

Экология, 2020, № 3, стр. 236-240
Морфологическая дифференциация синиц (Parus L.) в лесостепном Зауралье и правило Хатчинсона
А. В. Гилёв a, *, В. В. Тарасов a, А. Г. Ляхов a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: gilev@ipae.uran.ru
Поступила в редакцию 14.11.2018
После доработки 28.10.2019
Принята к публикации 06.11.2019
Дифференциация экологических ниш у совместно обитающих близкородственных видов – одна из интереснейших проблем современной экологии [1, 2]. Популярным объектом таких исследований стали совместно гнездящиеся синицы рода Parus L., образующие в зимний период смешанные стаи, в которые могут входить и другие виды [3–9].
С точки зрения экологии сообщество синиц безусловно является гильдией – группой видов, использующих ресурс функционально сходным образом [2]. Гильдия может состоять из родственных видов, как в данном случае, или из видов разных систематических групп [1]. Гильдия служит ареной наиболее интенсивных межвидовых отношений в сообществе. Конкурентные взаимоотношения, возникающие в гильдии, приводят к разнообразным последствиям, направленным на разделение ниш и снижение конкуренции.
Гильдия, состоящая из таксономически близких видов, представляет особый случай. Виды одного рода конкурируют между собой сильнее вследствие высокого морфологического и, следовательно, функционального сходства, и соответственно все происходящие при этом процессы проявляются более отчетливо. Кроме того, такое сообщество выступает как своеобразный единый функциональный блок в отношении других компонентов биоценоза.
Одним из наиболее простых и эффективных способов разделения экологических ниш сосуществующих видов является их морфологическая, в частности размерная, дифференциация. Дж. Хатчинсон (1959 – цит. по [2]) продемонстрировал, что отношение размеров тела у видов одного рода, входящих в одно сообщество и использующих сходный ресурс, составляет в среднем около 1.3 (от 1.1 до 1.4), а соответствующее этому отношение массы тела – 2. Эта эмпирическая закономерность, получившая название “правило Хатчинсона”, была подтверждена на многих группах живых организмов. Яркие примеры данного правила дало изучение птиц. Так, соотношение длины клювов у совместно обитающих видов, принадлежащих к одному роду, было проверено у представителей 46 семейств умеренных и тропических областей – везде оно в среднем выполнялось [2, 10]. В то же время его использование в исследованиях многовидовых сообществ неоднократно подвергалось критике [см. 2]. В настоящее время очевидно, что это правило применимо не ко всем организмам и экологическим ситуациям [2, 10]. Тем интереснее представляется изучение размерной дифференциации близких видов в различных сообществах и выявление случаев соответствия правилу Хатчинсона.
Известно, что у синиц произошло существенное разделение трофических ниш, и это уже стало хрестоматийным примером: разные виды синиц собирают пищу на разных породах деревьев, в разных ярусах леса, в разных частях кроны дерева [3–5, 11–14]. При этом нередко упоминается и их размерная дифференциация: мелкие виды собирают насекомых на тонких периферийных ветках, более крупные – на толстых ближе к стволу, но фактические данные обычно не приводятся.
Целью нашей работы был анализ размерных характеристик видов синиц рода Parus L. в лесостепном Зауралье и проверка их соответствия правилу Хатчинсона.
Длину крыла и массу тела 5 видов синиц (пухляк P. montanus, московка P. ater, лазоревка P. caeruleus, князек P. cyanus, большая синица P. major) измеряли в ходе неизбирательных отловов мелких кустарниковых видов птиц в период осенних кочевок. Птиц отлавливали паутинными сетями с ячеей 14 мм и длиной 7 и 10 м в августе–октябре 2013 и 2014 гг. и июле–сентябре 2016 г. соответственно в Кетовском, Шадринском и Частоозерском районах Курганской области [15–17]. Всего поймано 1475 особей, длина крыла измерена у 1212 особей, масса тела – у 630. Полученные данные обрабатывали статистически с использованием дисперсионного (ANOVA и MANOVA) и post-hoc анализов. Все расчеты выполнены в программах Statistica 6.0 (StatSoft Ink., 1984–2001) и Microsoft Excel 2003.
На рис. 1 приведены размерные характеристики отловленных особей синиц. Видно, что 4 вида (московка, пухляк, князек, большая синица) выстраиваются по этим характеристикам практически в одну линию и один вид (лазоревка) размещается не между соседними видами (пухляк и князек), а немного сбоку от обоих. Средние значения длины крыла и массы тела изученных видов приведены в табл. 1, отношения размеров пар видов – в табл. 2.
Рис. 1.
Размерно-весовая структура сообщества синиц по данным 2013, 2014 и 2016 гг.: 1 – московка, 2 – пухляк, 3 – лазоревка, 4 – князек, 5 – большая синица. Сплошной линией выделена область лазоревки.
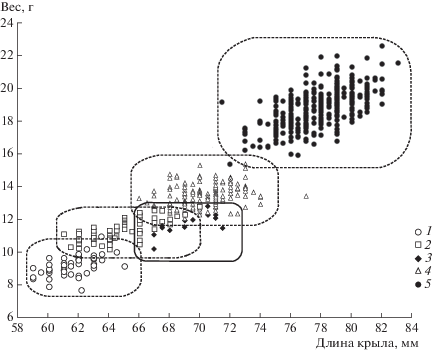
Таблица 1.
Размерно-весовые характеристики синиц, M ± m (n)
| Вид | Длина крыла, мм | Масса тела, г | |||
|---|---|---|---|---|---|
| 2013 г. | 2014 г. | 2016 г. | 2013 г. | 2016 г. | |
| Московка | 61.75 ± 0.32 (16) | 60.36 ± 0.36 (11) | 61.42 ± 0.20 (45) | 9.73 ± 0.20 (9) | 9.08 ± 0.07 (45) |
| Пухляк | 64.54 ± 0.54 (13) | 64.83 ± 0.23 (54) | 64.63 ± 0.23 (67) | 11.20 ± 0.18 (12) | 11.33 ± 0.10 (51) |
| Лазоревка | 69.00 ± 1.05 (5) | 68.58 ± 0.24 (72) | 68.53 ± 0.35 (19) | 11.98 ± 0.29 (5) | 11.56 ± 0.19 (15) |
| Князек | 70.48 ± 0.59 (21) | 70.30 ± 0.21 (56) | 70.06 ± 0.11 (242) | 13.41 ± 0.13 (13) | 13.40 ± 0.06 (143) |
| Большая синица | 77.71 ± 0.18 (144) | 78.05 ± 0.17 (171) | 77.75 ± 0.13 (276) | 18.81 ± 0.12 (89) | 18.91 ± 0.07 (248) |
Таблица 2.
Отношение длины крыла и массы тела у ближайших по размеру видов синиц
| Пары видов | Длина крыла | Масса тела | |||||
|---|---|---|---|---|---|---|---|
| 2013 г. | 2014 г. | 2016 г. | по [18] | 2013 г. | 2016 г. | по [18] | |
| Пухляк/московка | 1.05 | 1.07 | 1.05 | 1.13 | 1.15 | 1.25 | 1.28 |
| Лазоревка/пухляк | 1.07 | 1.06 | 1.06 | 1.06 | 1.07 | 1.02 | 1.0 |
| Князек/лазоревка | 1.02 | 1.03 | 1.02 | 1.01 | 1.12 | 1.16 | 1.13 |
| Князек/пухляк | 1.09 | 1.08 | 1.08 | 1.06 | 1.20 | 1.18 | 1.13 |
| Большая синица/князек | 1.10 | 1.11 | 1.11 | 1.13 | 1.40 | 1.41 | 1.28 |
Двухфакторный дисперсионный анализ (MANOVA) не выявил достоверного влияния года исследований, поэтому мы объединили выборки разных лет. Однофакторный дисперсионный анализ (ANOVA) показал высокую достоверность различий синиц разных видов по длине крыла (df = 4, F = 2412.4, p < 0.001) и массе тела (df = 4, F = 2182.74, p < 0.001). Попарное (post-hoc) сравнение методом Тьюки показало, что только в одном случае нет достоверных различий между видами: по массе тела не различались пухляк и лазоревка (p = 0.606), а остальные различия высоко достоверны (p < 0.001).
Если не учитывать лазоревку, которая проникла в Зауралье лишь в XXI в. (см. ниже), изученная гильдия синиц достаточно хорошо соответствует правилу Хатчинсона. Соотношение средней длины крыла соседних пар видов составляет 1.05–1.11, что немного меньше, чем предсказывает правило Хатчинсона. На наш взгляд, это можно объяснить тем, что мы используем не абсолютные размеры птиц, что довольно сложно оценить, и не размеры пищедобывательных структур, а длину крыла – признак, варьирующий и связанный с общими размерами тела аллометрически и, возможно, нелинейно. При этом в более широком регионе (Урал, Приуралье и Западная Сибирь – по [18]) соотношение этого признака у данных видов оказывается чуть выше: 1.06–1.13. Следует отметить, что и в примерах, приводимых П. Джиллером [2], соотношение длины клюва у тропических птиц было меньше, чем 1.3 (от 1 до 1.7, в среднем 1.15).
Соотношение массы тела у соседних видов синиц, по нашим данным, составляет 1.15–1.41 (по [18] – 1.13–1.28), что также меньше 2, как можно было ожидать по правилу Хатчинсона. В приводимом П. Джиллером [2] примере с новогвинейскими голубями такое соотношение составляло от 1.33 до 2.73, что достаточно близко к нашим значениям.
Однако следует подчеркнуть, что при рассмотрении обоих признаков одновременно перекрывание размерных характеристик у изученных видов синиц незначительно, а между некоторыми видами и вообще отсутствует, что свидетельствует о четком расхождении морфологических признаков.
Существует точка зрения, что половой диморфизм у ряда видов также связан с ослаблением внутривидовой конкуренции за пищу (Schoener, 1977 – цит. по [10]). У всех изученных нами видов синиц размерные распределения оказались двухвершинные, однако надежно определить пол можно лишь у большой синицы. Размерные характеристики самцов и самок этого вида приведены в табл. 3. Видно, что межполовые различия у большой синицы заметно меньше, чем различия между видами синиц, и существенно меньше, чем предсказанные правилом Хатчинсона. Таким образом, половой диморфизм у больших синиц обусловлен, скорее, не конкуренцией за пищу, а другими факторами, вероятно, половым отбором.
Таблица 3.
Размерно-весовые характеристики M ± m (n) и их отношение у самцов и самок больших синиц
| Пол | Длина крыла, мм | Масса тела, г | |||
|---|---|---|---|---|---|
| 2013 г. | 2014 г. | 2016 г. | 2013 г. | 2016 г. | |
| Самцы | 79.49 ± 0.20 (66) | 79.77 ± 0.19 (78) | 79.28 ± 0.13 (130) | 19.42 ± 0.13 (45) | 19.39 ± 0.09 (119) |
| Самки | 76.21 ± 0.15 (77) | 76.62 ± 0.16 (79) | 76.26 ± 0.14 (119) | 18.15 ± 0.14 (43) | 18.27 ± 0.11 (105) |
| Отношение | 1.04 | 1.04 | 1.04 | 1.07 | 1.06 |
Правило Хатчинсона и тесно связанные с ним представления о лимитирующем сходстве лежат в русле проблем размерности экологической ниши и дифференциального перекрывания, что в свою очередь составляет одно из центральных направлений исследования организации многовидовых сообществ и принципов функционирования экосистем [1, 2]. Обычно правило Хатчинсона связывают с пищевым ресурсом, чаще всего с размерами пищевых объектов.
Одной из наиболее ярких иллюстраций правила Хатчинсона является размерная дифференциация у новогвинейских голубей. Различия между видами разных размеров отмечаются, во-первых, по предпочитаемому размеру потребляемых плодов и, во-вторых, по размещению на разных частях ветвей при кормежке [2]. Для синиц также известно расхождение по разным частям кроны при поиске пищи, по тонким и толстым веткам, дальше и ближе к стволу [3–6, 9, 12]. В ряде работ было показано и различие разных видов синиц по пищевым объектам, в том числе их размерам [9, 19–23]. Это важный момент, поскольку различия в размерах предпочитаемых пищевых объектов тесно связаны с расхождением по размерам и самих видов и, прежде всего, их пищедобывательных структур [2]. Впрочем, все авторы отмечают, что спектры питания разных видов синиц могут сильно перекрываться. Возможно, некоторое несоответствие соотношения их массы и размеров правилу Хатчинсона связано как раз не столько с размерами пищевых объектов, сколько с различиями в предпочитаемых участках поиска корма.
Соответствие многовидового сообщества правилу Хатчинсона фактически является следствием конкуренции за один ресурс, а значит, и свидетельством такой конкуренции в прошлом. Нам представляется, что появление морфологических различий – это самый медленный, но и самый экономный путь расхождения по нишам. Другие способы разделения экологических ниш – пространственно-временная и поведенческая дифференциация – могут оказаться энергетически более затратны. Показано, что правило Хатчинсона выполняется в зрелых, сложившихся сообществах, где виды прошли определенный путь совместной эволюции, и фаза острой конкуренции осталась позади [24]. Такую же конкуренцию в прошлом можно предположить и у синиц, учитывая факты их расхождения по ярусам фуражировки, способам поиска и поимки насекомых [3–5] и соответствие правилу Хатчинсона размерных различий (наши данные).
Интересно отметить также, что у синиц увеличивается расхождение признаков по мере роста их размеров (см. табл. 2). Так, различия между московкой и пухляком составляют 1.05–1.07 по длине крыла и 1.15–1.25 по весу, а между князьком и большой синицей – 1.10–1.11 по длине крыла и 1.40–1.41 по весу. Эту закономерность отмечали многие авторы (см. обзор [10]), однако она, на наш взгляд, не является опровержением правила Хатчинсона, а, скорее, дополнением к нему. Более крупные виды питаются более крупной добычей, относительное обилие которой меньше, чем мелких жертв, поэтому конкуренция между ними сильнее и расхождение выше [10]. Следует особо подчеркнуть, что и у Дж. Хатчинсона и его последователей получался довольно большой разброс коэффициентов соотношения, а значение 1.3 – среднее [2].
Единственный вид, который не вписывается в обнаруженную нами закономерность (возможно, пока), – это лазоревка, обитавшая до конца XX в. лишь в европейской части России [25]. Впервые в Зауралье она найдена на гнездовании в июне 1993 г., когда 3 пары заняли искусственные гнездовья в колках в Каменском районе Свердловской области [26]. В Курганской области в июне 2000 г. впервые наблюдали кормивших птенцов птиц в Далматовском районе [27], в мае 2008 г. – выстилавших дупло в долине Тобола [28], а в августе 2016 г. две молодые особи в птенцовом наряде отловлены у восточной границы области [17]. Таким образом, проникнув три десятилетия назад за Урал, лазоревка заселила вскоре всю территорию Курганской области. Сейчас она продолжает интенсивно расселяться дальше на восток по лесостепной зоне и, вероятно, уже достигла Иртыша [29, 30]. В связи с экспансией лазоревки в Зауралье участились случаи ее гибридизации с князьком [29], с которым она весьма близка по длине крыла, но значительно уступает в массе тела (см. табл. 1, 2). Последнее обстоятельство, возможно, способствует снижению конкуренции между этими видами.
По размерно-весовым характеристикам лазоревка размещается не между соседними видами (пухляк и князек), а немного сбоку от обоих (см. рис. 1). Казалось бы, ее морфологическая ниша оказывается уже вполне обособленной. Однако это не совсем так: на рисунке видно, что размерная ниша лазоревки вплотную прижата к нишам пухляка и князька, причем одни лазоревки оказываются среди самых длиннокрылых пухляков, другие – среди самых легких князьков. Этот случай интересен как пример протекающей на наших глазах перестройки многовидового сообщества с внедрением в него нового вида. Прежняя структура сообщества пока не претерпевает существенных изменений, однако новый вид не нашел себе еще новую нишу, и как будет развиваться ситуация, покажет время. В этой связи представляет интерес изучение размерной дифференциации сообществ синиц европейской территории, где князек практически отсутствует и вторым по размерам после большой синицы видом является лазоревка. Может оказаться, что там она крупнее, чем в Зауралье, и занимает размерную нишу князька.
Подобный сдвиг размеров у синиц в условиях наличия–отсутствия более крупных конкурентов отмечали и другие авторы. Так, московка на скандинавских островах при отсутствии двух близких по размерам видов – пухляка и хохлатой синицы P. cristatus – эволюционировала морфологически в их направлении и стала значительно (до 10%) крупнее по линейным размерам, чем в материковой Швеции, где эти три вида сосуществуют [31]. В Южной Корее дальневосточный подвид большой синицы P. m. minor в условиях конкуренции с более крупной тиссовой синицей P. varius стал существенно мельче европейского подвида P. m. major и занял нишу отсутствующей там лазоревки [32].
Таким образом, изученное нами сообщество синиц оказывается четко дифференцированным по размерно-весовым характеристикам, и эта дифференциация в принципе соответствует правилу Хатчинсона, несмотря на несколько меньшие значения индексов. Определяющим в сообществе синиц является расхождение по одной оси экологической ниши, в данном случае пищевой, а меньшие, чем у Хатчинсона, значения индексов могут указывать на то, что заметную роль играет расхождение и по другим осям. В целом наличие выраженной размерной дифференциации открывает новый аспект исследования организации и функционирования многовидовых сообществ мелких воробьиных птиц умеренной зоны.
Работа выполнена в рамках государственного задания Института экологии растений и животных УрО РАН.
Список литературы
Бигон М., Харпер Д., Таунсенд К. Экология: особи, популяции и сообщества. В 2-х т. М.: Мир, 1989. Т. 1. 667 с.; Т. 2. 477 с.
Джиллер П. Структура сообществ и экологическая ниша. М.: Мир, 1988. 184 с.
Боголюбов А.С. Структура и компоновка пространственных ниш видов, входящих в синичьи стаи в лесах Подмосковья // Зоол. журн. 1986. Т. 65. Вып. 11. С. 1664–1674.
Морозов Н.С. Структура пространственных ниш большой синицы (Parus major), лазоревки (P. caeruleus) и обыкновенной пищухи (Certhia familiaris) в дубраве московского парка зимой // Зоол. журн. 1987. Т. 66. Вып. 10. С. 1529–1539.
Шемякина О.А., Марочкина Е.А., Зацаринный И.В., Чельцов Н.В. Механизмы экологической сегрегации четырех совместно обитающих видов синиц – Parus major, P. caeruleus, P. montanus и P. cristatus // Рус. орнитол. журн. 2007. № 362. С. 759–783.
Alatalo R.V. Evidence for interspecific competition among European tits Parus spp.: a review // Ann. Zool. Fenn.1982. V. 19. № 4. P. 309–317.
Atiénzar F., Belda E.J., Barba E. Coexistence of Mediterranean tits: A multidimensional approach // Ecoscience. 2013. V. 20. P. 40–47.
Dhondt A.A. Interspecific competition in birds. Oxford: Oxford University Press, 2012. 282 p.
Lack D. Ecological Isolation in Birds. Oxford: Blackwell Scientific Publications, 1971. 404 p.
Роговин К.А. Морфологическая дивергенция и структура сообществ наземных позвоночных // ВИНИТИ: итоги науки и техники. Зоология позвоночных. 1986. Т. 14. С. 71–126.
Зацаринный И.В., Константинов В.М., Косякова А.Ю. и др. Сравнительная экология двух видов гаичек Восточной Европы (пространственные и трофические аспекты) // Поведение, экология и эволюция животных. Рязань: Голос губернии, 2012. С. 68–131.
Иноземцев А.А. Птицы и лес. М.: Агропромиздат, 1987. 302 с.
Lens L., Dhondt A. Interspecific competition in group territorial tits (Parus spp.): a study of niche shifts // Ann. Soc. Roy. Zool. Belg. 1989. V. 119. № 1. P. 87.
Morse D.H. Structure and foraging patterns of flocks of tits and associated species in an English woodland during the winter // Ibis. 1978. V. 120. № 3. P. 298–312.
Тарасов В.В., Ляхов А.Г. Наблюдения птиц в долине Тобола осенью 2013 года // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. 2013. Вып. 18. С. 197–209.
Тарасов В.В., Ляхов А.Г. Результаты отловов птиц на Исеть-Пышминском водоразделе осенью 2014 года // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. 2014. Вып. 19. С. 159–167.
Тарасов В.В., Ляхов А.Г. Осенние наблюдения птиц на Тоболо-Ишимском междуречье в 2016 году // Фауна Урала и Сибири. 2016. № 2. С. 205–214.
Рябицев В.К. Птицы Урала, Приуралья и Западной Сибири: справ.-определитель. 3-е изд. Екатеринбург: Изд-во Урал. ун-та, 2008. 634 с.
Иноземцев А.А. Динамика экологических ниш синиц и правило конкурентного исключения // Экология. 1987. № 5. С. 49–56.
García-Navas V., Ferrer E.S., Sanz J.J. Prey choice, provisioning behaviour, and effects of early nutrition on nestling phenotype of titmice // Ecoscience. 2013. V. 20. P. 9–18.
Navalpotro H., Pagani-Núñez E., Hernández-Gómez S., Senar J.C. Comparing prey composition and prey size delivered to nestlings by great tit, Parus major, and blue tit, Cyanistes caeruleus, in a Mediterranean sclerophyllous mixed forest // Animal Biodiversity and Conservation, 2016. V. 39.1. P. 129–139.
Park C.-R., Lee W.-S., Hino T. Temporal changes in foraging niche among breeding tits (Paridae) in a Korean temperate deciduous forest // Ornis Fennica. 2005. V. 82. P. 81–88.
Török J., Tóth L. Asymmetric competition between two tit species: a reciprocal removal experiment // J. of Animal Ecology. 1999. V. 68. P. 338–345.
Гилев А.В., Зиновьев Е.В., Нестерков А.В. Размерная дифференциация видов и организация многовидовых сообществ у муравьев // Успехи соврем. биол. 2007. Т. 127. № 2. С. 209–213.
Степанян Л.С. Конспект орнитологической фауны СССР. М.: Наука, 1990. 728 с.
Бельский Е.А., Ляхов А.Г. Орнитологические находки в Каменском районе Свердловской области // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. 1995. Вып. 1. С. 6–7.
Тарасов В.В. Заметки к фауне птиц Курганской области // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. 2000. Вып. 5. С. 180–181.
Тарасов В.В., Давыдов А.Ю. Новости по орнитофауне Курганской области // Материалы к распространению птиц на Урале, в Приуралье и Западной Сибири. 2008. Вып. 13. С. 100–108.
Ляхов А.Г., Тарасов В.В. Встречи гибридов белой Parus cyanus и обыкновенной P. caeruleus лазоревок на Среднем Урале и в Зауралье // Рус. орнитол. журн. 2014. № 1085. С. 4041–4045.
Нефедов А.А. К орнитофауне Омской области // Рус. орнитол. журн. 2017. № 1447. С. 2043–2079.
Norberg R.Å., Norberg U.M.L. Evolution of enlarged body size of coal tits Parus ater in geographic isolation from two larger competitors, the crested tit Parus cristatus and the willow tit Parus montanus, on six Scandinavian islands // Biology Open. 2015. V. 4. P. 1490–1508.
Jablonski P.G., Lee S.D. Foraging niche shifts in mixed-species flocks of tits in Korea // J. of Field Ornithology. 2002. V. 73. № 3. P. 246–252.
Дополнительные материалы отсутствуют.