

Экология, 2020, № 3, стр. 163-170
Краевой эффект, особенности роста и морфогенеза деревьев сосны обыкновенной в лесопарках и естественных насаждениях
С. А. Шавнин a, *, Д. Ю. Голиков a, А. А. Монтиле a, **, А. И. Монтиле a°, b
a Ботанический сад УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202а, Россия
b Уральский государственный лесотехнический университет
620100 Екатеринбург, Сибирский тракт, 37, Россия
* E-mail: sash@botgard.uran.ru
** E-mail: org17@mail.ru
Поступила в редакцию 13.06.2019
После доработки 25.10.2019
Принята к публикации 12.11.2019
Аннотация
С помощью метода обследования трансект изучали степень проявления и глубину краевого эффекта в длительно существующих в условиях урбанизации (г. Екатеринбург, Россия) фрагментах лесопарковых насаждений и аналогичных естественных древостоях. Использовали метод количественной оценки выраженности ростовых особенностей ствола и кроны деревьев по комплексу морфологических признаков. Объектами исследований были границы насаждений сосны обыкновенной. Из пяти оцениваемых признаков наличие краевого эффекта установлено по охвоению и сучковатости, характеризующим развитость ассимиляционного аппарата деревьев. Общие тренды указывают на увеличение этих характеристик с удалением от опушки в естественных древостоях и уменьшение – в лесопарке. Нелинейность проявления признаков вдоль трансект связана с различными типами ответных реакций деревьев на комплекс определяющих морфогенез факторов. В естественных насаждениях выделены четыре статистически значимо отличающиеся зоны, расположенные на расстояниях 0–75, 100–125, 150–175 и 200–225 м от края насаждения, в лесопарке только две зоны. Унифицированный подход к анализу данных позволяет заключить, что в обоих местообитаниях граница краевого эффекта наблюдается на удалении 75–100 м по сучковатости, а по охвоению – на 100–125 м. Выявлено наличие нескольких типов морфогенетических реакций деревьев, различаемых по степени изменений охвоения и сучковатости в условиях краевого эффекта.
Урбанизация лесопокрытых территорий усиливает фрагментацию лесов. В связи с этим возрастает актуальность изучения влияния краевых эффектов на состояние лесных экосистем и их биологическое разнообразие в различных условиях городской и пригородной локализации [1–5]. К основным сопутствующим фрагментации факторам относятся изменение совокупности микроклиматических условий среды [6] и влияние комплекса биотических факторов (внутри- и межвидовая конкуренция, изменение состава растительных сообществ и других компонентов биоты, а также свойств почв) [7]. Известно, что первичные изменения на границе насаждений вызывают долговременные ответные реакции биоценозов и их развитие в глубь леса. По разным оценкам для древостоев глубина краевого эффекта составляет 20–100 м [5, 8–11], у растительности травяно-кустарничкового яруса – около 30 м [12], а в тропических лесах в зависимости от конкретных характеристик – до 300 м [13].
Ранее при изучении загородных, лесопарковых и внутригородских сосновых насаждений было установлено повышение индекса жизненного состояния деревьев (густоты охвоения и времени жизни хвои) в глубине древостоя [11]. Эти изменения сопровождаются трансформацией морфологической структуры древостоя, в частности на крае насаждения уменьшаются густота [14] и относительная высота деревьев как показатель их световой потребности [1, 8, 15, 16], при этом в старшевозрастных насаждениях краевой эффект менее выражен [17].
Исследование краевого эффекта на молодых и более старых границах леса с помощью метода закладки и обследования трансект показало, что с удалением от края насаждения высота, диаметр, жизненное состояние, густота и запас сосновых древостоев линейно возрастают [18]. При этом нелинейных зависимостей вплоть до 250 м не установлено – на старых трансектах краевой эффект был статистически значим, а на молодых не наблюдался. Ввиду того, что развитие краевого эффекта в глубину насаждения происходит на протяжении десятков лет [6], сопровождающие это явление процессы связаны с адаптацией к действию совокупности внешних факторов. Приспособительный характер реакций на уровне отдельных деревьев и древостоя позволяет предположить, что одним из проявлений краевого эффекта может быть реализация морфогенетического адаптационного потенциала в виде накопления в течение онтогенеза отдельных морфологических признаков.
Ранее на примере сосновых насаждений было показано [19], что количественный подход к оценке отдельных характеристик формы ствола и кроны, отражающих интегральные реакции деревьев на действие внешних факторов, эффективен и перспективен для изучения их роста и развития. В связи с этим при планировании данного исследования были выдвинуты две рабочие гипотезы. Первая – в градиентах, вызывающих краевой эффект условий среды, наблюдается комплекс адаптационных реакций деревьев, проявляющихся в виде особенностей их роста и морфогенеза. При этом некоторые морфометрические признаки являются информативными, так как закономерно изменяются в зависимости от расстояния до края насаждения. Вторая – пространственная динамика значений признаков различна в естественных насаждениях и лесопарке.
Основная цель настоящего исследования – выявление в зоне краевого эффекта информативных для изучения пространственной динамики морфометрических признаков сосны обыкновенной (Pinus sylvestris L.) и определение (на основе сравнительного анализа их величин) степени проявления и глубины краевого эффекта в длительно существующих фрагментах естественных насаждений и лесопарка, расположенного на территории крупного промышленного центра (г. Екатеринбург).
Для проверки высказанных гипотез были поставлены следующие задачи:
1. Провести измерения комплекса характеристик роста и морфогенеза деревьев на репрезентативном количестве трансект, длина которых позволяет фиксировать краевой эффект в естественных насаждениях и лесопарке.
2. Проанализировать данные и выявить чувствительные к краевому эффекту (информативные) признаки, определив наличие и характер связи величин признаков с расстоянием до края насаждения и его влияние на варьирование отдельных характеристик.
3. На основании анализа зависимостей информативных признаков от расстояния до края насаждения, с дополнительным привлечением кластерного анализа установить и сравнить границы зон, отличающихся характером проявления краевого эффекта в изучаемых древостоях.
МАТЕРИАЛ И МЕТОДЫ
Исследования проводили в 2015–2017 гг. в лесопарке и естественных сосновых насаждениях. Лесорастительные условия и характеристика антропогенных факторов, влияющих на состояние насаждений г. Екатеринбурга, соответствуют описанным ранее [11]. Изучаемые участки относятся к Зауральской предгорной провинции Южнотаежного округа [20]. Территория города подвержена влиянию аэропромышленных загрязнений, среди которых преобладают выбросы автотранспорта, при этом в почвах накоплено значительное количество тяжелых металлов [21–23].
Для оценки наличия и глубины краевого эффекта использовали метод закладки и обследования трансект. Условия подбора участков описаны ранее [18]. На каждой трансекте закладывали по 10 учетных площадок радиусом 11.3 м, которые располагали в глубь от края насаждения с расстояниями 25 м между их центрами. Трансекты создавали на возникших более 20 лет назад краях насаждений, расположенных в лесопарке Юго-Западный и на удалении 10–30 км от города в южном и западном направлениях (вблизи населенных пунктов Арамиль, Бобровский, Двуреченск, Кашино, Хрустальный). Всего обследовано 6 трансект в лесопарке и 10 – в естественных насаждениях (номера 7–9, 11, 14, 15 и 16–19, 23–25, 28–30 соответственно). Из них трансекты 14 и 18 были укороченными (6 учетных площадок). На каждой площадке проводили сплошной перечет деревьев c диаметром не менее 8 см. Всего обследовали 2315 деревьев, растущих на 152 площадках.
Для изучения краевого эффекта при определении диагностических признаков деревьев использовали метод количественной оценки выраженности ростовых особенностей, связанных с морфогенетической адаптацией [19]. Измеряемые характеристики ствола и кроны (многоствольность, многовершинность, кривизна ствола, сучковатость, охвоение ветвей) возникают и изменяются в ходе онтогенеза отдельного дерева. Кривизна ствола определяется по количеству изгибов со стрелой прогиба не менее 0.5 диаметра, сучковатость – по количеству и разветвленности живых и отмерших сохранившихся ветвей на всем стволе, охвоение характеризует количество, степень густоты и распределение хвои в кроне. Таким образом, два последних признака описывают развитость ассимиляционного аппарата дерева. При этом все признаки являются измеряемыми дискретно во времени, а их величины определяются с помощью балльных шкал.
Статистический анализ полученных данных проводили с помощью стандартного пакета прикладных программ Statistica 8.0 (StatSoft Inc., 2007). Использовали методы регрессионного, кластерного и непараметрический аналог однофакторного дисперсионного анализов. При проведении кластерного анализа применяли евклидову метрику и метод Уорда. Для визуального представления пространственной динамики показателей с удалением от края насаждения рассчитывали стандартную ошибку и 95%-ный доверительный интервал для среднего на соответствующих площадках. Так как значения признаков сучковатости и охвоения изначально измерены в баллах, а их распределения обладают значительной асимметрией (стандартизованная асимметрия от –2.6 до –10.4), при их анализе использовали медианы в качестве центральной тенденции и ранговый критерий Краскела-Уоллиса при сравнении центральных тенденций.
РЕЗУЛЬТАТЫ
Анализ результатов исследований показал, что на всех трансектах в насаждениях преобладают одноствольные деревья, а краевой эффект по признаку многоствольности не проявляется. Многовершинность и степень искривления стволов в лесопарке в среднем выше, чем за городом (H(1; 2315) = 24.88; p < 0.001 и H(1; 2315) = 12.89; p < 0.001 соответственно), однако корреляций значений этих признаков с расстоянием и влияния фактора расстояния на их вариацию в зоне краевого эффекта не обнаружено. Таким образом, проявлений краевого эффекта для данных признаков не установлено. Следует отметить, что величины сучковатости и охвоения выше в естественных насаждениях по сравнению с лесопарком (H(1; 2315) = 42.85; p < < 0.001 и H(1; 2315) = 68.25; p < 0.001 соответственно) и, как показано далее, изменяются с удалением от края насаждения.
В связи с этим на первом этапе изучения краевого эффекта в естественных насаждениях и лесопарке был проведен кластерный анализ пространственной динамики величин сучковатости и охвоения вдоль трансект. Признаками для анализа служили средние величины на каждой площадке трансекты, а объектами – изменения признака в зависимости от расстояния до края насаждения. Вследствие того, что величины данных характеристик находятся приблизительно в одном диапазоне (около 3 баллов), при расчете расстояний между пространственными изменениями признаков учитывали оба показателя отдельно. Кластеризация позволяет выделить две основные группы, состав которых, за отдельными исключениями, соответствует разделению трансект на лесопарковые и загородные (рис. 1). Анализ всех трансект, включая укороченные 14 и 18, по 6 площадкам подтверждает данную закономерность. Следует отметить, что при кластеризации всех деревьев в обоих местообитаниях по указанным выше признакам совместно выделяются 4 кластера (в связи с большим массивом данных и сложностью их представления результаты не приведены). Из них два кластера объединяют деревья с наибольшими и наименьшими значениями параметров соответственно и два – с промежуточными.
Рис. 1.
Кластеризация пространственной динамики величин сучковатости и охвоения на трансектах: Суч. и Охв. – сучковатость и охвоение; 7–30 – номера трансект.
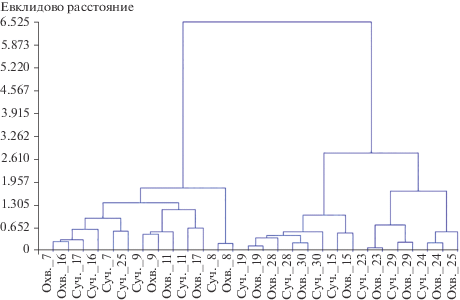
Установленные отличия трансект в лесопарке и естественных насаждениях указывают на целесообразность изучения краевого эффекта раздельно для данных местообитаний. В связи с этим в дальнейшем мы сравнивали усредненные значения величин признаков на этих объектах независимо.
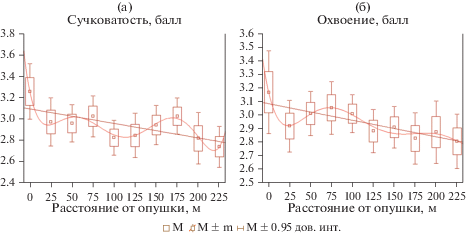
Сравнение естественных древостоев и лесопарков показывает, что линейные тренды величин сучковатости и охвоения вдоль трансект у них различаются (рис. 2, 3): у первых по данным признакам тренды слабые и положительные, а в лесопарке – отрицательные. При анализе зависимостей признаков от расстояния до края насаждения величины характеристик усредняли. В целях визуального представления формы зависимости признака от расстояния на рис. 2 и 3 дополнительно приведены полиномиальные тренды 6-го порядка, все коэффициенты которых, в отличие от полиномов других порядков, значимы (p < 0.05). Согласно полученным данным, на расстоянии 25 м от края насаждения наблюдается значимое снижение охвоения и сучковатости. С увеличением расстояния от края насаждения в лесопарке (см. рис. 3) отмечается относительная стабилизация этих показателей. В естественных насаждениях последовательно чередуются возрастание и снижение величин обоих признаков (см. рис. 2), при этом величины и положение экстремумов на регрессионных кривых значительно отличаются от соответствующих характеристик в лесопарке. Сложный вид полученных зависимостей затрудняет определение глубины краевого эффекта. Для ее установления было использовано разбиение данных всех трансект на группы, каждая из которых объединяет результаты измерений на смежных площадках. Принадлежность дерева к группе, соответствующей диапазону расстояний от края насаждения, рассматривалась как фактор, влияющий на сучковатость и охвоение.
Рис. 2.
Линейные, полиномиальные тренды и усредненные величины сучковатости (а) и охвоения (б) на разном расстоянии от опушки в естественных насаждениях.
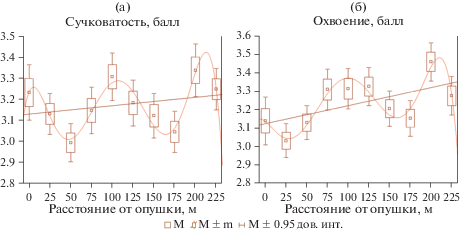
Рис. 3.
Линейные, полиномиальные тренды и усредненные величины сучковатости (а) и охвоения (б) на разном расстоянии от опушки в лесопарке.
Полный анализ всей комбинаторики пространственных зон для количества групп от 6 до 2 и количества объединяемых в группы площадок от 2 до 5 позволил установить на трансектах в естественных насаждениях четыре статистически значимо отличающиеся по обоим признакам зоны (см. табл. 1) с центрами площадок, расположенными на расстояниях 0–75, 100–125, 150–175 и 200–225 м от края насаждения.
Таблица 1.
Оценка значимости влияния расстояния от края насаждения на величины охвоения и сучковатости при выделении четырех зон в естественных насаждениях
| № зоны | Расстояние до центра площадки, м | Охвоение | Сучковатость | ||
|---|---|---|---|---|---|
| средний ранг | H-статистика; p-уровень | средний ранг | H-статистика; p-уровень | ||
| 1 | 0–75 | 825.87 | 32.80; <0.001 | 845.51 | 22.63; <0.001 |
| 2 | 100–125 | 959.85 | 942.94 | ||
| 3 | 150–175 | 838.05 | 823.45 | ||
| 4 | 200–225 | 978.25 | 964.98 | ||
В результате аналогичного анализа в лесопарке выделяются только две статистически значимо различающиеся группы площадок. При этом по сучковатости граница между ними локализуется на расстоянии 75–100 м от края насаждения (H(1;554) = 6.56; p = 0.010), а по охвоению – 100–125 м (H(1;554) = 8.33; p = 0.004). Величины этих расстояний в дальнейшем рассматриваются в качестве оценок глубины краевого эффекта по каждому из признаков.
Для сопоставления результатов по трансектам в лесопарке и естественных насаждениях при анализе последних рассмотрены варианты объединения площадок в 2 группы, из которых выбран один с лучшими относительно других показателями статистической значимости. Это позволило установить следующие границы краевого эффекта: по сучковатости – на расстоянии 75–100 м (H(1;1761) = 6.84; p = 0.009), а по охвоению – 100–125 м (H(1;1761) = 8.29; p = 0.004). Данные значения совпадают с локализацией границ краевого эффекта в лесопарке, однако следует отметить, что лучшие показатели для трансект в естественных насаждениях при выделении двух зон значительно уступают (в несколько раз) соответствующим показателям для четырех зон.
Анализ соотношения частот встречаемости деревьев с одновременно максимальными либо минимальными значениями двух чувствительных к краевому эффекту признаков (рис. 4) показал, что в естественных насаждениях формы кривых их зависимостей от расстояния до края насаждения характеризуются чередованием противоположно направленных экстремумов. При этом доминируют особи с минимальными значениями признаков. В лесопарке наблюдается аналогичная закономерность изменений частот, однако их пространственная динамика существенно отличается – экстремумы менее выражены и преобладают особи с максимальными проявлениями признаков.
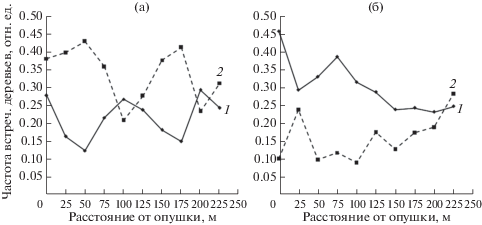
ОБСУЖДЕНИЕ
Основным фактором, вызывающим многоствольность и многовершинность сосны, является механическое или вызванное энтомовредителями повреждение верхушечного побега в молодом и более позднем возрастах, приводящее к последующему формированию нескольких стволов или вершин. Отсутствие отличий между древостоями в разных местообитаниях по первому признаку и достоверное увеличение многовершинности в лесопарке свидетельствуют о росте в условиях городской среды влияния данного фактора со временем. Причины формирования искривлений ствола сосны, помимо очевидных реакций на внешние воздействия (механические повреждения, гравии- и фототропизмы), до настоящего времени не установлены. Объяснение механизмов этого явления и роста его проявлений в городских условиях требует дополнительного изучения.
Отсутствие влияния краевого эффекта на три из пяти измерявшихся признаков связано с независимостью их формирования и указывает на то, что они не чувствительны к действию сопряженных с наличием краев насаждений особенностей условий среды [5]. Сучковатость и охвоение являются интегральными показателями и характеризуют развитость ассимиляционного аппарата в целом. Их более высокие значения и отличие пространственных динамик на трансектах в естественных древостоях по сравнению с лесопарком (см. рис. 1) свидетельствуют о наличии реакции определяющих формирование этих признаков процессов в зоне краевого эффекта. Противоположность их линейных трендов при увеличении расстояния до края насаждения (см. рис. 2, 3) объясняется влиянием урбанизации на состояние ассимиляционного аппарата деревьев. Совокупность приведенных фактов подтверждает первую гипотезу и, частично, вторую.
Вид кривых трендов, аппроксимирующих данные взаимосвязи сучковатости и охвоения с расстоянием, свидетельствует о существенной нелинейности их проявления вдоль трансект, которая связана, по-видимому, с наложением влияний факторов среды на измерявшиеся характеристики ответных реакций деревьев. Так, снижение показателей при удалении от края насаждения до 25 м объясняется очевидным изменением условий освещения [8] и температурного режима [1, 6]. В лесопарке этот эффект выше, чем в естественных насаждениях, что позволяет предположить наличие в городе пока неустановленных дополнительных формирующих краевой эффект факторов. Следует отметить, что в лесопарке, согласно расчетам относительных морфологических показателей деревьев сосны, наблюдается выражающееся в понижении относительных высот деревьев изменение строения древостоев вблизи края насаждения [16].
Результаты однофакторного дисперсионного анализа зависимостей охвоения и сучковатости от расстояния до края насаждения позволяют сделать предположение о структурированном (четыре различающиеся зоны) и возможно распространяющемся на существенное расстояние (около 200 м) влиянии краевого эффекта на пространственную динамику этих показателей сосны на загородных трансектах. Различия в количестве и локализации границ между зонами по данным признакам для естественных насаждений и лесопарка (в последнем выделяются только две зоны) дополнительно свидетельствуют об изменении состояния ростовых процессов деревьев в городе. При сокращении в ходе анализа данных о количестве зон в естественных насаждениях до двух (с целью сравнения результатов с лесопарком) установлено совпадение положений границ между ними. Если принять условие, что расстояние от края насаждения до границы между первой и второй зонами составляет глубину краевого эффекта, то она больше на 25 м для охвоения по сравнению с сучковатостью, что, по-видимому, отражает более высокую чувствительность данного признака к факторам среды. Следует отметить, что результаты анализа сведений о величинах четырех из пяти изучавшихся признаков (за исключением многоствольности), а также о пространственной динамике охвоения и сучковатости в изученных местообитаниях подтверждает правильность второй гипотезы.
Наличие четырех кластеров, выделенных при анализе данных всех (независимо от местоположения) обследованных по совокупности величин сучковатости и охвоения деревьев (группы с максимальными и минимальными значениями, а также две – с промежуточными), позволяет предположить существование в зоне краевого эффекта четырех различных типов ответных реакций деревьев сосны на комплекс определяющих морфогенез факторов. Кроме того, в ходе исследований установлено значительное несовпадение пространственных динамик встречаемости деревьев с контрастно отличающимися типами морфогенетических ответных реакций в естественных насаждениях и лесопарке. Следует отметить, что найденные закономерности и предположение о существовании типов адаптационных морфогенетических реакций основаны на результатах анализа величин признаков развитости ассимиляционного аппарата и согласуются с наличием разных стратегий роста деревьев по классификации Крафта.
ВЫВОДЫ
1. В результате исследований особенностей морфогенеза деревьев сосны установлено, что по многоствольности естественные насаждения и лесопарки не отличаются. Искривления стволов и многовершинность достоверно чаще отмечаются в лесопарке, однако по этим признакам краевой эффект отсутствует.
2. Краевой эффект наблюдается по признакам охвоения и сучковатости, линейные тренды изменений которых свидетельствуют об увеличении этих характеристик с удалением от края насаждения в естественных древостоях и уменьшении – в лесопарке. Пространственные динамики величин данных признаков также различны.
3. Нелинейность проявления признаков сучковатости и охвоения вдоль трансект указывает на сложный характер процессов, определяющих формирование краевого эффекта в древостое. Одной из причин нелинейности установленных зависимостей является наличие как в естественных древостоях, так и в лесопарке четырех типов ответных реакций деревьев на комплекс определяющих морфогенез факторов. От их сочетания, по-видимому, зависит формирование краевого эффекта.
4. Однофакторный дисперсионный анализ зависимости величин сучковатости и охвоения от расстояния до края насаждения позволяет выделить на трансектах в естественных насаждениях четыре статистически значимо отличающиеся по влиянию на вариацию значений данных признаков зоны. Центры входящих в зоны площадок расположены на расстояниях 0–75, 100–125, 150–175 и 200–225 м от опушки. Наличие нескольких зон указывает на сложный характер краевого эффекта в естественных древостоях. В лесопарке статистически значимо выделяются только две зоны, что связано, по-видимому, с изменением количества и/или вклада влияющих факторов.
5. Унифицированный подход к определению границы краевого эффекта, включающий выделение двух зон, позволяет сравнивать условную глубину эффекта в двух местообитаниях. В естественных древостоях и лесопарке краевой эффект по сучковатости наблюдается на удалении 75–100 м, а по охвоению – 100–125 м. Отличие глубины эффекта по данным характеристикам составляет около 25 м и объясняется, по-видимому, особенностями адаптационных морфогенетических реакций деревьев. Общая глубина краевого эффекта в лесопарке, с учетом обоих вариантов зонирования по каждому признаку, составляет около 125 м, а в естественных насаждениях – 175 м. Наиболее однонаправленно по признакам охвоения и сучковатости краевой эффект проявляется на расстоянии до 25 м от края насаждения.
6. Выявлено наличие нескольких типов морфогенетических реакций деревьев, различаемых по степени изменений охвоения и сучковатости в условиях краевого эффекта.
Авторы выражают благодарность д.б.н. Д.В. Веселкину за обсуждение проблемы и непосредственное участие в подборе объектов исследования. Авторы также благодарят рецензента за конструктивные замечания, учет которых позволил улучшить форму изложения и содержание статьи.
Работа выполнена при финансовой поддержке Комплексной программы УрО РАН на 2018–2020 гг. (грант № 18-4-4-10) и в рамках госзадания Ботанического сада УрО РАН.
Список литературы
Chen J., Franklin J.F., Spies T. Vegetation responses to edge environments in old-growth Douglas-fir forests // Ecol. Appl. 1992. V. 2. № 4. P. 387–396. https://doi.org/10.2307/1941873
Harper K.A., Macdonald S.E. Structure and composition of riparian boreal forest: new methods for analyzing edge influence // Ecology. 2001. V. 82. № 3. P. 649–659. https://doi.org/10.1007/S11258-006-9227-2
Harper K.A., Macdonald S.E. Structure and composition of edges next to regenerating clear-cuts in mixed-wood boreal forest // J. of Vegetation Science. 2002. V. 13. № 4. P. 535–546. https://doi.org/10.1111/j.1654-1103.2002.tb02080.x
Harper K.A., Macdonald S.E. Quantifying distance of edge influence: a comparison of methods and a new randomization method // Ecosphere. 2011. V. 2(8). P. 1–17. https://doi.org/10.1890/ES11-00146.1
Harper K.A., Macdonald S.E., Burton P.J. et al. Edge influence on forest structure and composition in fragmented landscapes // Conservation Biology. 2005. V. 19. № 3. P. 768–782. https://doi.org/10.1111/j.1523-1739.2005.00045.x
Chen J., Franklin J.F., Spies T.A. Microclimatic pattern and basic biological responses at the clearcut edges of old-growth Douglas-fir stands // Northwest Environment. 1990. V. 6(2). P. 424–425.
Рулев А.С., Рулева О.В., Юферев В.Г., Рулев Г.А. Термодинамика экотонных ландшафтов // Вестн. Воронежского гос. ун-та. Серия: География. Геоэкология. 2017. № 4. С. 5–14.
Murcia C. Edge effects in fragmented forests: implications for conservation // Trends in Ecology & Evolution. 1995. V. 10. № 2. P. 58–62. https://doi.org/10.1016/S0169-5347(00)88977-6
Kellman M. Redefining roles: Plant community reorganization and species preservation in fragmented systems // Global Ecology and Biogeography Letters. 1996. V. 5. № 3. P. 111–116. https://doi.org/10.2307/2997393
Пупырев Е.И., Якубов Х.Г., Авсиевич Н.А. Мониторинг состояния зеленых насаждений в условиях мегаполиса (состояние, проблемы и перспективы развития мониторинга в 2000 г.) // Вестн. Московского гос. ун-та леса – Лесной вестник. 2000. № 6(15). С. 12–15.
Шавнин С.А., Веселкин Д.В., Воробейчик Е.Л. и др. Факторы трансформации сосновых насаждений в районе города Екатеринбурга // Лесоведение. 2015. № 5. С. 346 – 355. [Shavnin S.A., Veselkin D.V., Vorobeichik E.L. et al. Factors of pine-stand transformation in the city of Yekaterinburg // Contemp. Probl. Ecol. 2016. V. 9. № 7. P. 844–852.] https://doi.org/10.1134/S199542551607009X
Веселкин Д.В., Коржиневская А.А., Подгаевская Е.Н. Краевой эффект для травяно-кустарничкового яруса урбанизированных южнотаежных лесов // Экология. 2018. № 6. С. 411–420. [Veselkin D.V., Korzhinevskaya A.A., Podgaevskaya E.N. The edge effect on the herb-dwarf shrub layer of urbanized southern taiga forests // Rus. J. Ecol. 2018. V. 49. № 6. P. 465–474.] https://doi.org/10.1134/S1067413618060139
Laurance W.F. Hyperdynamism in fragmented habitats // Journal of Vegetation Science. 2002. V. 13, № 4. P. 595–602. https://doi.org/10.1111/j.1654-1103.2002.tb02086.x
Kuliesis A.A., Kuliesis A. Edge effect on forest stand growth and development // Baltic Forestry. 2006. V. 12. № 2. P. 158–169.
Reinmann A.B., Hutyra L.R. Edge effects enhance carbon uptake and its vulnerability to climate change in temperate broadleaf forests // Proceedings of the Nat. Acad. of Sci. of the USA. 2017. V. 114. № 1. P. 107–112. https://doi.org/10.1073/pnas.1612369114
Галако В.А., Шавнин С.А., Власенко В.Э. и др. Особенности морфологической структуры сосновых древостоев пригородных насаждений г. Екатеринбурга // Изв. Оренбургского гос. аграрного ун-та. 2017. № 5(67). С. 88–90.
Spies T.A., Franklin J.F. Old-growth and forest dynamics in the Douglas-fir region of western Oregon and Washington // Natural Areas J. 1988. V. 8. № 3. P. 190–201.
Веселкин Д.В., Шавнин С.А., Воробейчик Е.Л. и др. Краевые эффекты для сосновых древостоев в крупном городе // Экология. 2017. № 6. С. 405–412. [Veselkin D.V., Shavnin S.A., Vorobeichik E.L. et al. Edge effects on pine stands in a large city // Rus. J. Ecol. 2017. V. 48. № 6. P. 499–506.] https://doi.org/10.1134/S1067413617060121
Шавнин С.А., Овчинников И.С., Монтиле А.А., Голиков Д.Ю. Оценка формы ствола и кроны сосны обыкновенной по комплексу морфологических признаков // Лесоведение. 2019. № 1. С. 64–74.
Колесников Б.П., Зубарева Р.С., Смолоногов Е.П. Лесорастительные условия и типы лесов Свердловской области: Практическое руководство. Свердловск: Изд-во УНЦ АН СССР, 1974. 176 с.
Стурман В.И. Природные и техногенные факторы загрязнения атмосферного воздуха российских городов // Вестн. Удмуртского ун-та. Биология. Науки о земле. 2008. Вып. 2. С. 15–29.
Государственный доклад “О состоянии и об охране окружающей среды Свердловской области в 2010 г.” Екатеринбург, 2011. 350 с.
Государственный доклад “О состоянии и об охране окружающей среды Свердловской области в 2012 г.” Екатеринбург, 2013. 307 с.
Дополнительные материалы отсутствуют.