

Экология, 2020, № 2, стр. 156-159
Онтогенетические изменения морфологии сперматозоидов рыжей полевки (Clethrionomys glareolus)
Г. Ю. Смирнов a, Ю. А. Давыдова a, *
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: davydova@ipae.uran.ru
Поступила в редакцию 15.09.2019
После доработки 20.10.2019
Принята к публикации 13.11.2019
Устойчивость популяций млекопитающих в пространстве и времени во многом определяется воспроизводством, для описания которого используют различные (от популяционных до суборганизменных) репродуктивные характеристики животных, например возраст вступления в размножение, плодовитость, продолжительность репродуктивного периода, морфофункциональное состояние репродуктивных органов и т.п. Исследования влияния различных (в том числе техногенных) факторов среды на репродуктивную сферу должны базироваться на оценках естественной изменчивости, в первую очередь онтогенетической. У мышевидных грызунов природных зон умеренного пояса можно рассматривать два аспекта онтогенетической изменчивости репродуктивных признаков: по вариантам онтогенеза и стадиям полового развития.
Первый из них связан с бивариантным развитием особей, при котором одна часть животных созревает в год рождения (I вариант), другая – на следующий год после зимовки (II вариант) [1, 2]. Реализация той или иной траектории развития зависит от времени рождения особи, состояния популяции и других факторов. Кроме продолжительности жизни (3–6 мес. у I варианта, 12–15 мес. – у II), этот тип изменчивости детерминирует различия многих признаков: метаболических [3], биоэнергетических [4], морфофизиологических [1], системы крови [5], возрастных изменений зубов [2] и др. Вариант онтогенеза также обусловливает различия в скорости роста и созревания, продолжительности репродуктивного периода и плодовитости, поэтому его учитывают при анализе динамики популяции [6–9]. Бивариантный онтогенез рассматривают как частный случай поливариантного онтогенеза, обеспечивающего перераспределение репродуктивных усилий в жизненном цикле особей [10]. Результат этого перераспределения – функциональную неоднородность популяции – отражает репродуктивно-возрастная структура, в которой обычно различают созревающих (половозрелых) и несозревающих (неполовозрелых) сеголеток и перезимовавших особей.
Второй аспект изменчивости связан со стадиями полового развития животных (неполовозрелость, созревание, половозрелость, угасание половой функции) и не зависит от варианта онтогенеза. Cамцов по комплексу экстерьерных и интерьерных признаков и дате поимки можно дифференцировать на группы неполовозрелых, созревающих, половозрелых (потенциально способных к размножению) и с угасанием репродуктивной функции, сопровождающейся инволюцией семенника. Оба аспекта изменчивости не конфликтуют между собой – всю совокупность онтогенетических (возрастных и репродуктивных) изменений можно исследовать, выделяя группы с учетом и варианта онтогенеза, и стадии полового развития.
Характеристики спермы животных – одни из наиболее важных в оценке мужской фертильности. Однако у млекопитающих из природных популяций их онтогенетическая изменчивость изучена слабо. Большинство исследований выполнено на человеке [11–13], в меньшей степени – на лабораторных и сельскохозяйственных животных [14–16], в редких случаях – на диких животных [17, 18]. Чаще всего регистрируют ухудшение качества спермы, в том числе ее морфологических характеристик, и снижение фертильности с возрастом [11, 12]. В других работах не выявлено ухудшения репродуктивных характеристик при старении [19–21].
Цель настоящей работы – оценить онтогенетическую изменчивость морфологических признаков сперматозоидов у широко распространенного вида мелких млекопитающих – рыжей полевки (Clethrionomys glareolus Schreber, 1780). Мы выясняли, нужно ли учитывать онтогенетическую изменчивость при анализе размеров нормальных сперматозоидов и частоты встречаемости клеток с дефектами.
В работе использовали материалы, полученные в ходе учетов численности мелких млекопитающих в Висимском заповеднике (2007–2017 гг., южная тайга, Средний Урал, 57°22′ с.ш., 59°46′ в.д.). Животных отлавливали с помощью трапиковых живоловок в течение весеннего (вторая половина мая–начало июня), летнего (вторая половина июля) и осеннего (конец августа–начало сентября) туров. Выборка самцов (n = 72) отражала структуру популяции в период отловов. Весной преобладали перезимовавшие особи, летом – перезимовавшие особи, половозрелые и неполовозрелые сеголетки, осенью – неполовозрелые сеголетки и сеголетки с инволюцией семенника. Созревающие сеголетки и перезимовавшие особи с инволюцией семенника практически отсутствовали, поэтому изменчивость сперматозоидов между репродуктивными стадиями рассматривали у сеголеток (половозрелых и с инволюцией семенника), а между вариантами онтогенеза – у половозрелых сеголеток и перезимовавших особей. Исследовали 7 половозрелых сеголеток (m), 9 сеголеток с инволюцией семенника (m/i) и 56 перезимовавших особей (ow). Календарный возраст определяли по возрастным изменениям зубов [2].
Для анализа морфологии сперматозоидов готовили мазковые препараты содержимого хвостовой части эпидидимиса семенника. Клетки фотографировали с помощью микроскопа и цифровой камеры (Leica Microsystems, Germany) при увеличении ×400. На изображениях различали нормальные сперматозоиды, клетки с дефектами головки и дефектами хвоста [22]. У каждого животного оценивали 200 сперматозоидов, затем подсчитывали долю аномальных клеток. У 30 нормальных сперматозоидов измеряли максимальные длину и ширину головки, длину средней и основной части хвоста.
Для анализа изменчивости размеров нормальных сперматозоидов и частоты встречаемости дефектов использовали обобщенные линейные модели (GLM) для непрерывных и дихотомических признаков (линейная и логит-регрессия). Отношения шансов (OR) и их 95%-ные доверительные интервалы (ДИ) приведены после потенцирования коэффициентов логит-регрессии – логарифмов отношения шансов (exp(lnOR) или 1/exp(lnOR)). В качестве референтной группы выбраны половозрелые сеголетки. Контроль над ожидаемой частотой ложных отклонений при множественных проверках статистических гипотез осуществляли с помощью поправки Беньямини–Йекутили (приведены скорректированные значения уровня значимости p). Статистический анализ выполняли в пакете Statistica 8.0 и среде R.
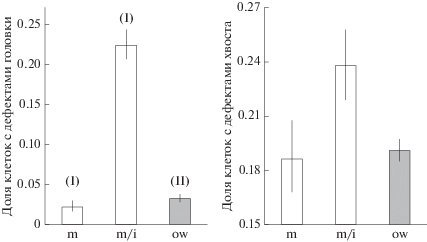
Различий морфометрических признаков сперматозоидов между репродуктивно-возрастными группами животных не обнаружено (рис. 1): для длины головки – p = 0.17, ширины головки – p = = 0.72, длины средней части хвоста – p = 0.99, основной части хвоста – p = 0.17. В то же время доля аномальных сперматозоидов различалась между группами: для дефектов головки – χ2(2) = 807.5; p < 0.0001, для дефектов хвоста – χ2(2) = 22.8; p < < 0.0001. Минимальная частота встречаемости аномальных сперматозоидов обнаружена у половозрелых сеголеток (рис. 2). Доля клеток с дефектами головки у этой группы оказалась в 4.7 (ДИ: 4.1–5.4) раза ниже по сравнению с сеголетками с инволюцией семенника (χ2(1) = 446.4; p < 0.0001) и в 1.7 (1.5–2.0) раза – с перезимовавшими (χ2(1) = 59.4; p < 0.0001). Доля сперматозоидов с дефектами хвоста у половозрелых сеголеток была ниже в 1.2 (1.1–1.3) раза, чем у сеголеток с инволюцией семенника (χ2(1) = 20.0; p < 0.0001), и в 1.1 (1.0–1.2) раза, чем у перезимовавших (χ2(1) = 6.2; p < 0.01).
Рис. 1.
Морфометрические признаки сперматозоидов (мкм, среднее и ДИ) рыжей полевки разных репродуктивно-возрастных групп: половозрелые сеголетки (m), сеголетки с инволюцией семенников (m/i), перезимовавшие самцы (ow); I, II – варианты онтогенеза.
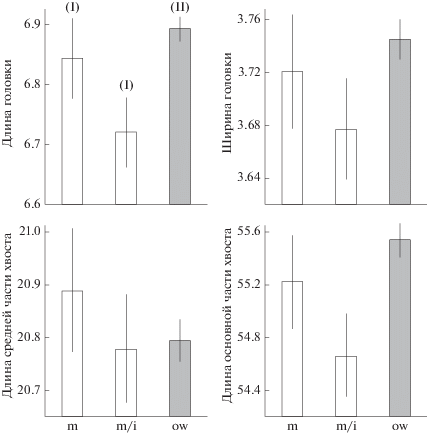
Рис. 2.
Частота встречаемости аномальных сперматозоидов (среднее и ДИ) рыжей полевки разных репродуктивно-возрастных групп: половозрелые сеголетки (m), сеголетки с инволюцией семенников (m/i), перезимовавшие самцы (ow); I, II – варианты онтогенеза.
Отсутствие различий в размерах нормальных сперматозоидов у разных репродуктивно-возрастных групп свидетельствует о консервативности половых клеток. Высокая частота встречаемости дефектов головки сперматозоидов у сеголеток с инволюцией семенника, вероятно, связана со структурно-функциональными перестройками, сопровождающими сезонную/возрастную блокировку репродуктивной функции.
Обнаруженные различия частоты сперматозоидов с дефектами головки между половозрелыми сеголетками и перезимовавшими оказались сходными со значениями, полученными нами ранее для другой выборки самцов [22], что может свидетельствовать об их закономерном характере. Можно предположить, что они обусловлены разницей в календарном возрасте животных и “накоплением” поломок. В то же время большая частота встречаемости дефектов головки у сеголеток с инволюцией по сравнению с перезимовавшими особями свидетельствует о том, что эффекты блокировки репродуктивной функции намного сильнее эффектов, связанных с календарным возрастом животных.
Таким образом, для анализа морфологии нормальных сперматозоидов не существенно, на какой стадии зрелости/старения находятся животные и когда они достигают половозрелости – в год своего рождения или после зимовки. Этот же вывод можно сделать и в отношении частоты встречаемости сперматозоидов с дефектами хвоста, поскольку обнаруженные эффекты, несмотря на статистическую значимость, оказались слабыми. Наоборот, при оценке частоты сперматозоидов с дефектами головки необходимо учитывать репродуктивно-возрастной статус животных. Кроме того, пока остается неоцененной онтогенетическая изменчивость других параметров спермы – концентрации и подвижности сперматозоидов.
Авторы выражают благодарность Е.Л. Воробейчику за обсуждение работы, анонимному рецензенту – за ценные замечания, которые позволили улучшить изложение материалов. Исследование выполнено при финансовой поддержке РФФИ в рамках научного проекта № 19-34-90004.
Список литературы
Оленев Г.В. Альтернативные типы онтогенеза цикломорфных грызунов и их роль в популяционной динамике (экологический анализ) // Экология. 2002. № 5. С. 341–350. [Olenev G.V. Alternative types of ontogeny in cyclomorphic rodents and their role in population dynamics: An ecological analysis // Rus. J. Ecol. 2002. V. 33. № 5. P. 321–330.]
Оленев Г.В. Определение возраста цикломорфных грызунов, функционально-онтогенетическая детерминированность, экологические аспекты // Экология. 2009. № 2. С. 103–115. [Olenev G.V. Determining the age of cyclomorphic rodents: Functional-ontogenetic determination, ecological aspects // Rus. J. Ecol. 2009. V. 40. № 2. P. 93–104.]
Orekhova N.A., Modorov M.V., Davydova Yu.A. Structural-functional modifications of the liver to chronic radioactive exposure in pygmy wood mouse (Apodemus uralensis) within the East-Urals Radioactive Trace // J. of Environmental Radioactivity. 2019. V. 199–200. P. 25–38.
Новиков Е.А., Кондратюк Е.Ю., Петровский Д.В. Влияние типа онтогенеза на биоэнергетические показатели красной полевки (Myodes rutilus Pall.) из горно-таежной популяции юга Западной Сибири // Экология. 2015. № 5. С. 387–391. [Novikov E.A., Kondratyuk E.Y., Petrovskii D.V. Effect of the life history pattern on bioenergetic parameters of northern red-backed voles (Myodes rutilus Pall.) in a mountain taiga population from the south of Western Siberia // Rus. J. Ecol. 2015. V. 46. № 5. P. 476–480.]
Тарахтий Э.А., Давыдова Ю.А. Сезонная изменчивость показателей системы крови рыжей полевки (Clethrionomys glareolus) разного репродуктивного состояния // Изв. РАН. Сер. Биол. 2007. № 1. С. 14–25.
Nyholm N.E.I., Meurling P. Reproduction of the bank vole, Clethrionomys glareolus, in northern and southern Sweden during several seasons and in different phases of the vole population cycle // Ecography. 1979. V. 2. № 1. P. 12–20.
Bondrup-Nielsen S., Ims R.A. Reproduction and spacing behavior of females in a peak density population of Clethrionomys glareolus // Ecography. 1986. V. 9. № 2. P. 109–112.
Löfgren O. Do intrinsic or extrinsic factors limit reproduction in cyclic populations of Clethrionomys glareolus and C. rufocanus? // Ecography. 1989. V. 12. № 1. P. 54–59.
Klemme I., Soulsbury C.D., Henttonen H. Contrasting effects of large density changes on relative testes size in fluctuating populations of sympatric vole species // Proceedings of the royal society B: Biological Sciences. 2014. V. 281. https://doi.org/10.1098/rspb.2014.1291
Новиков Е.А., Мошкин М.П. Роль стресса в модификации онтогенетических программ // Успехи современной биологии. 2009. Т. 129. № 3. С. 227–238.
Sengupta P., Nwagha U. The aging sperm: Is the male reproductive capacity ticking to biological extinction? // J. of Basic and Clinical Reproductive Sciences. 2014. V. 3. № 1. P. 1–7.
Johnson S.L., Dunleavy J., Gemmell N.J., Nakagawa S. Consistent age-dependent declines in human semen quality: a systematic review and meta-analysis // Ageing Research Rev. 2015. V. 19. P. 22–33.
Gunes S., Hekim G.N.T., Arslan M.A., Asci R. Effects of aging on the male reproductive system // J. of Assisted Reproduction and Genetics. 2016. V. 33. № 4. P. 441–454.
Beatty R.A., Mukherjee D.P. Spermatozoan characteristics in mice of different ages // J. of Reproduction and Fertility. 1963. V. 6. № 2. P. 261–268.
Martí J.I., Aparicio I.M., García-Herreros M. Head morphometric changes in cryopreserved ram spermatozoa are related to sexual maturity // Theriogenology. 2011. V. 75. № 3. P. 473–481.
Kipper B.H., Trevizan J.T., Carreira J.T. et al. Sperm morphometry and chromatin condensation in Nelore bulls of different ages and their effects on IVF // Theriogenology. 2017. V. 87. P. 154–160.
Wolf K.N., Wildt D.E., Vargas A. et al. Age-dependent changes in sperm production, semen quality, and testicular volume in the black-footed ferret (Mustela nigripes) // Biology of Reproduction. 2000. V. 63. № 1. P. 179–187.
Martínez A.F., Martínez-Pastor F., Álvarez M. et al. Sperm parameters on Iberian red deer: electroejaculation and post-mortem collection // Theriogenology. 2008. V. 70. № 2. P. 216–226.
Nieschlag E., Lammers U., Freischem C. et al. Reproductive functions in young fathers and grandfathers // The Journal of Clinical Endocrinology & Metabolism. 1982. V. 55. № 4. P. 676–681.
Rolf C., Kenkel S., Nieschlag E. Age-related disease pattern in infertile men: increasing incidence of infections in older patients // Andrologia. 2002. V. 34. № 4. P. 209–217.
Tang W.H., Jiang H., Ma L.L. et al. Relationship of sperm morphology with reproductive hormone levels in infertile men // Zhonghua nan ke xue = National J. of Andrology. 2012. V. 18. № 3. P. 243–247.
Смирнов Г.Ю., Давыдова Ю.А. Влияние промышленного загрязнения среды обитания на встречаемость аномальных сперматозоидов у рыжей полевки (Myodes glareolus) // Экология. 2018. № 5. С. 403–408. [Smirnov G.Yu., Davydova Yu.A. Effect of industrial pollution of the environment on the frequency of abnormal spermatozoa in the bank vole, Myodes glareolus // Rus. J. Ecol. 2018. V. 49. № 5. P. 459–463.]
Дополнительные материалы отсутствуют.