

Экология, 2020, № 3, стр. 190-198
Влияние низкоинтенсивного хронического облучения на репродуктивные показатели полевки-экономки (Alexandromys oeconomus): реакция родителей и потомков
О. В. Ермакова a, *, Л. А. Башлыкова a, О. В. Раскоша a, Н. Н. Старобор a
a Институт биологии Коми научного центра УрО РАН
167982 Сыктывкар, ул. Коммунистическая, 28, Россия
* E-mail: ermakova@ib.komisc.ru
Поступила в редакцию 16.02.2019
После доработки 17.07.2019
Принята к публикации 16.09.2019
Аннотация
Исследовали влияние радиационного воздействия на репродукцию полевки-экономки, населяющей территории с повышенным содержанием тяжелых естественных радионуклидов (ТЕРН). Для оценки репродуктивных показателей у животных, подвергавшихся радиационному воздействию в течение многих поколений, и их потомков проводили два эксперимента с разницей в 25 лет (в 1984 и 2009 гг.). Обнаруженная интенсификация процессов репродукции (например, увеличение доли самок и их плодовитости) свидетельствует об адаптивной реакции популяции полевки-экономки к низкоинтенсивному хроническому излучению и направлена на компенсацию высокой эмбриональной и постнатальной смертности. Однако результаты близкородственного скрещивания подтверждают наличие наследуемых генетических повреждений в популяции полевок, подвергающихся воздействию ТЕРН в среде обитания даже через 100 поколений животных.
Изучение биологических эффектов малых доз ионизирующего излучения (ИИ) на живые организмы, обитающие в природных условиях, является одной из наиболее актуальных проблем современной радиоэкологии, так как в настоящее время значительные территории подвергаются повышенному воздействию ионизирующей радиации: на месте проведения атомных взрывов, в зонах вокруг предприятий по добыче, переработке и хранению радиоактивных материалов, а также в районах аварийного радиоактивного загрязнения. Необходимость изучения адаптивных процессов в популяциях животных, позволяющих им стабильно существовать в радиоактивно загрязненной среде, не подвергается сомнению.
В последние годы принципиально изменились представления о реакциях различных биологических систем (от клетки до популяции) на воздействие ИИ в малых дозах. В отличие от ИИ в больших дозах, которое вызывает существенные клинические нарушения и гибель организма, облучение низкой интенсивности не приводит к соматической гибели организма, но оно способно модифицировать клеточные и тканевые процессы, что в конечном итоге приводит к изменению многих жизненно важных функций [1–4]. Особую значимость в условиях естественной среды обитания приобретает проблема хронического воздействия радиации на живые организмы в связи с многообразием экологических, химических и физических факторов, которые в сочетании с ИИ могут вызывать широкий спектр биологических эффектов [5–7]. Все это осложняет оценку состояния и прогнозирование ожидаемых экологических последствий для организма и популяций. Существуют и определенные методические трудности постановки экспериментов в природных условиях, необходимость поисков удобного, хорошо изученного объекта, а также сложность вычленения действия малых доз радиации в комплексе экологических факторов внешней среды.
Особенно важным для оценки возможных радиобиологических изменений, определяющих жизнеспособность, благополучие и устойчивость природных популяций мелких млекопитающих, представляет исследование интенсивности воспроизводства. Этот процесс проходит под генетическим контролем, а также под контролем нейроэндокринной и иммунной систем, и изменения в нем могут свидетельствовать о нарушениях в этих системах и геноме животных [8]. Показатели плодовитости, скорости полового созревания и величина эмбриональной смертности у мышевидных грызунов служат индикаторами популяционной приспособленности к различным неблагоприятным факторам. Изучение фолликулогенеза в яичниках позволяет оценить резервные и компенсаторно-приспособительные возможности репродуктивной функции самок в экстремальных условиях существования.
В связи с вышеизложенным представляется весьма актуальным изучение влияния хронического ИИ на репродуктивные показатели животных на основных уровнях организации (клеточный, органный, организменный и популяционный) при тех дозах и интенсивностях, которые реально существуют на загрязненных территориях.
Цель настоящей работы – оценить влияние низкоинтенсивного хронического излучения на репродуктивные показатели животных, длительно обитающих в условиях повышенного содержания тяжелых естественных радионуклидов (ТЕРН) и перемещенных в условия вивария. Для характеристики адаптивных возможностей популяции, подвергавшейся радиационному воздействию на протяжении многих поколений, репродуктивные показатели исследовали у разных поколений животных в двух экспериментах, один из которых проведен в 1980-е годы, а другой через 25 лет (т.е. по прошествии еще 30–40 поколений полевок).
МАТЕРИАЛ И МЕТОДЫ
Репродуктивные показатели исследовали у полевки-экономки (Аlexandromys oeconomus Pallas) [9], населяющей территории с повышенным содержанием ТЕРН. Вследствие широкого распространения, высокой численности, быстрой смены поколений, пониженной миграционной активности, ограниченности индивидуальных участков, доступности пищевой базы, а также тесного контакта с почвой, загрязненной радионуклидами [5], полевка-экономка может служить удобной моделью для прогнозирования последствий загрязнения природной среды, к тому же этот вид является объектом многолетнего радиоэкологического мониторинга.
Участок с повышенным радиационным фоном – “радиевый” – образовался при добыче радия из пластовых вод вследствие деятельности радиохимических заводов, которые функционировали в пос. Водный Ухтинского района Республики Коми в 1931–1956 гг. Вынос естественных радиоактивных элементов на поверхность земли с грунтовыми водами происходил на этой территории в течение многих лет. С середины XVIII в. этот процесс ускорила эксплуатация открытых здесь нефтяных месторождений, а с 1931 г. – промышленная добыча радия из пластовых вод, в которых концентрации 226Ra и 228Ra составляли соответственно 290 и 187 Бк/л [10, 11].
Мощность экспозиционной дозы на “радиевом” участке составляла 0.50–20 мкГр/ч, на “контрольном”, который находился на расстоянии 8 км, – 0.10–0.15 мкГр/ч. Геоботаническая и радиоэкологическая характеристики участков исследования описаны ранее [10].
Расчеты доз, выполненные с использованием программного комплекса ERICA Assessment Tool по данным о содержании радионуклидов в организме животных и почве [12], показали, что значения взвешенных поглощенных доз в 1970-е годы на “радиевом” участке составили 4–5 мкГр/ч и оставались на этом уровне более 30 лет, что в 10 раз и более превосходит значения мощностей доз облучения полевок, обитающих в условиях нормального радиационного фона (0.04–0.6 мкГр/ч). В этот период вклад 226Ra во внутреннее облучение полевок “радиевого” участка составлял 75.4%, остальные 24.6% приходились на 210Po и изотопы U и Th [13]. Средние значения мощности поглощенной дозы для взрослой особи варьировали в диапазоне 28–83 мГр/год (3–9 мкГр/ч) [13]. Эти значения не превышают наиболее жесткий из существующих на данном этапе рекомендуемый безопасный уровень радиационного воздействия на биоту – 10 мкГр/ч [14, 15].
Животных отлавливали с помощью живоловок [16] на “контрольном” и “радиевом” участках в период активного размножения (июль – август) в годы высокой численности популяции (1984 и 2009 гг.). Репродуктивно-возрастное состояние животных определяли по комплексу признаков (характер волосяного покрова, степень развития саггитальных межглазничных гребней черепа, состояние половой системы), на основе которых животные были разделены на неполовозрелых и половозрелых сеголеток и перезимовавших особей. Неполовозрелых сеголеток доставляли в виварий Института биологии Коми НЦ УрО РАН (http://www.ckp-rf.ru/usu/471933/) для проведения экспериментов по размножению. Животных содержали в стандартных пластиковых клетках размером 35 × 22 × 90 см со свободным доступом к воде и пище при естественном освещении и постоянном температурном режиме (18–20°С). Манипуляции с полевками проводили с соблюдением требований международных принципов Хельсинской декларации о гуманном отношении к животным для экстирпации органов [17].
Через два месяца после отлова из полевок, достигших к этому времени половозрелости, формировали пары для размножения: в первом эксперименте – 10 пар составляли “контрольную” группу и 5 пар “радиевую”, во втором – 8 и 11 пар соответственно. У каждой самки регистрировали дату рождения детенышей, их количество в помете и число пометов. Детенышей отсаживали через 20 дней после рождения. Индивидуальную интенсивность размножения определяли как отношение фактического числа пометов самки к потенциальному числу пометов, выраженное в %. Потенциальное число пометов, которое самка могла принести за свой репродуктивный период, вычисляли как частное от деления длительности всего периода размножения самки на продолжительность беременности [18]. Учитывали гибель детенышей в первые три месяца после рождения и рассчитывали показатель постнатальной гибели потомства, который характеризует частоту доминантных полулеталей, обусловливающих смертность новорожденных детенышей в ранние (до одного месяца) сроки развития [19].
Для определения уровня рецессивных леталей применяли метод близкородственного скрещивания (инбридинг). Из самцов и самок первого поколения (возраст 3 мес.) “контрольного” и “радиевого” участков были сформированы четыре группы животных разных вариантов скрещивания: контрольный/родственное, контрольный/неродственное, радиевый/родственное и радиевый/неродственное (всего 40 пар в первом эксперименте и 46 – во втором). Для исследования эмбриональной смертности проводили дополнительный эксперимент на потомках полевок F2–F4. Для преодоления барьера нескрещиваемости сибсов родственные пары подбирали от одних и тех же родителей, но из разных пометов.
На 18-й день беременности у самок в первом эксперименте определяли количество желтых тел в яичнике, число живых и мертвых эмбрионов [20]. Подсчет фолликулов разной степени зрелости и желтых тел проводили на серийных парафиновых срезах в расчете на одно поле зрения с помощью микроскопа NU-2 (Carl Zeiss). Гистологические срезы яичников изготавливали по стандартной методике с окраской гематоксилин-эозином [21].
В первом эксперименте обследовано 950 полевок-экономок, во втором – 1022. Результаты обрабатывали с помощью пакетов прикладных программ Microsoft Excel, Statistica (6), используя t‑критерий Стьюдента, критерий малых долей [22] и усовершенствованный точный метод Фишера для статистической обработки 4-польных таблиц [23].
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
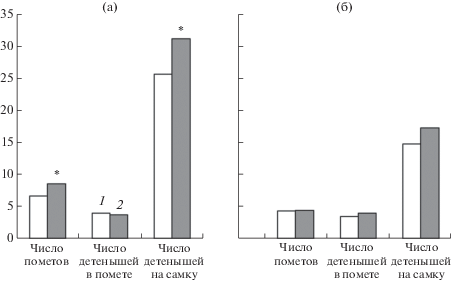
Индивидуальная интенсивность размножения в первом эксперименте (1984 г.) у полевок “радиевой” группы значимо превышала значения “контрольной” группы (81.5 и 55% соответственно). У самок “радиевой” группы продолжительность репродуктивного периода не превышала одного года, тогда как 33% самок “контрольной” группы размножались больше года [24]. Во втором эксперименте (2009 г.) значимых различий в индивидуальной интенсивности размножения не обнаружено (86.4 ± 3.9% – “радиевая” группа, 85.5 ± 3.7% – “контрольная” группа). Плодовитость самок “радиевой” группы в первом эксперименте была выше, чем в контроле (рис. 1). Во втором эксперименте значимых отличий по показателям плодовитости не обнаружено, но максимальное количество пометов, полученных от самок “радиевой” группы, – 12, а от самок “контрольной” группы – 9.
Рис. 1.
Плодовитость полевок “контрольной” (1) и “радиевой” (2) групп в первом (а) и втором (б) экспериментах; * – различия с “контрольной” группой статистически значимы при р < 0.05, уровень значимости различий вычислен с использованием усовершенствованного точного метода Фишера для статистической обработки 4-польных таблиц.
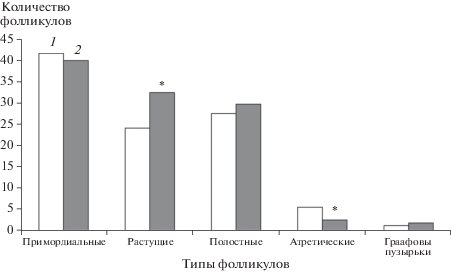
У самок, отловленных на радиевом участке в 1984 г., количество растущих фолликулов, Граафовых пузырьков и желтых тел свидетельствует о том, что потенциальная плодовитость у самок “радиевой” группы превышает наблюдаемую в “контрольной” группе, тогда как запас примордиальных фолликулов у этих животных не отличается от контроля (рис. 2). Кроме того, обнаружено снижение числа атрезирующих фолликулов.
Рис. 2.
Соотношение разных типов фолликулов в яичниках полевок-экономок “контрольной” (1) и “радиевой” (2) групп в первом эксперименте; * – различия с “контрольной” группой статистически значимы при р < 0.05.
Потомство облученных животных оказалось менее жизнеспособно. В первом эксперименте в течение первого месяца после рождения в “радиевой” группе гибнет 22.1% детенышей (в контроле – 9.4%), а до половозрелого состояния (до трехмесячного возраста) доживает 68% детенышей (в “контрольной” группе – 84%, р < 0.05). Во втором эксперименте наблюдали незначительное увеличение постнатальной смертности у полевок “радиевой” группы в первый месяц после рождения – 25.3%, в контроле – 21.8%, к возрасту трех месяцев выживаемость детенышей в “контрольной” и “радиевой” группах практически не отличается (65 и 68% соответственно). Ранняя постнатальная гибель потомства облученных родителей, рассчитанная по формуле Б.Н. Ильина [19], в первом эксперименте составляла 15%, во втором – 4.5%.
Потомство полевок-экономок отличалось и по соотношению полов. Так, в пометах полевок “радиевой” группы количество рожденных самок больше, чем у полевок “контрольной” группы (56 и 44% соответственно), во втором эксперименте эта закономерность сохранялась и составляла 58 и 44%.
Репродуктивные особенности исследовали и на потомках F1 при разных вариантах их скрещивания (табл. 1). При неродственном скрещивании в первом эксперименте в “радиевой” группе число размножающихся пар значимо превышало контроль, во втором различий не наблюдалось. Число детенышей в расчете на одну самку в “радиевой” группе в первом эксперименте при неродственном скрещивании значимо больше, чем в контроле, во втором эксперименте отличий не обнаружено (табл. 2).
Таблица 1
. Число размножающихся пар у F1 полевок-экономок, родители которых обитали на контрольном и радиевом участках
| Вариант скрещивания | Первый эксперимент | Второй эксперимент | ||||
|---|---|---|---|---|---|---|
| число пар |
пары, вступившие в размножение, % | число пар | пары, вступившие в размножение, % | |||
| lim | X ± m | lim | X ± m | |||
| Контрольный/неродственное | 10 | 15–66.7 | 34.3 ± 6.7 | 11 | 85.7–100 | 94.6 ± 3.0 |
| Контрольный/родственное | 10 | Не размножались | 9 | 44.5–100 | 86.4 ± 5.1 | |
| Радиевый/неродственное | 10 | 20–100 | 78.0 ± 5.9* | 15 | 80–100 | 93.1 ± 2.2 |
| Радиевый/родственное | 10 | 10–100 | 56.0 ± 7.0o | 11 | 45.5–85.7 | 67.9 ± 6.3*o |
Таблица 2.
Характеристика размножения F1 полевок-экономок при различных вариантах скрещивания (первый эксперимент, X ± m)
| Число | “Контрольная” группа | “Радиевая” группа | |
|---|---|---|---|
| неродственное скрещивание | неродственное скрещивание | родственное скрещивание | |
| Пометов на самку | 2.3 ± 0.29 | 3.2 ± 0.33* | 3.4 ± 0.27 |
| Детенышей в помете | 4.0 ± 0.29 | 4.8 ± 0.29* | 3.9 ± 0.28# |
| Детенышей на самку | 9.3 ± 1.97 | 15.4 ± 2.19* | 13.3 ± 2.63 |
В первом эксперименте у контрольных животных был обнаружен эффект нескрещиваемости сибсов в первом поколении, а у животных “радиевой” группы при родственном скрещивании число детенышей в помете было меньше, чем при неродственном (см. табл. 1). Отсутствие скрещивания сибсов в “контрольной” группе не позволяло сравнить уровень рецессивных мутаций “радиевой” и “контрольной” групп. Во втором эксперименте феномен нескрещиваемости сибсов в контроле не наблюдали, но при родственном скрещивании количество размножающихся пар было в 2 раза меньше, чем при неродственном, и составляло в первый месяц спаривания 44.7%. При родственном скрещивании обнаружили более низкие показатели числа пометов, количества детенышей в помете и общей плодовитости в “радиевой” группе по сравнению с контролем (табл. 3).
Таблица 3.
Характеристика размножения F1 полевок-экономок при различных вариантах скрещивания (второй эксперимент, X ± m)
| Число | “Контрольная” группа | “Радиевая” группа | ||
|---|---|---|---|---|
| неродственное скрещивание | родственное скрещивание | неродственное скрещивание | родственное скрещивание | |
| Пометов на самку | 2.9 ± 0.46 | 3.3 ± 0.62 | 3.0 ± 0.65 | 2.3 ± 0.30 |
| Детенышей в помете | 4.2 ± 0.28 | 4.3 ± 0.26 | 4.3 ± 0.27 | 3.7 ± 0.31 |
| Детенышей на самку | 12.1 ± 1.97 | 14.3 ± 2.50 | 12.8 ± 2.63 | 8.4 ± 1.19* |
Увеличение ранней постнатальной смертности у F2 полевок “радиевой” группы отмечали в первом эксперименте [18]. Во втором эксперименте смертность у этих полевок при неродственном скрещивании также была выше (в 2 раза) (табл. 4). Таким образом, уровень смертности выше и у родителей, и у потомков полевок “радиевого” участка. При родственном скрещивании в “радиевой” группе смертность в 4.2 раза выше, чем в контроле. Показатель ранней постнатальной гибели [19] для полевок F2 при неродственном скрещивании составил в первом эксперименте 21.2%, во втором – 6.5%.
Таблица 4.
Постнатальная смертность F2 полевок-экономок при различных вариантах скрещивания (второй эксперимент, X ± m)
| Вариант скрещивания | Число детенышей | Смертность (%) в возрасте, мес. | Смертность за три месяца | ||
|---|---|---|---|---|---|
| один | два | три | |||
| Контрольный/родственное | 129 | 7.8 ± 2.4 | 3.1 ± 1.57 | 5.4 ± 2.06 | 16.3 ± 3.32 |
| Контрольный/неродственное | 134 | 8.9 ± 2.4 | 0 | 3.0 ± 1.31 | 11.9 ± 2.71 |
| Радиевый/родственное | 92 | 32.6 ± 4.5* | 0 | 0 | 32.6 ± 4.58* |
| Радиевый /неродственное | 192 | 14.8 ± 2.5 | 4.7 ± 1.53 | 2.6 ± 1.15 | 22.1 ± 2.99* |
Спонтанный уровень эмбриональной гибели у потомства (F2–F4) полевок, родители которых обитали в условиях повышенного радиационного фона (“радиевая” группа), в первом эксперименте при неродственном скрещивании выше, чем в контроле. Во втором эксперименте эта закономерность сохраняется (табл. 5). При родственном скрещивании, напротив, эмбриональная смертность оказалась выше у контрольных животных. Во втором эксперименте наблюдали подобную закономерность – у потомков животных “радиевого” участка при родственном скрещивании более низкий уровень эмбриональной смертности по сравнению с “контрольным” (см. табл. 5).
Таблица 5.
Частота эмбриональной смертности у потомков полевок-экономок, родители которых обитали на контрольном и радиевом участках
| № группы | Число самок | Число | Летали, % | ||||
|---|---|---|---|---|---|---|---|
| желтых тел | эмбрионов | резорбций | до | после | суммарно | ||
| имплантации | |||||||
| Первый эксперимент (F2–F4) | |||||||
| Неродственное скрещивание | |||||||
| 1. “Контрольная” | 16 | 50 | 48 | 0 | 4.0 | 0 | 4.0 |
| 2. “Радиевая” | 40 | 178 | 167 | 3 | 4.5* | 1.8* | 6.2 |
| p1-2 | 0.046* | 0.014 | 0.095 | ||||
| Родственное скрещивание | |||||||
| 3. “Контрольная” | 14 | 56 | 46 | 5 | 8.9 | 9.8 | 17.8 |
| 4. “Радиевая” | 27 | 113 | 101 | 5 | 6.2* | 4.7* | 10.6* |
| р3-4 | 0.04 | 0.001 | 0.028 | ||||
| Второй эксперимент (F3) | |||||||
| Неродственное скрещивание | |||||||
| 5. “Контрольная’ | 9 | 41 | 37 | 1 | 7.3 | 2.6 | 9.8 |
| 6. “Радиевая” | 11 | 51 | 40 | 8 | 5.9 | 16.7* | 21.6* |
| р5-6 | 0.88 | 0.05 | 0.002 | ||||
| Родственное скрещивание | |||||||
| 7. “Контрольная” | 8 | 42 | 28 | 5 | 21.4 | 15.2 | 33.3 |
| 8. “Радиевая” | 8 | 38 | 32 | 2 | 10.5* | 5.9* | 15.7* |
| р7-8 | 0.037 | 0.037 | 0.01 | ||||
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Результаты двух экспериментов, проведенных с разницей в 25 лет, показали, что для полевок-экономок, отловленных на территориях с повышенным уровнем ТЕРН, характерны высокие показатели эмбриональной и постэмбриональной смертности, что компенсируется высокой интенсивностью размножения животных, повышением доли самок и их плодовитости. Увеличение доли самок в пометах, возможно, является компенсацией гибели детенышей в ранний постнатальный период развития.
В яичниках облученных животных на фоне снижения резервных возможностей (количества примордиальных фолликулов) происходит ускоренное созревание растущих фолликулов. Снижение в яичниках числа атрезирующих фолликулов и, следовательно, количества атретических тел, которые являются основными продуцентами эстрогенов, косвенно свидетельствует о том, что ионизирующее излучение модифицирует не только репродуктивную, но и эндокринную функцию органа [25].
Рецессивные летальные мутации могут наследоваться в гетерозиготном состоянии и вызывать повышенную эмбриональную смертность и пониженную жизнеспособность у потомков. В гомозиготном состоянии они проявляются не только в резорбции эмбрионов, но и снижении фактической плодовитости при родственном скрещивании, а также оказывают влияние на ранних стадиях жизни, увеличивая мертворождение и раннюю постнатальную смертность. Анализ репродуктивных показателей у полевок-экономок свидетельствует о повышении эффективности размножения потомков полевок “радиевой” группы. Обнаруженный нами эффект нескрещиваемости сибсов в первом поколении контрольных животных наблюдали и другие исследователи, например при размножении в условиях вивария обыкновенной полевки (Microtus arvalis Pallas) [26]. У потомков (F1) полевок “радиевой” группы барьер инбридинга отсутствовал, что, возможно, направлено на увеличение плодовитости. Известно, что количество детенышей в помете положительно коррелирует с концентрацией основного гормона беременности прогестерона и гормонов надпочечника – глюкокортикоидов [27]. Повышенная концентрация этих иммуносупрессоров, вероятно, обусловливает подавление иммунитета у самок “радиевой” группы.
Сравнение показателей размножения полевок F1 “радиевой” группы демонстрирует во втором эксперименте результаты, аналогичные первому, – уменьшение количества детенышей при родственном скрещивании. Снижение общей плодовитости при инбридинге и уменьшение доли успешно размножающихся пар наблюдали и другие исследователи [28, 29].
Известно, что смертность молодняка является одним из косвенных свидетельств увеличения генетического груза [19]. В связи с этим следует отметить высокий процент постнатальной смертности у родственных пар потомков “радиевой” группы, родители которых в течение многих поколений обитали в условиях повышенного содержания ТЕРН. Повышенную эмбриональную смертность у животных, подвергавшихся воздействию хронического облучения, наблюдали в популяциях полевок Северного Урала [30], ВУРСа [31] и в зоне аварии на ЧАЭС [7, 32].
При родственном скрещивании, напротив, выше оказалась эмбриональная смертность контрольных животных, что свидетельствует об их большей чувствительности. Такой “перевертыш” в эмбриональной смертности при родственном и неродственном скрещивании можно объяснить генетическим контролем, ответственным за успех инбридинга [33]. Это подтверждают и результаты эксперимента, приведенного выше, согласно которому не наблюдалось скрещивания первого поколения полевок контрольной популяции.
Во втором эксперименте обнаруживали такую же закономерность – у потомков животных “радиевого” стационара при родственном скрещивании отмечен более низкий уровень эмбриональной смертности по сравнению с контролем. Вероятно, у “контрольной” группы животных смертность на постимплантационной стадии эмбриогенеза при родственном скрещивании связана не столько с проявлением рецессивных летальных мутаций [34], сколько с элиминацией гомозигот, что в свою очередь способствует поддержанию гетерогенности популяций.
Снижение или отсутствие контроля за элиминацией гомозигот в “радиевой” группе при родственном скрещивании и определяет более низкий уровень эмбриональной смертности по сравнению с “контрольной” группой. Кроме того, родственное скрещивание является нетипичным для природных популяций мышевидных грызунов, и его проведение в условиях вивария действует на полевок как сильный стрессирующий фактор [35]. Таким образом, причиной различий в эмбриональной смертности “контрольной” и “радиевой” групп может быть и сингенное скрещивание, являющееся стрессирующим фактором для контрольной популяции.
ЗАКЛЮЧЕНИЕ
Результаты исследований показали, что на территориях с повышенным содержанием ТЕРН полевки-экономки характеризуются высокими показателями эмбриональной и постэмбриональной смертности. Эти внутрипопуляционные сдвиги компенсируются более высокой интенсивностью размножения, повышением плодовитости самок, что способствует поддержанию популяционного гомеостаза.
В первом эксперименте различия в показателях воспроизводства потомства полевок, отловленных в условиях нормального и повышенного радиационного фона, были наиболее выражены, чем во втором, через 25 лет. Это свидетельствует об адаптивном значении индивидуальных морфофункциональных перестроек, предоставляющих материал для последующего отбора наиболее приспособленных особей. Вероятно, воздействие радиационного и сопутствующих ему факторов на протяжении множества поколений в природной среде приводит к увеличению радиорезистентности, о чем можно судить по показателям размножения первого и второго экспериментов. В то же время эти протяженные во времени функциональные изменения ослабляют жизненные ресурсы отдельных особей, составляющих популяцию, и, являясь своеобразной “ценой адаптации”, свидетельствуют о том, что все приспособительные реакции организма обладают лишь относительной целесообразностью.
Результаты близкородственного скрещивания подтверждают наличие наследуемых генетических повреждений в популяции полевок, подвергающихся воздействию ТЕРН в среде обитания даже через 100 поколений животных. Процессы, происходящие на организменном и популяционном уровнях в ответ на хроническое воздействие ТЕРН, не обладают специфичностью и подчиняются общебиологическим закономерностям реакции живых систем на неблагоприятные условия окружающей среды. Исчисляемый миллиардами лет период полураспада многих ТЕРН обусловливает и большую длительность развертывания во времени адаптивных изменений, формирующихся в популяциях мелких млекопитающих, населяющих территории с повышенным естественным радиационным фоном.
Работа выполнена в рамках государственного задания Института биологии Коми НЦ УрО РАН (№ ГР АААА-А18-118011190102-7.)
Список литературы
Бурлакова Е.Б. Особенности действия сверхмалых доз биологически активных веществ и физических факторов низкой интенсивности // Российский химич. журн. (журнал Российского химического общества им. Д.И. Менделеева). 1999. Т. 43. № 5. С. 3–11.
Осипов А.Н. Молекулярные и цитогенетические эффекты в клетках системы крови млекопитающих при длительном воздействии низкоинтенсивного ионизирующего излучения и тяжелых металлов: Автореф. дис. … докт. биол. наук. М.: МГУ им. М.В. Ломоносова, 2004. 41 с.
Шевлюк Н.Н., Рыскулов М.Ф., Прилепина А.С. и др. Морфофункциональные особенности размножения позвоночных в условиях техногенно преобразованных ландшафтов // Альманах молодой науки. Оренбург: Оренбургский гос. мед. ун-т, 2017. № 1. С. 29–33.
Zaichkina S.I., Klokov Dmitry, Rozanova O.M., Smirnova E.N. Relationship between the duration of radiation adaptive response in bone marrow cells of mice and the dose of y-irradiation in vivo // Rus. J. of Genetics. 1999. V. 35. № 9. P. 1097–1102.
Алексахин Р.М., Архипов Н.П., Бархударов Р.М. и др. Тяжелые естественные радионуклиды в биосфере (миграция и биологическое действие на популяции и биогеоценозы / Под ред. Алексахина Р.М. М.: Наука, 1990. 368 с.
Григоркина Е.Б., Оленев Г.В., Тарасов О.В., Верхотурцев М.С. Радиоэкологические исследования на мелких млекопитающих в зоне влияния Восточно-Уральского радиоактивного следа: некоторые итоги // Вопросы радиационной безопасности. Спецвыпуск. 2013. № 4. С. 14–24.
Рябоконь Н.И. Сравнительный анализ влияния радиационного загрязнения на популяции двух видов мышевидных грызунов // Тр. Белорусского общества генетиков и селекционеров. Горки, 1992. С. 16.
Чернявский Ф.Б., Ткачёв А.В. Популяционные циклы леммингов в Арктике. Экологические и эндокринные аспекты. М.: Наука, 1982. 164 с.
Абрамсон Н.И., Лисовский А.А. Отряд Rodentia. Млекопитающие России: систематико-географический справочник / Ред. Лисовский А. А., Павлинов И.Я. // Сборник трудов Зоологического музея МГУ. Т. 52. М.: Тов-во научн. изд. КМК, 2012. С. 142–312.
Евсеева Т.И., Таскаев А.И. Водный промысел. Сыктывкар: Изд. Коми НЦ УрО РАН, 2000. 39 с.
Богоявленский Л.Н. Ухтинское месторождение радия // Докл. АН СССР. 1928. Сер. А. Вып. 14/15. С. 156.
Hosseini A., Brown J.E., Evseeva T.I. et al. Elaboration on a radiological environmental impact assessment methodology for Northern environments // Radioprotection. 2011. V. 46. № 6. P. 765–770.
Моисеев А.А., Маслов В.И., Тестов Б.В. и др. Дозовая нагрузка на мышевидных грызунов, обитающих на участках повышенной естественной радиоактивности. Москва, 1973. 30 с.
Beresford N.A., Wright S.M., Barnett C.L. et al. A case study in the Chernobyl zone. Part 2: Predicting radiation induced effects in biota // Radioprotection. 2005. V. 40. № 1. P. 299–305.
Garnier-Laplace J., Della-Vedova C., Andersson P. et al. A multi-criteria weight of evidence approach for deriving ecological benchmarks for radioactive substances // J. Radiol. Prot. 2010. V. 30. P. 215–233.
Тупикова Н.В. Изучение размножения и возрастного состава популяции мелких млекопитающих // Методы изучения природных очагов болезней человека. М.: Медицина, 1964. С. 154–191.
World Medical Association Declaration of Helsinki: Ethical Principles for Medical Research Involving Human subjects // Ums. 2002. P. 42–46.
Башлыкова Л.А. Эколого-генетические процессы в популяциях мышевидных грызунов, обитающих в условиях радиоактивных загрязнений: Автореф. дис. … канд. биол. наук. Сыктывкар, 2000. 22 с.
Ильин Б.Н., Борисова В.В., Ветух В.А. Отдаленные биологические эффекты комбинированного действия радионуклидов различной тропности. М.: Энергоатомиздат, 1991. 161 с.
McGee E.A., Hsueh A.J. Initial and cyclic recruitment of ovarian follicles // Endocrin Rev. 2000. V. 21. № 2. P. 200–214.
Волкова О.В., Елецкий Ю.К. Основы гистологии. М.: Медицина, 1982. 304 с.
Лакин Г.Ф. Биометрия. М.: Высшая школа, 1990. 352 с.
Сусликов В.И. Об уменьшении смертности млекопитающих от кишечного синдрома при частичном экранировании кроветворной системы или постлучевой трансплантации кроветворных клеток // Радиобиология. 1973. Т. 13. Вып. 6. С. 880–888.
Башлыкова Л.А., Ермакова О.В. Сравнительная характеристика популяций мышевидных грызунов из районов с повышенной радиоактивностью // Радиоэкологические исследования в зоне отчуждения Чернобыльской АЭС (к 20-летию аварии на Чернобыльской АЭС). Сыктывкар, 2006. С. 69–98. (Тр. Коми НЦ УрО РАН, № 180).
Ermakova O.V. Comparative morphological analysis of peripheral endocrine glands of small mammals inhabiting areas with high levels of radioactivity and exposed to chronic irradiation in model experiments // Biophysics. 2011. V. 56. № 1. P. 135–139.
Покровский А.В., Большаков В.Н. Экспериментальная экология полевок. М.: Наука, 1978. 148 с.
Мошкин М.П., Герлинская Л.А., Евсиков В.И. Иммунная система и реализация поведенческих стратегий размножения при паразитарных прессах // Журн. общ. биологии. 2003. Т. 64. № 1. С. 23–44.
Кокенова Г.Т. Влияние брачного подбора и длительного инбредного разведения на репродуктивные характеристики степной пеструшки (Lagurus lagurus Pallas, 1773): Дис. … канд. биол. наук. Новосибирск, ИСиЭЖ СО РАН, 2007. 121 с.
Евсиков В.И., Потапов М.А. Эволюционная экология плодовитости животных: 50 лет изучения размножения как связующего звена поколений млекопитающих // Вавиловский журнал генетики и селекции. 2011. Т. 15. № 1. С. 7–21.
Тестов Б.В. Репродукция полевок в различных радиоэкологических условиях // Радиоэкология биогеоценозов с повышенным фоном естественной радиоактивности. Сыктывкар, 1987. С. 27–36. (Тр. Коми фил. АН СССР, вып. № 81).
Ильенко А.И., Крапивко Т.П., Мажейките Р.Б. и др. Изучение влияния загрязнения 90Sr биогеоценоза на популяцию лесных мышей // Проблемы и задачи радиоэкологии животных. М.: Наука, 1980. С. 97–120.
Померанцева М.Д., Шевченко В.А., Рамайя Л.К. и др. Генетические повреждения у домовых мышей, обитающих в условиях повышенного фона радиации // Генетика. 1990. Т. 26. № 3. С. 466–473.
Евсиков В.И., Осетрова Т.В., Беляев Д.К. Генетика плодовитости животных. Сообщение IV. Эмбриональная смертность и ее влияние на плодовитость мышей линии BALB и C57BL и их реципрокных гибридов // Генетика. 1972. Т. 8. № 2. С. 55–66.
Семенов Х., Уолкер Д. Роль рецессивных летальных генов в спонтанной эмбриональной смертности в популяциях нелинейных мышей и крыс в условиях острова Куба // Бюл. эксперим. биологии и медицины. 1983. № 3. С. 71–73.
Беляев Д.К. Эволюционная генетика. Межвуз сб. Л., 1982. С. 35–59.
Дополнительные материалы отсутствуют.