

Экология, 2020, № 4, стр. 316-320
Изменчивость жевательной поверхности М3 и М1 моляров у рыжей полевки в циклической популяции
М. И. Чепраков *
Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: cheprakov@ipae.uran.ru
Поступила в редакцию 03.06.2019
После доработки 26.09.2019
Принята к публикации 04.10.2019
В циклической популяции рыжей полевки (Clethrionomys glareolus Schreber, 1780), изучению которой посвящено данное исследование, выявлено множество эффектов популяционной плотности [1]. Установлено, что у этого вида складчатость М3 и М1 моляров меняется в зависимости от фаз популяционного цикла [2, 3]. В обеих работах использованы материалы по прибылым особям. Однако именно перезимовавшие особи осуществляют преемственность в популяции от предыдущего года к следующему. В начале сезона размножения при любом уровне численности только они обеспечивают воспроизводство популяции и формирование ее репродуктивного ядра, а в годы пиков именно они составляют основу ядра популяции на протяжении всего сезона размножения. Поэтому принципиально важно знать, как изменяются параметры жевательной поверхности М3 и М1 моляров у перезимовавших полевок в циклической популяции.
Цель настоящей работы – изучить влияние фаз популяционного цикла, пола, возраста и положения в челюсти (правая и левая части) на изменчивость строения жевательной поверхности М3 и М1 моляров у перезимовавших рыжих полевок. Для этого необходимо было: 1) оценить влияние факторов (независимых переменных) на изменчивость таких составляющих жевательной поверхности М3 и М1 моляров, как складчатость (число выступающих углов) и число замкнутых дентиновых полей; 2) выявить взаимодействия факторов и раскрыть их содержание; 3) изучить взаимосвязи между составляющими изменчивости жевательной поверхности в пределах одного моляра и между молярами.
Наблюдения за динамикой обилия локальной популяции рыжей полевки проводили на краевом участке елово-пихтового леса площадью около 8 га на Среднем Урале (локалитет Шигаево, 57.35° с.ш., 58.71° в.д.). Этот вид занимает доминирующее положение в изучаемом сообществе мелких млекопитающих. С 1999 г. по 2017 г. раз в сезон во второй половине июля проводили отловы животных живоловушками, помещенными в линию с интервалом 7 м. Приманкой служил хлеб с подсолнечным маслом. Ловушки проверяли 3 раза в сутки: утром, днем и вечером. Для оценки популяционного обилия рыжей полевки использовали показатели попадаемости на 100 лов.-сут в первые два дня отлова. Выделяли три уровня численности: низкую (от 0 до 12, спад), среднюю (от 13 до 25, подъем) и высокую (более 25 ос/100 лов.-сут, пик). Подробности изложены в работе [1]. Возраст полевок определяли по уравнению, связывающего их возраст (y) в днях с индексом корня второго верхнего моляра, полученному как отношение длины корня к высоте зуба (x): y = 70 + 426.4x. Это уравнение было вычислено по данным Г.В. Оленева [4].
Для каждой особи определяли дату рождения. Общее количество исследованных перезимовавших полевок составило 172 экз. При анализе использовали правый и левый моляры. Использование правых и левых моляров в одной выборке позволяет избежать погрешности, связанной с асимметрией морфотипов на индивидуальном уровне [5]. Всего было изучено 338 третьих верхних моляров M3 (табл. 1) и 342 первых нижних моляра M1 (табл. 2). Собранные материалы хранятся в Музее ИЭРиЖ УрО РАН, г. Екатеринбург. Изменчивость моляров оценивали по двум составляющим: складчатости (числа выступающих углов) и числу замкнутых дентиновых полей (рис. 1).
Таблица 1.
Число моляров (экз.) для двух составляющих изменчивости жевательной поверхности М3 и группирующих факторов
| Возраст, мес. |
Пол | Фазы цикла | Категории складчатости | Категории числа замкнутых полей | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | всего | 1 | 2 | 3 | всего | |||
| 10–12 | Самцы | Спад Подъем Пик |
3/4 1/1 13/11 |
2/1 3/3 13/15 |
0/0 0/0 5/5 |
5/5 4/4 31/31 |
0/0 2/1 7/8 |
3/3 1/2 22/18 |
2/2 1/1 2/5 |
5/5 4/4 31/31 |
| Самки | Спад Подъем Пик |
2/2 0/0 4/5 |
1/1 2/2 19/17 |
0/0 0/0 6/7 |
3/3 2/2 29/29 |
1/0 2/2 11/5 |
1/1 0/0 14/20 |
1/2 0/0 4/4 |
3/3 2/2 29/29 |
|
| 13–15 | Самцы | Спад Подъем Пик |
3/4 8/10 18/17 |
6/4 11/9 11/11 |
1/2 0/0 1/2 |
10/10 19/19 30/30 |
1/1 8/8 11/13 |
6/5 8/9 15/15 |
3/4 3/2 4/2 |
10/10 19/19 30/30 |
| Самки | Спад Подъем Пик |
7/7 3/3 5/7 |
4/6 4/4 7/7 |
2/0 0/0 4/2 |
13/13 7/7 16/16 |
3/4 1/2 4/5 |
6/5 6/5 11/10 |
4/4 0/0 1/1 |
13/13 7/7 16/16 |
|
Таблица 2.
Число моляров (экз.) для двух составляющих изменчивости жевательной поверхности М1 и группирующих факторов
| Возраст, мес. |
Пол | Фазы цикла | Категории складчатости | Категории числа замкнутых полей | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | всего | 1 | 2 | 3 | всего | |||
| 10–12 | Самцы | Спад Подъем Пик |
2/2 3/3 9/14 |
0/0 1/1 12/7 |
3/3 0/0 10/10 |
5/5 4/4 31/31 |
0/0 2/2 3/2 |
3/3 1/1 19/20 |
2/2 1/1 9/9 |
5/5 4/4 31/31 |
| Самки | Спад Подъем Пик |
1/1 1/1 7/6 |
1/1 0/1 11/10 |
1/1 1/0 11/13 |
3/3 2/2 29/29 |
0/0 1/2 1/1 |
1/1 1/0 22/22 |
2/2 0/0 6/6 |
3/3 2/2 29/29 |
|
| 13–15 | Самцы | Спад Подъем Пик |
5/5 11/10 16/14 |
4/4 5/5 8/12 |
2/2 2/3 7/5 |
11/11 18/18 31/31 |
0/0 3/3 7/4 |
6/6 11/10 17/23 |
5/5 4/5 7/4 |
11/11 18/18 31/31 |
| Самки | Спад Подъем Пик |
7/7 3/4 7/10 |
2/3 1/1 5/5 |
4/3 3/2 5/2 |
13/13 7/7 17/17 |
1/1 0/1 1/1 |
6/5 5/4 9/6 |
6/7 2/2 7/10 |
13/13 7/7 17/17 |
|
Рис 1.
Морфотипы складчатости третьего верхнего (1A–1D) и первого нижнего (2A–2C) моляров. Цифры со стрелками – номера точек разделения дентинового поля.
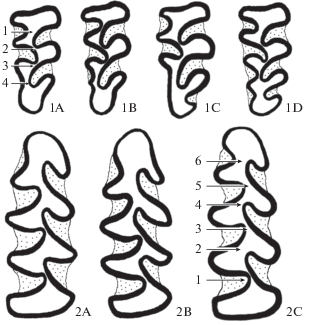
Выделяли по три категории в каждой составляющей изменчивости. К первой категории складчатости относили наименее сложные моляры: с тремя буккальными (BSA) и тремя лингвальными (LSA) выступающими углами (3/3, 1A) для M3 и с четырьмя BSA и четырьмя LSA (4/4, 2A) для M1. Во вторую категорию включали моляры с семью SA (4/3, 1B и 3/4, 1C) для M3 и со слабо выраженным 5LSA для M1 (2B). В третью категорию попадали моляры с восемью SA (4/4, 1D) для M3 и хорошо выраженным 5LSA для M1 (2C). По числу замкнутых полей моляры делили на категории: первая – с одним-двумя замкнутыми полями, вторая – с тремя полями, третья – с четырьмя-пятью замкнутыми полями. При статистической обработке данных использовали пакет программ StatSoft STATISTICA for Windows 6.0: логлинейный анализ, статистику χ2, ранговую корреляцию Спирмена – RS, множественный регрессионный анализ, регрессионный коэффициент B.
Когда данные представлены в виде многомерных (многофакторных) таблиц частот, логлинейный анализ позволяет оценить влияние факторов (объясняющих или независимых переменных), присутствующих в таблице сопряженности, на изучаемый параметр – переменную отклика или зависимую переменную. Взаимодействие факторов в их влиянии на переменную отклика также может быть оценено. Логлинейный анализ позволяет представить многомерную таблицу частот как отражение главных и взаимодействующих влияний, которые складываются линейным образом. Статистическая значимость влияний оценивается с помощью статистики χ2 [6].
Динамика численности популяции рыжей полевки. За период исследований наблюдали шесть полных циклов: четыре трехлетних, один двухлетний и один четырехлетний. Двухлетний цикл состоит из года средней и высокой численности. Четырехлетний цикл представлен тремя годами низкой численности и пиковым годом. В трехлетних циклах годы низкой (спад), средней (подъем) и высокой (пик) численности следуют друг за другом [1].
Изменчивость жевательной поверхности M3 моляра. Самые сложные по складчатости M3 моляры (4/4) наименее многочисленны (11% от n = 338), наименее сложные встречаются в 41% случаев, а преобладают (48%) моляры второй категории складчатости (3/4 – 45% и 4/3 – 3%). Доминируют моляры с тремя замкнутыми дентиновыми полями (55%), с одним-двумя полями встречаются реже (30%), с четырьмя-пятью полями – самые малочисленные (15%). Разделение 2 встречается в 91% из 307 моляров, далее идут разделение 3 (72%), 4 (13%), 1 (8%) и отсутствие разделений (3%). Моляры всех трех категорий сложности для обоих признаков встречаются у полевок, достигших возраста около 15 мес.
Факторы, влияющие на изменчивость изучаемых показателей, оценивали с помощью логлинейного анализа. Положение в челюсти (правая и левая части) не влияет на изменчивость всех изучаемых признаков (p > 0.42). Если изменчивость числа замкнутых полей не связана с возрастом как для левых и правых моляров, так и для объединенной выборки (χ2 < 7.3, df = 6, p > 0.29), то частота первой категории складчатости повышается с возрастом (χ2 > 12.7, df = 2, p < 0.002): в возрасте 10–12 мес. она составляет 31% (левые-s – 31%, правые-d – 31%), а в 13–15 мес. достигает 48% (s – 51%, d – 46%). Подобные изменения наблюдали на выборках из других популяций этого вида [3, 7].
Изменчивость числа замкнутых полей меняется в зависимости от фазы популяционного цикла статистически значимо (χ2 = 21.6, df = 4, p < 0.001). Для односторонних выборок (правая либо левая сторона челюсти) эта связь также статистически значима (χ2 > 11.8, df = 4, p < 0.02). При низкой численности доля моляров третьей категории выше (35%, s – 32%, d – 39%), а первой – ниже (16%, s – 16%, d – 16%), чем при средней и высокой – соответственно 11% (s – 12%, d – 9%) и 11% (s – 10%, d – 11%), 41% (s – 41%, d – 41%) и 30% (s – 31%, d – 29%). Логлинейный анализ показал, что ни пол, ни возраст, ни сторона челюсти не изменяют характер этой зависимости, и она остается отрицательной (для трехфакторных взаимодействий χ2 < 4.8, df = 4, p > 0.30).
Из результатов логлинейного анализа следует, что для складчатости связь с фазами популяционного цикла более сложная. Она проявляется при взаимодействии либо с полом (χ2 = 5.5, df = 1, p = 0.02), либо с возрастом (χ2 = 8.6, df = 1, p < 0.01).
У самок доля моляров первой категории складчатости при низкой численности выше (56%, s – 56%, d – 56%, χ2 > 4.5, df = 1, p < 0.04), чем при средней и высокой вместе взятых (25%, s – 22%, d – 28%). У самцов доля моляров этой категории меняется от низкой численности (47%, s – 53%, d – 40%) к средней и высокой вместе взятых (47%, s – 46%, d – 48%) статистически незначимо (χ2 < 0.3, df = 1, p > 0.58). Эти же результаты могут быть представлены другим образом: различия в пропорциях категорий складчатости между самцами и самками при низкой численности отсутствуют, а при средней и высокой вместе взятых они выражены и заключаются в большей доле сложных моляров у самок (3/4), чем у самцов (1/2).
У полевок в возрасте 13–15 мес. частота моляров первой категории складчатости меняется по фазам популяционного цикла статистически незначимо – от 46% (s – 48%, d – 42%) до 51% (s – 52%, d – 50%, χ2 < 0.5, df = 2, p > 0.77). В возрасте 10–12 мес. эта частота при низкой численности выше (69%, s – 75%, d – 63%, χ2 = >4.1, df = 1, p < 0.04), чем при средней и высокой вместе взятых – 27% (s – 26%, d – 27%).
При низкой численности связь с возрастом доли моляров первой категории складчатости отрицательная (RS = –0.27, p = 0.03) по сравнению с другими фазами популяционного цикла, когда она положительная (RS = 0.37, p = 0.003 и RS = 0.25, p = 0.001). Это могло произойти в результате действия избирательной элиминации в постнатальный период в пользу упрощенной категории складчатости при низкой численности среди полевок, имеющих возраст 10–12 мес. Сбалансированный полиморфизм складчатости M3 моляра у рыжей полевки трактуется как проявление двухаллельной системы с неполным доминированием [8, 9]. Видимо, рыжие полевки при низкой численности проявляют большую степень зеленоядности, чем при более высокой. В изучаемой популяции более высокая численность рыжих полевок чаще сочетается с большими урожаями шишек ели, являющейся основным лесообразующим элементом [1]. В годы низкой продукции семян лесообразующих деревьев в желудках рыжих полевок плодокорм встречается реже (19%), чем в годы высокой урожайности (70%) [10].
Изменчивость жевательной поверхности M1 моляра. Моляры первой категории складчатости встречаются наиболее часто (44% от n = 342), второй и третьей категорий с одинаковой частотой – 27% и 29%. Моляры с тремя замкнутыми полями преобладают (59%), с четырьмя-пятью встречаются реже (30%), с одним-двумя наиболее малочисленные (11%). Чаще (в 95 и 90% случаев) встречаются разделения 1 и 3, далее идут разделение 4 (19%), 5 (13%), отсутствие разделений (3%) и разделение 2 (2%). Разделение 6 и моляры с более чем четырьмя разделениями не обнаружены. Моляры всех трех градаций сложности для обоих признаков встречаются у полевок вплоть до возраста 15 мес.
Как и для M3 моляра, изменчивость числа замкнутых дентиновых полей для M1 моляра не связана с возрастом как для левых и правых моляров, так и для объединенной выборки (χ2 < 2.1, df = 2, p > 0.34). Для категорий складчатости M1 моляра такая связь присутствует. Частота моляров первой категории складчатости повышается с возрастом (χ2 = 13.0, df = 2, p < 0.002): в возрасте 10–12 мес. она составляет 34% (s – 31%, d – 36%), а в 13–15 мес. эта частота достигает 51% (s– 51%, d– 52%). Подобные изменения наблюдали на выборках из других популяций этого вида [3].
Изменчивость числа замкнутых полей M1 моляра меняется в зависимости от фазы популяционного цикла статистически значимо (χ2 = 21.5, df = 4, p < 0.001). Доля моляров с четырьмя-пятью замкнутыми полями при низкой численности выше (48%), чем при средней (24%) и высокой (27%). Для односторонних выборок эта связь также статистически значима (χ2 > 5.5, df = 1, p < 0.02). Изменение некоторых элементов жевательной поверхности, связанных с числом замкнутых полей на M1 моляре, в зависимости от динамики численности наблюдали у рыжей полевки [11].
Для категорий складчатости связь с фазами популяционного цикла отсутствует (χ2 < 6.1, df = 4, p > 0.19). На выборках, состоящих из прибылых особей, такую связь наблюдали [3].
Взаимосвязь между составляющими изменчивости жевательной поверхности M3 и M1 моляров между собой и другими факторами. Для установления этой взаимосвязи мы использовали множественную регрессию. Когда складчатость M3 моляра выступает в роли зависимой переменной, только пол (B = 0.21 ± 0.07, p < 0.01) и возраст (B = –0.19 ± 0.08, p < 0.02) из всего набора независимых переменных статистически значимо влияют на нее, а фазы популяционного цикла влияют на уровне тенденции (B = 0.08 ± 0.05, p = 0.07). Для складчатости M1 моляра, числа замкнутых полей M3 и M1 моляров и стороны челюсти B < 0.05 ± 0.04, p > 0.29.
Если в роли зависимой переменной выступает число замкнутых полей M3 моляра, то статистически значимые связи наблюдаются с фазами популяционного цикла (B = –0.15 ± 0.04, p < 0.001) и числом замкнутых полей M1 моляра (B = 0.37 ± 0.06, p < 0.001), а на уровне тенденции – с возрастом (B = –0.13 ± 0.07, p = 0.07). Для складчатости M3 и M1 моляров, пола и стороны челюсти B < 0.07 ± 0.04, p > 0.18.
При использовании складчатости M1 моляра в роли зависимой переменной обнаружены статистически значимые связи с возрастом (B = – 0.27 ± 0.09, p < 0.01) и числом замкнутых полей M1 моляра (B = 0.17 ± 0.08, p < 0.03). Для складчатости и числа замкнутых полей M3 моляра, фаз популяционного цикла, пола и стороны челюсти B < 0.10 ± 0.06, p > 0.18.
При использовании числа замкнутых полей M1 моляра в роли зависимой переменной обнаружены статистически значимые связи с числом замкнутых полей M3 моляра (B = 0.32 ± 0.05, p < 0.001), со складчатостью M1 моляра (B = 0.09 ± 0.04, p < 0.03) и полом (B = 0.13 ± 0.06, p < 0.05). Для складчатости M3 моляра, фаз популяционного цикла, возраста и стороны челюсти B < 0.07± 0.04, p > 0.18. Если исключить из числа независимых переменных число замкнутых полей M3 моляра, то связь с фазами популяционного цикла становится статистически значимой (B = –0.10 ± 0.04, p < 0.03).
В целом результаты множественного регрессионного анализа хорошо согласуются с данными логлинейного анализа. Кроме того, они позволили установить, что изменчивость складчатости M3 моляра не связана с изменчивостью ни числа замкнутых полей этого же моляра, ни с одной из составляющих изменчивости жевательной поверхности M1 моляра. В то же время между числом замкнутых полей M3 и M1 моляров, складчатостью и числом замкнутых полей M1 моляра наблюдается взаимосвязь.
Таким образом, впервые у перезимовавших особей рыжей полевки изучена изменчивость числа замкнутых дентиновых полей М3 и М1 моляров в зависимости от фаз динамики численности, возраста, пола и положения в челюсти. Обнаружено ранее неизвестное, ярко выраженное явление, состоящее в увеличении при низкой численности популяции доли M3 и M1 моляров с повышенным числом замкнутых дентиновых полей на жевательной поверхности. Вероятно, это явление возникает как приспособление к большей степени зеленоядности в данный период. Оно могло возникнуть в результате действия избирательной элиминации отдельных особей. На представителях Arvicolinae из лабораторных колоний было показано, что число замкнутых полей – наследственно детерминированный признак [12, 13]. Известно также, что более зеленоядная красно-серая полевка (Cl. rufocanus) отличается большим числом замкнутых полей на жевательной поверхности M3 и M1 моляров [14].
Следовательно, у замкнутых дентиновых полей М3 и М1 моляров не выражена возрастная изменчивость в пределах перезимовавших особей, в то время как складчатость M3 и M1 моляров подвержена этой изменчивости. Последнее было известно ранее и подтверждено нами.
Работа выполнена в рамках государственного задания № АААА-А19-119031890087-7 Института экологии растений и животных УрО РАН.
Список литературы
Чепраков М.И. Изменчивость демографических характеристик рыжей полевки (Clethrionomys glareolus) в циклической популяции // Экология. 2018. № 4. С. 312–317. [Cheprakov M.I. Variability of demographic parameters in a cyclic population of the bank vole (Clethrionomys glareolus) // Rus. J. Ecol. 2018. V. 49. № 4. P. 343–348.]
Окулова H.М., Андреева Т.А. Межвидовая и внутривидовая дифференциация лесных полевок рода Clethrionomys (Rodentia, Cricetidae) по данным изменчивости жевательной поверхности зуба М3 // Зоол. журн. 2008. Т. 87. Вып. 8. С. 991–1003.
Zejda J., Zukal J., Steiner H.M. Variation in the shape pattern of molars in populations of the bank vole (Clethrionomys glareolus Schreb.) // Fol. Zool. 1994. V. 43. № 2. P. 121–133.
Оленев Г.В. Определение возраста цикломорфных грызунов, функционально-онтогенетическая детерминированность, экологические аспекты // Экология. 2009. № 2. С. 102–115. [Olenev G.V. Determining the age of cyclomorphic rodents: functional-ontogenetic determination, ecological aspects // Rus. J. Ecol. 2009. V. 40. № 2. P. 93–104.]
Markova E.A., Malygin V.M., Montuire S. et al. Dental variation in sibling species Microtus arvalis and M. rossiaemeridionalis (Arvicolinae, Rodentia): between-species comparisons and geography of morphotype dental patterns // J. Mammal. Evol. 2010. V. 17. P. 121–139.
Bishop Y.M., Fienberg S.E., Holland P.W. Discrete multivariate analysis: theory and practice. New York: Springer Science et Business Media, 2007. 559 p.
Zejda J. The influence of age on the formation of the third upper molar in the bank vole (Clethrionomys glareolus Shreber, 1780) (Mammalia: Rodentia) // Fol. Zool. 1960. V. 9. № 2. P. 159–166.
Steven D.M. Untersuchungen über die britischen formen von Clethrionomys // Z. Saugetierk. 1955. Bd 20. H 2–3. S. 70–74.
Bauchau V., Chaline J. Variabilité de la troisième molaire supérieure de Clethrionomys glareolus (Arvicolidae, Rodentia) et sa signification évolutive // Mammalia. 1987. V. 51. № 4. P. 587–598.
Drozdz A. Food habits and food supply of rodents in the beech forest // Acta Theriol. 1966. V. 11. № 10–20. P. 363–384.
Фоминых М.А., Маркова Е.А., Бородин А.В., Давыдова Ю.А. Внутрипопуляционная изменчивость одонтометрических признаков рыжей полевки Myodes glareolus Schreber, 1780 на Среднем Урале // Экология. 2010. № 6. С. 468–471. [Fominykh M.A., Markova E.A., Borodin A.V., Davydova Yu. A. Intrapopulation variation in odontometric characters of the bank vole Myodes glareolus Schreber, 1780 in the Middle Urals // Rus. J. Ecol. 2010. V. 41. № 2. P. 93–104.]
Большаков В.Н., Васильева И.А., Малеева А.Г. Морфотипическая изменчивость зубов полевок. М.: Наука, 1980. 140 с.
Markova E., Smirnov N. Phenotypic diversity arising from a limited number of founders: A study of dental variation in laboratory colonies of collared lemmings, Dicrostonyx (Rodentia: Arvicolinae) // Biol. J. Linn. Soc. 2018. V. 125. P. 777–793.
Громов И.М., Поляков И.Я. Фауна СССР. Млекопитающие. Т. 3. Вып. 8. Л.: Наука, 1977. 502 с.
Дополнительные материалы отсутствуют.