при этом появляется возможность детально описывать специфику местообитаний: сомкнутость крон деревьев [10], рельеф [11], менеджмент [12], свойства почв [13] и освещенность склонов как характеристику энергии [14].
В локальном и региональном масштабах информативно также деление растительности на группы, поскольку разные группы растений по-разному адаптированы к факторам среды [7]. Наиболее известно деление сосудистых растений на жизненные формы (ЖФ) по простому, но фундаментальному принципу – положению почек возобновления относительно поверхности почвы, показывающему их защищенность от неблагоприятного периода (морозов и/или засух). К. Раункиер [15] выделил пять основных ЖФ с возрастающей защищенностью: фанерофиты (почки возобновления выше 25 см над землей), хамефиты (над землей, но ниже 25 см), гемикриптофиты (на почве), криптофиты (под землей и/или под водой) и терофиты (возобновление только из семян: однолетние травы). Он также предложил использовать спектры ЖФ, т.е. доли ВБ каждой ЖФ (в %). В спектрах ЖФ фанерофиты имеют наибольшую долю во влажных тропиках (~96%), гемикриптофиты – в умеренном климате (~63%), терофиты – в пустынях (~73%) [16], криптофиты могут преобладать в прибрежно-водных регионах степной зоны (~69%) [17].
Распределение долей ВБ различных ЖФ обычно удовлетворяет следующей закономерности: если доминирующая ЖФ занимает долю k объема экологической ниши сообщества, то следующая по числу видов ЖФ занимает примерно такую же долю от оставшейся части ниши (1 – k), следующая – от (1 – k)2 и т.д. Это дает геометрическую прогрессию pi = p1(1 – k)i– 1, где pi – доля ВБ ЖФ ранга i, а p1 – доля ВБ доминирующей ЖФ. Впервые данную закономерность обнаружил И. Мотомура [18] (см. также [19]) при изучении числа особей видов моллюсков, губок и т.д. в приливно-отливной зоне. Это явление известно как захват ниши [20] и часто выполняется для ВБ ЖФ сосудистых растений [21]. Показатель прогрессии d = 1 – k характеризует спектр ЖФ и отражает их доминирование: чем меньше d, тем оно сильнее. Однако при использовании вместо ВБ характеристик обилия растений распределение может отличаться от геометрической прогрессии [21].
Полагают, что главными факторами среды, влияющими на ВБ растений, являются свет, влага, pH и питательные вещества почв [22]. Известна гипотеза, что ВБ зависит от локального или регионального видового пула [22]. В локальном масштабе часто важен микроклимат, который создается рельефом и различен на северных и южных склонах. На изучаемом нами участке для травянистой растительности под пологом леса главными факторами среды были свет и влага [14]. ВБ обычно уменьшается с ростом сомкнутости крон для травянистой растительности под пологом леса [23]. Однако и сама сомкнутость крон зависит от доступного деревьям света [14].
Для изучения распределения солнечной энергии склонами в локальном масштабе иногда используют взятые порознь экспозицию и крутизну склонов [10, 24]. Однако они действуют в определенном сочетании, которое описывает перпендикулярность падения солнечных лучей на земную поверхность. Это сочетание, относительная освещенность склонов [25], характеризует энергию в локальном [14] или региональном [26] масштабах.
Для изучаемого нами участка было показано [14], что сомкнутость крон 15% является критической: при меньшей сомкнутости лимитирующим фактором становится недостаток влаги, при большей – света. Найдена тесная (R2 = 0.77) связь ВБ с освещенностью склонов и другими факторами среды и построены карты ВБ, pH почв, сомкнутости полога и зеленой массы травянистых растений [14]. Однако роль ЖФ в разнообразии растительности в литературе не изучалось. Полезно сравнить реакции ЖФ на действие факторов, поскольку они имеют различные механизмы адаптации к условиям среды. Важно также сравнить влияние на ВБ ЖФ локального и регионального видовых пулов. Цель данной работы – сравнительное изучение действия факторов среды в локальном масштабе на ВБ и обилие разных ЖФ, а также сравнение влияния на ВБ ЖФ локального и регионального видовых пулов.
МАТЕРИАЛ И МЕТОДЫ
Карта участка исследования с указанным на ней положением точек наблюдения показана на рис. 1. Участок (382 га) находится на невысокой (265 м) горе Могутовой (Самарская Лука) в Самарской области (Среднее Поволжье) и на 79% покрыт лесом. Здесь, на южной границе лесостепи, климат полуаридный и континентальный: средняя температура января –10°С, июля +20°С, среднегодовые осадки 566 мм, температура +4.8°С. Почвы участка в основном дерново-карбонатные, в местах выхода карбонатов – слаборазвитые литосоли или немногим более мощные рендзины.
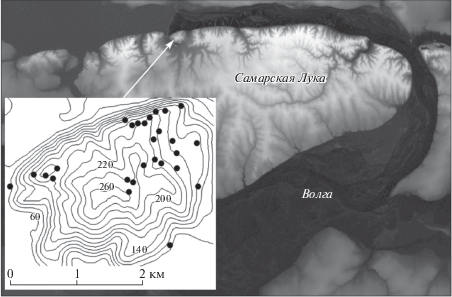
Полевые измерения. На участке выбраны 27 площадок: 23 представляют лесные сообщества и 4 – сообщества луговой и каменистой степи. На каждой из них, размером 20 × 20 м для лесных и 10 × 10 м для степных сообществ, случайно отбирали 4 квадрата со стороной 0.5 м. На площадках с конца июля до начала августа подсчитывали число видов растений травяного яруса, взвешивали сухую зеленую массу, визуально оценивали сомкнутость полога. Растения собирали однократно, делили на виды и высушивали до воздушно-сухого состояния. Всего было найдено 138 видов. Влажность почв измеряли в слое 0–10 см как описано в работе [14].
Освещенность и экспозиция склонов. Перпендикулярность падения солнечных лучей на земную поверхность описывается относительной освещенностью склонов F(a,b), зависящей от двух углов: склонения Солнца a над горизонтом и азимута Солнца b [25]. Ее можно выражать в процентах (100% для перпендикулярного падения лучей и 0% – для теневых склонов) или в единицах энергии (Вт/м2). Число пасмурных дней летом на юге лесостепи невелико, и рассеянное излучение играет небольшую роль, лишь снижая контраст освещенности склонов [27]. Угол a брали равным 35°, а угол азимута b изменяли так, чтобы он отвечал максимально тесной связи с изучаемым свойством фитоценоза, т.е. в процессе создания моделей (уравнений регрессии) тестировали различные значения азимута b (от восточного через южный до западного через каждые 5°) и выбирали тот из них, для которого коэффициент детерминации является наибольшим.
Проникающую под полог леса солнечную радиацию оценивали как Fg(a,b) = F(a,b)[1 – (Сомкнутость/100)] [14]. Измерения люксметром в этом случае неадекватны, так как они отражают только сомкнутость, не учитывая крутизну и экспозицию склонов. Для степных сообществ Fg(a,b) совпадает с F(a,b), уменьшаясь в лесу с ростом сомкнутости полога.
Экспозицию (A0) преобразовывали в нециклические переменные, а именно sin A0, cos A0, sin A45 и cos A45, где A45 = A0 + 45° [28]. На основании статистических сравнений выбирали ту из них, для которой связь с изучаемым свойством фитоценоза была наиболее тесной. Влияние экспозиции крутых и пологих склонов на данный показатель учитывали с помощью произведения или частного функции на крутизну GA [29]: GA × cos A0 или cos A0/GA.
Другие характеристики рельефа. Для описания рельефа использовали систему 18 морфометрических величин (МВ), описанную в [25]. Все МВ рассчитывали по цифровой модели рельефа ASTER (Advanced Spaceborne Thermal Emission and Reflection Radiometer), преобразованной в проекцию Гаусса-Крюгера для 9-й зоны с разрешением 40 м.
Жизненные формы. Для травяного яруса подсчитывали ВБ и сухую зеленую массу пяти ЖФ: фанерофитов, хамефитов, гемикриптофитов, геофитов (почки возобновления под землей) и терофитов. Фанерофиты верхних ярусов не учитывали, а из фанерофитов и хамефитов в травяном ярусе учитывали только растения высотой до 15 см. Данные о зеленой массе, влажности и pH почв, сомкнутости полога и освещенности приведены в работе [14].
Статистический анализ. Разработанная П.А. Шарым программа “Аналитическая ГИС Эко” использовалась для расчета МВ, статистического анализа и построения карт. Преобразованные для приближения их статистического распределения к нормальному МВ (см. [14]) помечены верхним индексом “T”. Проверку моделей проводили с помощью показателя деградации Degr и эмпирического критерия успешности проверки Degr < 50% [14]. Индикаторная переменная ЛЕС служила для различения лесных и степных сообществ: 1 – лес, 0 – степь. Ниже в моделях регрессии предикторы расположены в порядке убывания статистической значимости. Оценивали независимость предикторов, и любую комбинацию зависимых предикторов исключали из рассмотрения. Карты строили по уравнениям регрессии путем расчета зависимой переменной для всех пикселей.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Из 27 площадок лесных и степных сообществ травянистых растений горы Могутовой 18 занимали широколиственные леса: кленовники (Acer platanoides), липняки (Tilia cordata) и ильмовники (Ulmus laevis). Эти леса располагались на пологих склонах верхней части горы, ориентированных преимущественно на север и северо-восток. Средняя сомкнутость древесного полога широколиственных лесов составляла 60%, почва здесь была наиболее увлажненной (средняя влажность за сезон измерений 30.4%). Площадки с этими лесами также характеризовались наиболее высоким средним значением топографического индекса влажности и самым низким значением индекса мощности потоков. Пять площадок были представлены сосняками (Pinus sylvestris, P. cretacea), березняками (Betula pendula) и осинниками (Populus tremula), которые имели среднюю сомкнутость 20%. На горе они располагаются ниже широколиственных лесов на крутых склонах разной экспозиции. Средняя влажность почв в этих редкостойных лесах 23.5%, площадки имеют более низкое значение топографического индекса влажности и заметно более высокое – индекса мощности потоков. Площадки луговых и каменистых степей расположены в самом нижнем по высоте ярусе, преимущественно на южных относительно крутых склонах. Средняя влажность почв на степных площадках наиболее низкая (16.7%), минимально также значение топографического индекса влажности.
Травяной покров на площадках широколиственных лесов имеет относительно высокое среднее значение общего проективного покрытия (ОПП) – 62%, наиболее низкое – числа видов (9.1) и зеленой фитомассы (86.3 г/м2). Для редкостойных лесов среднее значение ОПП меньше – 46%, число видов в 1.5 раза больше – 14.4, зеленой фитомассы в 2.5 раза больше – 203.8 г/м2. Степные площадки характеризовались наиболее высокими средними значениями ОПП – 65%, числа видов – 23, зеленой фитомассы – 229.5 г/м2.
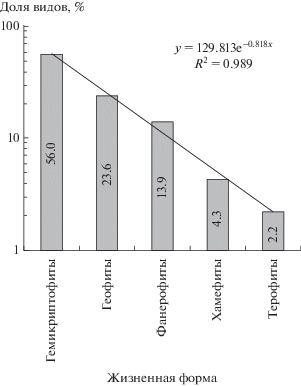
Представленный на рис. 2 спектр ЖФ исследованного участка свидетельствует о преобладании гемикриптофитов, что обычно для умеренного климата. Экспоненциальный тренд для спектра в полулогарифмических координатах есть прямая, поэтому доли ВБ ЖФ убывают в геометрической прогрессии, т.е. по закону $y = 129.8 \times {{d}^{x}},$ где показатель прогрессии d = 0.441, а x есть ранг ЖФ (от 1 до 5), упорядоченных по убыванию их долей (d = e–b при использовании тренда y = ae–bx; на рис. 2 – a = 129.813, b = 0.818). Показатель d характеризует влагообеспеченность площадок: чем меньше d, тем ниже влагообеспеченность. Рассчитанные нами по данным [16] значения d возрастают в ряду сухой злаковник (0.422) – дубовое редколесье (0.573) – лес умеренно холодной зоны (0.630). Близкое к нашему значение d = 0.459 найдено для спектра ЖФ Среднего Поволжья до данным [30], однако доля терофитов для этого спектра (17%) выше обнаруженной нами (2.2%, см. рис. 2). Это объясняется малым покрытием Среднего Поволжья лесами, в то время как на нашем участке 79% площади занято лесными сообществами со средней сомкнутостью крон 42%.
Зеленая масса ЖФ также убывала в геометрической прогрессии, т.е. по закону $y = 135.9 \times {{d}^{x}},$ где d = 0.400. Меньшая величина d означает, что зеленые массы ЖФ уменьшались более резко, чем доли ВБ. Зеленая масса терофитов составляла 1.5% от общей зеленой массы на площадках.
Доли видового богатства ЖФ не независимы друг от друга. На участке это проявляется в отрицательной связи между долями ВБ доминирующих ЖФ – гемикриптофитов и геофитов (R2 = 0.789, P < 10–6). Сравнительно редкие ЖФ из-за быстрого убывания их долей в геометрической прогрессии часто отсутствуют на площадках; для их представления удобен графический способ. Некоторые из закономерностей их распределения на площадках показаны на рис. 3.
Рис. 3.
Гистограмма долей жизненных форм на площадках. Столбцы упорядочены сначала по убыванию (слева направо) доли терофитов, а далее, где эти доли равны нулю, – по убыванию (слева направо) освещенности под пологом с юго-запада.
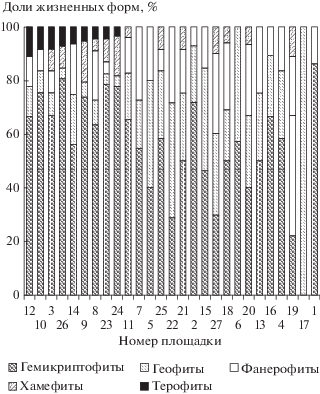
Терофиты встречались только на 4 площадках степных сообществ и на 4 с редкостойными лесами (березняками, осинником и сосняком). Площадки с терофитами характеризовались низкими значениями сомкнутости полога и влажности почв, а также высокой освещенностью с юго-запада под пологом. Такие площадки с терофитами назовем сухими. Найденное ранее критическое значение сомкнутости крон совпадает со средним для сухих площадок: для сомкнутости крон ниже 15% лимитирующий фактор – недостаток влаги, выше – света [14]. Терофиты адаптированы к недостатку влаги, но в меньшей степени – к затенению. Их присутствие в сообществе связано с обилием света и сухими условиями. В более затененных и влажных условиях их заменяют другие ЖФ.
Помимо терофитов, на сухих площадках увеличивались доли хамефитов и гемикриптофитов в 2 и 1.5 раза соответственно, в то время как доля геофитов снижалась почти в 4 раза (табл. 1). Показатели обилия ЖФ претерпевали более резкие изменения: зеленая масса хамефитов на них была почти в 12 раз выше, чем на остальных, а зеленая масса геофитов и фанерофитов, напротив, в 2 раза меньше (см. табл. 1). Отношение ВБ гемикриптофитов и геофитов возрастало на сухих площадках почти шестикратно. Поэтому для ВБ гемикриптофитов и геофитов можно было ожидать не только различных связей с факторами среды, но и противоположных.
Таблица 1.
Значение средних характеристик травяного покрова для разных групп фитоценозов
| Группа фитоценозов |
Зеленая масса травяного покрова, г/м2 | Влажность почв, % | ||||
|---|---|---|---|---|---|---|
| гемикриптофиты | геофиты | фанерофиты | хамефиты | терофиты | ||
| Разреженные леса* и степи | 164.7 | 15.2 | 4.0 | 18.3 | 6.0 | 19.7 |
| Широколиственные леса | 50.1 | 32.1 | 6.2 | 1.5 | 0 | 30.8 |
| Их отношение | 3.29 | 0.47 | 0.64 | 12.5 | ∞ | 0.64 |
| Группа фитоценозов | Доля видового богатства, % | Освещен- ность**, % | ||||
| гемикриптофиты | геофиты | фанерофиты | хамефиты | терофиты | ||
| Разреженные леса и степи | 71.0 | 8.1 | 7.7 | 6.7 | 6.6 | 40.4 |
| Широколиственные леса | 48.6 | 31.3 | 17.1 | 3.1 | 0 | 23.0 |
| Их отношение | 1.46 | 0.26 | 0.45 | 2.18 | ∞ | 1.76 |
Связь доли ВБ гемикриптофитов Nsh% с долей ВБ геофитов Nsg% и факторами среды выражалась уравнением регрессии
(1)
$\begin{gathered} Nsh\% = --0.673Nsg\% --415ЛЕС{{({{{\text{sin}}{{A}_{{45}}}} \mathord{\left/ {\vphantom {{{\text{sin}}{{A}_{{45}}}} {GA}}} \right. \kern-0em} {GA}})}^{2}} + \\ + \,\,27.4K{{R}^{{\text{T}}}} + 4.04 \times {{10}^{{--5}}}Fg{{90}^{2}} + 48.4; \\ {{R}^{2}} = 0.904\,(Degr = 3.6\% ),\,\,\,\,P < {{10}^{{--6}}}. \\ \end{gathered} $(2)
$\begin{gathered} Nsh\% = --0.128ЛЕС \times Z + 30.0M_{{ + 2.43}}^{{\text{T}}} - \\ --\,\,475ЛЕС{{({{{\text{sin}}{{A}_{{45}}}} \mathord{\left/ {\vphantom {{{\text{sin}}{{A}_{{45}}}} {GA}}} \right. \kern-0em} {GA}})}^{2}} + {\text{ }}11.7kh{{e}^{{\text{T}}}} + {\text{ }}40.5; \\ {{R}^{2}} = 0.744\,(Degr = 10\% ),\,\,\,\,P < {{10}^{{--5}}}; \\ \end{gathered} $(3)
$\begin{gathered} Nsg\% = 0.192Z - 5.95{{\left( {{{H}^{{\text{T}}}}} \right)}^{2}} - \\ - \,\,0.0426Fg(35^\circ ,245^\circ ) + 9.41kmi{{n}^{{\text{T}}}} + 16.5; \\ {{R}^{2}} = 0.646\,(Degr = 47\% ),\,\,\,\,P < {{10}^{{--3}}}. \\ \end{gathered} $Согласно модели (2), доля гемикриптофитов уменьшается на пологих северо-восточных и юго-западных склонах (предиктор (sin A45/GA)2), в то же время из парных связей известно, что доля геофитов Nsg% на таких склонах возрастает. С другой стороны, согласно модели (3), на хорошо освещенных под пологом юго-западных склонах (предиктор Fg(35°, 245°)) доля ВБ гемикриптофитов заметно растет. Пространственное разделение этих двух доминирующих ЖФ происходит не только под влиянием солнечной радиации, но и высоты Z: из (2) и (3) видно, что доли имеют разные знаки связи с Z, так что на меньших высотах преобладает доля ВБ гемикриптофитов, на бóльших – геофитов.
Уравнения (2) и (3) включают также другие характеристики рельефа: несферичность M, принимающую нулевые значения на округлых формах рельефа и положительные – на вытянутых (водоразделах и долинах); квадрат средней кривизны H 2, также увеличивающийся на вытянутых формах рельефа; минимальную кривизну kmin, принимающую большие по модулю отрицательные значения в доньях долин, и избыточную горизонтальную кривизну khe, описывающую расчлененность рельефа [25]. Эти МВ указывают на приуроченность долей ВБ гемикриптофитов и геофитов к вытянутым формам рельефа, которые связаны с известной геологической структурой горы Могутовой. Именно ее в направлении с юго-востока на северо-запад пересекают активные разломы, выражающиеся в рельефе параллельными долинами и гребнями в данном направлении [31].
Карты долей ВБ гемикриптофитов и геофитов, рассчитанные по уравнениям (2) и (3), представлены на рис. 4, где эти геологические структуры проявляются в заметных изменениях значений долей ВБ доминирующих ЖФ в области расположения разломов. Кроме того, доля гемикриптофитов возрастает в степных сообществах и вблизи них, уменьшаясь под лесом, а ВБ геофитов демонстрирует меньшую зависимость от недостатка света. Противоположные реакции двух доминирующих ЖФ на изменение одних и тех же условий среды потенциально способствуют снижению конкуренции между ними.
Рис. 4.
Карты долей видового богатства гемикриптофитов (а) и геофитов (б), построенные по моделям (2) и (3) соответственно.

Рассмотрим, как изменяются зеленая масса и доли ВБ различных ЖФ при сомкнутости крон меньше и больше 15%. Из табл. 1 видно, что значения долей ВБ и зеленой массы гемикриптофитов, хамефитов и терофитов больше при сомкнутости крон <15%; по этим же показателям фанерофитов и геофитов больше при сомкнутости >15%. Из приведенных в табл. 1 отношений можно заключить, что гемикриптофиты, хамефиты и терофиты светолюбивы, в то время как фанерофиты травяного яруса и геофиты можно характеризовать как теневыносливые ЖФ. Особенно светолюбивы терофиты и хамефиты, поскольку для гемикриптофитов отношения сравнительно малы и лежат между 1.5 по числу видов и 3.3 по зеленой массе.
Геометрическая прогрессия описывает существенное доминирование ЖФ [21], которое и наблюдается на исследуемом нами участке (см. рис. 2). Как отмечено выше, найденная для участка величина d = 0.441 лежит между значениями для сухого злаковника и дубового редколесья. Хотя 79% участка покрыто лесом, это значение d может рассматриваться как указание на низкую влагообеспеченность участка, так как он находится на южной границе лесостепи.
Если рассматривать характеристики обилия сосудистых растений вместо долей ВБ ЖФ, то распределение видов может иногда заметно отклоняться от геометрической прогрессии. В этом случае последняя обычно все еще применима в начальной стадии сукцессии, когда видов мало и отчетливо выражено их доминирование. Когда же видов становится много и их доминирование не столь отчетливо, причем действует много факторов среды, распределение может приближаться к логнормальному, а кривая зависимости от ранга вида уже не выглядеть прямой (как на рис. 2) и иметь S-образную форму [20, 21]. Однако для зеленой массы на участке этого не наблюдалось.
Как уже отмечалось, наиболее адаптированы к недостатку влаги терофиты. Однако они значительно менее теневыносливы, так что их доля ВБ составляет лишь 2% под пологом дубового леса на севере Калифорнии [23] и 2.2% на изучаемом участке. Здесь терофиты приурочены главным образом к степным сообществам и близким к ним участкам редколесья. Как показывают наши результаты, изменение ВБ и зеленой массы трав при сомкнутости 15% относится больше к терофитам, чем к гемикриптофитам. В неблагоприятные периоды от терофитов остаются только выносливые семена. Тем не менее в редколесье доля ВБ терофитов 6.6% (см. табл. 1) почти в 3 раза меньше величины 17%, характерной для мало покрытого лесами Среднего Поволжья [30]. Если принять гипотезу о связи ВБ с локальным или региональным пулом [22], то это свидетельствует о том, что для ВБ терофитов значительно важнее локальный видовой пул лесного сообщества участка, чем более отдаленный региональный пул Среднего Поволжья.
Сравнение узоров карт ВБ гемикриптофитов и геофитов (см. рис. 4) показывает, что они имеют тенденцию к разделению в пространстве, что уменьшает конкуренцию между ними. Например, терофиты полностью отсутствуют в затененных местообитаниях с сомкнутостью крон больше 15%, где средняя доля ВБ гемикриптофитов (71% при сомкнутости крон меньше 15% и 49% при сомкнутости больше 15%) все еще остается значительной, компенсируя отсутствие терофитов. Это свидетельствует о том, что гемикриптофиты более толерантны к затенению, чем терофиты, хотя зеленая масса гемикриптофитов все же уменьшается в 3.3 раза с ростом сомкнутости крон деревьев (см. табл. 1), т.е. они являются светолюбивыми. Геофиты и фанерофиты ведут себя противоположно терофитам, хамефитам и гемикриптофитам, поскольку имеют отношение при разных сомкнутостях меньше единицы как в разнообразии, так и в обилии (см. табл. 1). Такие соотношения позволяют рассматривать первые как более теневыносливые. Это ожидаемо для фанерофитов в травяном ярусе, так как они адаптированы для возобновления леса. Для геофитов, однако, механизмы адаптации к затененным местообитаниям изучены сравнительно мало.
Нестабильность местообитаний часто рассматривают в экологии как один из факторов, потенциально уменьшающих видовое разнообразие [7, 16, 21]. На рис. 4 видны резкие колебания значений долей ВБ ЖФ вблизи активных разломов. Особенно ярко эти колебания проявляются для геофитов, ВБ которых в зоне разломов местами становится близким к нулю (см. рис. 4б). Узоры представленных здесь карт показывают, что эта нестабильность имеет различное влияние на видовое богатство разных жизненных форм.
Список литературы
O’Brien E.M., Field R., Whittaker R.J. Climatic gradients in woody plant (tree and shrub) diversity: water-energy dynamics, residual variation, and topography // Oikos. 2000. V. 89. P. 588–600.
Морозова О.В. Пространственные тренды таксономического богатства флоры сосудистых растений // Биосфера. 2011. Т. 3. № 2. С. 190–207.
Currie D.J. Energy and large-scale patterns of animal- and plant-species richness // The American Naturalist. 1991. V. 137. P. 27–49.
Kier G., Mutke J., Dinerstein E. et al. Global patterns of plant diversity and floristic knowledge // J. of Biogeography. 2005. V. 32. P. 1–10.
Mutke J., Barthlott W. Patterns of vascular plant diversity at continental to global scales // Biologiske Skrifter. 2005. V. 55. P. 521–531.
Kreft K., Jetz W., Mutke J., Barthlott W. Contrasting environmental and regional effects on global pteridophyte and seed plant diversity // Ecography. 2010. V. 33. P. 408–419.
Richerson P.J., Lum K.-L. Patterns of plant species diversity in California: relation to weather and topography // The American Naturalist. 1980. V. 116. P. 504–536.
Baldwin B.G., Thornhill A.H., Freyman W.A. et al. Species richness and endemism in the native flora of California // American J. of Botany. 2017. V. 104. P. 487–501.
Moser D., Dullinger S., Englisch T. et al. Environmental determinants of vascular plant species richness in the Austrian Alps // J. of Biogeography. 2005. V. 32. P. 1117–1127.
Vetaas O.R. The effect of canopy disturbance on species richness in a central Himalayan oak forest // Plant Ecology. 1997. V. 132. P. 29–38.
Шарый П.А., Шарая Л.С., Коротков В.Н. Роль зон аккумуляции в пространственном распределении почв и эколого-ценотических групп // Лесоведение. 2015. № 5. С. 346–358.
Martín-Queller E., Diez J.M., Ibáñez I., Saura S. Effects of silviculture on native tree species richness: interactions between management, landscape context and regional climate // J. of Applied Ecology. 2013. V. 50. P. 775–785.
Шарая Л.С., Сидякина Л.В. Пространственная изменчивость зеленой массы травянистой растительности горы Могутова (Самарская Лука) // Фиторазнообразие Восточной Европы. 2018. Т. 12. № 4. С. 94–103.
Шарый П.А., Шарая Л.С., Сидякина Л.В., Саксонов С.В. Влияние солнечной энергии и сомкнутости крон деревьев на богатство видов травянистой растительности юга лесостепи // Сибирский эколог. журн. 2017. № 5. С. 539–552. [Shary P.A., Sharaya L.S., Sidyakina L.V., Saksonov S.V. Influence of solar energy and tree-crown closure on the species richness of grasses at the south of forest steppe // Contemporary Problems of Ecology. 2017. V. 10. № 5. P. 464–475. https://doi.org/10.1134/S1995425517050134]https://doi.org/10.15372./SEJ20170502
Raunkiaer C. Plant life forms. Oxford: Clarendon Press, 1937. 104 p.
Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1980. 327 с.
Айпеисова С.А. Анализ жизненных форм растений флористических комплексов Актюбинского флористического округа // Acta Biologica Sibirica. 2017. Т. 3. № 1. С. 46–51.
Motomura I. A statistical treatment of associations // Japanese J. of Zoology. 1932. V. 44. P. 379–383. (In Japanese)
Левич А.П. Структура экологических сообществ. М.: Изд-во МГУ, 1980. 182 с.
Whittaker R.H. Evolution and measurement of species diversity // Taxon. 1972. V. 21. P. 213–251.
Джиллер П. Структура сообществ и экологическая ниша. М.: Мир, 1988. 184 с.
Zobel M., van der Maarel E., Dupré C. Species pool: the concept, its determination and significance for community restoration // Applied Vegetation Science. 1998. V. 1. P. 55–66.
Whittaker R.H. Vegetation of the Siskiyou Mountains, Oregon and California // Ecological Monographs. 1960. V. 30. P. 279–338.
Gillet F., Murisier B., Buttler A. et al. Influence of tree cover on the diversity of herbaceous communities in subalpine wooded pastures // Applied Vegetation Science. 1999. V. 2. P. 47–54.
Shary P.A., Sharaya L.S., Mitusov A.V. Fundamental quantitative methods of land surface analysis // Geoderma. 2002. V. 107. P. 1–32.
Шарый П.А., Шарая Л.С. Изменение NDVI лесных экосистем Северного Кавказа как функция рельефа и климата // Лесоведение. 2014. № 5. С. 83–90. [Shary P.A., Sharaya L.S. Change of NDVI in Northern Caucasus as related to topography and climate // Contemporary Problems of Ecology. 2014. V. 7. № 7. P. 687–693. https://doi.org/10.1134/S1995425514070099]
Pierce K.B. Jr., Lookingbill T., Urban D. A simple method for estimating potential relative radiation (PRR) for landscape-scale vegetation analysis // Landscape Ecology. 2005. V. 20. P. 137–147.
Шарый П.А., Смирнов Н.С. Механизмы влияния солнечной радиации и анизотропии местности на растительность темнохвойных лесов Печоро-Илычского заповедника // Экология. 2013. № 1. С. 11–19. [Shary P.A., Smirnov N.S. Mechanisms of the effects of solar radiation and terrain anisotropy on the vegetation of dark conifer forests in the Pechora-Ilych State Biosphere Reserve // Rus. J. of Ecology. 2013. V. 44. P. 9–17.https://doi.org/10.1134/S1067413613010116]https://doi.org/10.7868/S0367059713010113
Stage A.R. An expression for the effect of aspect, slope, and habitat type on tree growth // Forest Science. 1976. V. 22. P. 457–460.
Воецкий А.Д., Колбасова Н.И. Видовое и биоморфологическое разнообразие как фактор адаптированности растительных семейств в травянистых сообществах Среднего Поволжья // Теоретические и прикладные проблемы использования, сохранения и восстановления биологического разнообразия травяных экосистем. Ставрополь: АГРУС, 2010. С. 92–93.
Могутова гора и ее окрестности. Подорожник / Под ред. Саксонова С.В., Сенатора С.А. Тольятти: Изд-во Кассандра, 2013. 134 с.
Дополнительные материалы отсутствуют.