

Экология, 2020, № 5, стр. 341-353
Межгодовая изменчивость качества семенного потомства пустырника пятилопастного (Leonurus quinquelobatus) в условиях хронического облучения
Е. В. Антонова a, *, В. Н. Позолотина a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: selena@ipae.uran.ru
Поступила в редакцию 03.02.2020
После доработки 06.03.2020
Принята к публикации 27.04.2020
Аннотация
Исследована восьмилетняя динамика качества семенного потомства пустырника пятилопастного (Leonurus quinquelobatus Gilib.), произрастающего более 60 лет в условиях хронического облучения в зоне Восточно-Уральского радиоактивного следа (ВУРС), в сравнении с фоновыми ценопопуляциями. Мощности поглощенных доз материнских растений в зоне ВУРСа превышают фоновый уровень на 1–3 порядка величин. Не обнаружено однозначной зависимости между дозовой нагрузкой материнских растений и качеством их семенного потомства. Ключевую роль в формировании семян пустырника играют погодные факторы, а также сочетанное действие метеоусловий и хронического облучения. Все исследованные характеристики качества семенного потомства пустырника из зоны ВУРСа позитивно коррелировали с суммой эффективных температур за апрель, т.е. с тепловым режимом в начале вегетации растений. Основными метеорологическими факторами, влиявшими на физиологический отклик (жизнеспособность, мутабильность, радиочувствительность) пустырника из фоновой и импактной зон, являются индекс Селянинова за апрель текущего сезона и сумма осадков за ноябрь предыдущего года. При этом по индексу Селянинова для фоновых и импактных выборок отмечены отрицательные корреляции по жизнеспособности и положительные – по мутабильности семенного потомства. Физиологический отклик на метеоусловия, оцененный по радиоустойчивости семян, в фоновых выборках был позитивный, а в импактных – негативный. Зависимости между суммой осадков за ноябрь прошлого года и качеством семян в фоновых выборках были положительные, а в импактных – отрицательные.
Природные экосистемы существуют в изменчивых погодных условиях. Дополнительная антропогенная нагрузка создает повышенное напряжение в экосистемах и порождает разные ответы организмов на сочетанные внешние воздействия: синергизм, антагонизм или индифферентные реакции [1–3]. Травянистые растения широко распространены, ведут прикрепленный образ жизни и используются в качестве “референтных организмов” для оценки отдаленных последствий действия ионизирующей радиации на природные экосистемы [4]. Поскольку популяции являются основными элементарными единицами в микроэволюционных процессах [5], система защиты окружающей среды от действия радиации должна быть ориентирована на популяции и экосистемы [4, 6–10].
Многолетний мониторинг за состоянием популяций растений, произрастающих на загрязненных территориях, выявил изменчивость качества семенного потомства у разных видов, связанную с флуктуацией погодных условий, а также с взаимодействием факторов радиационной и нерадиационной природы [3, 11–16]. Однако остается не ясным, какие погодные факторы являются наиболее значимыми для формирования семенного потомства растений, произрастающих на техногенно-нарушенных территориях. Такие предикторы могли бы служить маркерами успешности существования популяции во времени и пространстве.
Цель данного исследования – изучение в динамике качества семенного потомства пустырника пятилопастного (Leonurus quinquelobatus), произрастающего в зоне радиоактивного загрязнения и фоновых условиях. Мы предположили, что для мезофита и многолетнего поликарпика L. quinquelobatus: 1) качество семенного потомства снижается в условиях избыточного увлажнения и низких температур; 2) погодные условия осенне-зимнего периода, предшествующего новой вегетации, оказывают значимое влияние на качество семенного потомства, при этом осадкам отводится доминирующая роль; 3) фоновые и импактные популяции характеризуются разными физиологическими откликами при действии одного и того же климатического фактора.
МАТЕРИАЛ И МЕТОДЫ
Характеристика района исследования. Восточно-Уральский радиоактивный след (ВУРС) сформировался в результате Кыштымской аварии на предприятии “Маяк” в 1957 г., главным долгоживущим радионуклидом является 90Sr, загрязняющий территорию до сих пор – плотность загрязнения почв в ближней к эпицентру зоне достигает 70 000 кБк/м2 (фоновый уровень 1.5−3.0 кБк/м2) и уменьшается с расстоянием от эпицентра взрыва согласно экспоненциальной функции [17, 18]. В 1967 г. зона ВУРСа была загрязнена повторно путем переноса радиоактивного ила и песка с берегов обмелевшего оз. Карачай, которое использовалось предприятием в качестве открытого хранилища радиоактивных отходов. Основным загрязнителем был 137Cs. В настоящее время ВУРС является уникальным полигоном для радиоэкологических исследований, поскольку живые организмы в зоне более 60 лет подвергаются хроническому воздействию малых доз радиации [13, 17–20].
На территории ВУРСа были выделены условно импактная и буферная части: первая расположена в пределах центральной оси на расстоянии 4–18 км от эпицентра аварии, вторая – на западной, северной и восточной перифериях зоны [20, 21]. На рис. 1 показано расположение четырех импактных площадок: южный берег оз. Урускуль (имп-1), берег оз. Бердениш (имп-2), лежневая дорога вдоль южной границы ВУРСа (имп-3), просека на южной границе ВУРСа (имп-4), и 2 буферных: западная граница ВУРСа (буф-1), юго-западный берег оз. Урускуль (буф-2). Географические координаты площадок и мощности поглощенных доз материнских растений пустырника приведены в табл. 1. На всех площадках произрастают злаковые и разнотравно-злаковые сообщества [18]. Почвы преимущественно серые лесные разных подтипов и дерново-подзолистые, на площадке импакт-2 на месте отселенной деревни встречается чернозем среднемощный среднесуглинистый. Подробная характеристика почв и растительных сообществ представлена в монографии [20].
Рис. 1.
Расположение точек отбора проб в пределах радиоактивно-загрязненной зоны и фоновой территории: Рассоха (фон-1), Шеелит (фон-2), биостанция УрФУ (фон-3), лагерь Гагарина (фон-4), западная граница ВУРСа (буф-1), юго-западный берег оз. Урускуль (буф-2), южный берег оз. Урускуль (имп-1), берег оз. Бердениш (имп-2), лежневая дорога вдоль южной границы ВУРСа (имп-3), просека на южной границе ВУРСа (имп-4). См. табл. 1.
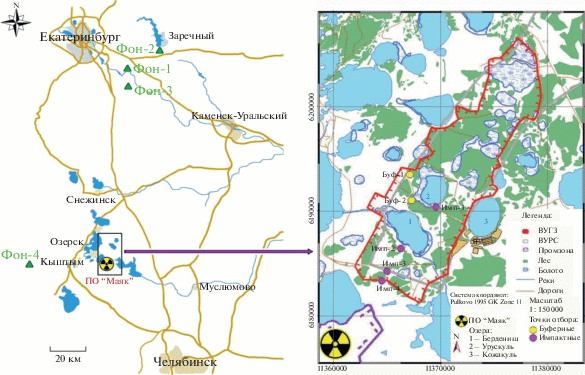
Таблица 1.
Характеристика радиационной обстановки на изученных участках произрастания материнских растений L. quinquelobatus
| Участок | Символ | GPS координаты | Расстояние от эпицентра аварии, км | Мощность дозы, мкГр/ч*** | Кратность увеличения дозы |
|---|---|---|---|---|---|
| Рассоха* | Фон-1 | 56°42′ N; 61°02′ E | 112 | 0.10174 | 1 |
| Шеелит* | Фон-2 | 56°47′ N; 61°17′ E | 125 | 0.10353 | |
| Биостанция УрФУ** | Фон-3 | 56°36′ N; 61°3′ E | 100 | 0.10303 | |
| Лагерь Гагарина** | Фон-4 | 55°37′ N; 60°27′ E | 25 | 0.10283 | |
| Среднее фоновое значение | 0.1028 | ||||
| Западная граница ВУРСа* | Буф-1 | 55°50′ N; 60°52′ E | 13–15 | 0.221 | 2.2 |
| Юго-западный берег оз. Урускуль* | Буф-2 | 55°49′ N; 60°54′ E | 14–15 | 4.06 | 39.5 |
| Южный берег оз. Урускуль** | Имп-1 | 55°49′ N; 60°53′ E | 15–16 | 12.1 | 117.7 |
| Южный берег оз. Бердениш* | Имп-2 | 55°46′ N; 60°51′ E | 10–12 | 15.2 | 147.9 |
| Лежневая дорога вдоль южной границы ВУРСа* | Имп-3 | 55°45′ N; 60°50′ E | 6–8 | 32.5 | 316.2 |
| Просека на южной границе ВУРСа** | Имп-4 | 55°45′ N; 60°50′ E | 4–5 | 70.2 | 682.9 |
* Данные приведены в работе [19]. ** Результаты текущего исследования. *** В оценку включено среднее значение мощности дозы естественного радиационного фона для Уральского региона (~0.1 мкГр/ч).
Фоновые площадки были выбраны в сходных условиях, вне зоны какого-либо загрязнения и на удалении от автомобильных дорог (см. рис. 1). Населенный пункт Рассоха, бывший пос. Шеелит и биостанция Уральского федерального университета (УрФУ) расположены в Свердловской области, в подзоне южной тайги [22]. Тип сообществ оценивается как вторичный суходольный разнотравно-злаковый луг [18, 23]. На участках доминировали дерново-подзолистые суглинистые и серые лесные суглинистые почвы [20]. Фоновая площадка около детского лагеря им. Ю.А. Гагарина расположена в подзоне Южно-Зауральской провинции. Растительное сообщество представлено смешанным сосново-березовым лесом. В регионе преобладает тип почв подбуры темные таежные [24].
Объект исследования. Пустырник пятилопастной (Leonurus quinquelobatus Gilib.) – многолетнее травянистое растение из сем. Lamiaceae, диплоид (2n = 18), рудеральный вид, поликарпик, мезофит [25]. Смеси семян L. quinquelobatus были собраны в 2006–2018 гг. с 50–70 растений в каждой ценопопуляции.
Определение радионуклидов в образцах почв и растений. Содержание изотопов 90Sr и 239,240Pu в почвах и растениях определяли апробированными радиохимическими методами [19, 21], концентрации 137Cs в образцах – с помощью многоканального гамма-анализатора с полупроводниковым детектором Canberra-1510 (Canberra-Packard, USA). Радиохимический анализ был проведен в лаборатории общей радиоэкологии Отдела континентальной радиоэкологии ИЭРиЖ УрО РАН (сертификат САРК RU.0001.441492).
Расчет поглощенных доз для пустырника был выполнен с помощью ERICA Assessment Tool 1.2. Использованы эмпирические данные о содержании радионуклидов (90Sr, 137Cs, 239,240Pu) в компонентах экосистем (почвы, наземная масса растений), о коэффициентах перехода в системе почва–растение, отношениях сырой и сухой массы растений и некоторые другие параметры. Этапы расчетов изложены в работе [19].
Погодные условия в разные годы были оценены с использованием среднемесячных температур, суммы температур за каждый месяц, суммы эффективных температур, суммы осадков за каждый месяц, суммы осадков при эффективных температурах, индекса аридности (IA) по Селянинову. Эффективные температуры – это среднесуточные температуры выше +10°C. Индекс Селянинова рассчитывали по формуле
где IA – индекс Селянинова, P – сумма осадков при эффективных температурах за изучаемый период, ΣTn – сумма эффективных температур за тот же период [26]. Метеорологические данные были взяты из двух локальных метеостанций, расположенных вблизи фоновых ценопопуляций (№ 28 440) и вблизи зоны ВУРСа (№ 28541). В анализе использовано 184 комбинаций погодных условий текущего и предыдущего сезонов.Лабораторные эксперименты и параметры оценки качества семян. Смесь семян высушивали, очищали, хранили в холодильнике при +5°C до следующего года. В физиологически благоприятный период семена ежегодно проращивали по 25 или 50 шт. в 4 повторностях на каждый вариант. Проращивание проводили в рулонной культуре на дистиллированной воде при +24°C, 12-часовом световом режиме в течение 3 недель. Ежедневно сосуды с проростками рандомизировали на стеллажах и доливали в них воду. Для оценки радиоустойчивости проростков семена облучали перед посевом гамма-лучами в дозах 25, 50, 100, 150, 200, 250, 300, 350, 400 Гр, используя γ-установку “Игур” (мощность источника 137Cs 38–41 Гр/с). Всего в экспериментах использовано 29 360 семян.
Для оценки жизнеспособности семенного потомства использовали следующие параметры: всхожесть, выживаемость, доля проростков с настоящими листьями, длина главного и наличие латеральных корней. Мутабильность оценивали по частоте встречаемости проростков с морфозами (некроз или изменение формы и цвета семядолей, листьев, корней, нарушение гелиотропизма). Адаптивный потенциал (радиоустойчивость) семенного потомства оценивали, сравнивая параметры жизнеспособности в вариантах с острым облучением и без него, выражая значения в % к необлученному контролю. Всего для оценки использовано 172 физиологических отклика.
Анализ данных. На начальном этапе с помощью линейного корреляционного анализа и кластерного анализа была проведена проверка на скоррелированность комбинаций погодных данных, а также биологических параметров. Значения коэффициентов корреляции считали значимыми при уровне р < 0.01. Средние температуры за месяц коррелировали с эффективными температурами и суммой температур, поэтому для анализа учитывали только эффективные температуры как наиболее физиологически значимые для растений. Кроме того, для анализа погодных условий использовали сумму осадков за месяц и индекс аридности Селянинова, а также такие соотношения факторов, как сумма осадков при эффективных температурах за определенные периоды. Физиологические параметры (всхожесть семян, выживаемость проростков, листообразование) коррелировали между собой. В дальнейшем анализировали показатель выживаемости проростков на стадии листообразования, т.е. долю проростков, сформировавших к 21-му дню настоящие листья, в % от числа выживших. Этот показатель имеет прогностическое значение, поскольку характеризует работу верхушечных меристем. Кроме того, использовали нескоррелированный с другими характеристиками показатель “длина корней”, оценивающий работу корневых меристем.
Зависимость параметров жизнеспособности, мутабильности и радиоустойчивости (y) от погодных условий (x) описывалась уравнением линейной регрессии y = b0 ± b1x. Разность независимых переменных, выраженных в долях, оценивали с помощью метода сравнения верхних и нижних границ 95%-ных доверительных интервалов (метод сравнения пропорций) [27]. Кроме того, для проверки статистических гипотез были использованы одно- и многофакторный дисперсионный анализ и тест Данна в программе STATISTICA 10.0.
РЕЗУЛЬТАТЫ
Оценка мощностей поглощенных доз
Мощности поглощенных доз (МПД) материнских растений пустырника в зоне ВУРСа варьировали: 0.22–4.0 мкГр/ч – в буферной зоне и 12–70 мкГр/ч – в импактной (см. табл. 1). Эти величины превышают фоновый уровень мощности дозы в Уральском регионе (~0.1 мкГр/ч) на 1–3 порядка величин, но не выходят за пределы МПД 70 мкГр/ч, принятой за критический уровень для референтной группы “растения”, после которой можно ожидать появления радиобиологических эффектов [9].
Изменчивость показателей жизнеспособности семенного потомства пустырника, обусловленная хроническим облучением и переменными погодными условиями
Метод сравнения пропорций показал, что в 2006 г. выживаемость проростков на стадии листообразования из зоны ВУРСа была значимо выше, а в 2009 и 2010 гг. – значимо ниже, чем в фоновых выборках (рис. 2). В 2008 г. вариабельность выживаемости проростков в фоновых популяциях была высока, сравнение семенного потомства пустырника из облучаемых выборок со средним фоновым значением не выявило различий, хотя с выборкой фон-2 они были значимы. В 2011–2018 гг. вариабельность выживаемости проростков была высокой в импактных выборках. В большинстве случаев, когда различия между импактными и фоновыми выборками были значимы, популяции из зоны ВУРСа обладали более низкой выживаемостью (исключение – выборки 2018 г.).
Рис. 2.
Изменчивость выживаемости проростков пустырника на стадии листообразования (1 – % к общему числу выживших) и длины их корней (2 – мм, вспомогательная шкала) в градиенте загрязнения ВУРСа. На каждом рисунке указан уровень значимости коэффициентов линейной регрессии. Звездочками обозначены значимые попарные сравнения между показателями выживаемости (метод сравнения пропорций) и длины корней (тест Данна) проростков пустырника из облучаемых и фоновых выборок.
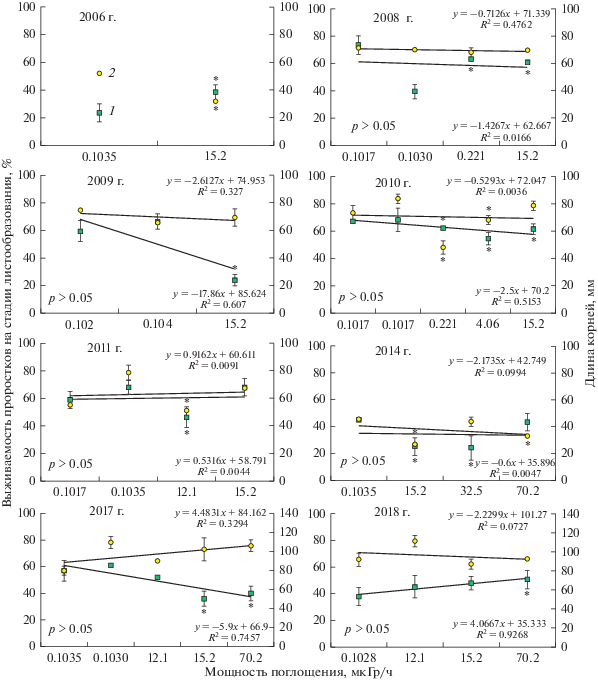
Влияние факторов “вегетационный сезон (год)” и “участок (дозовая нагрузка)” на качество семенного потомства оценили с помощью двухфакторного дисперсионного анализа (табл. 2). Оба фактора, а также их взаимодействие вносят значимый вклад в изменчивость выживаемости проростков на стадии листообразования. При этом в однофакторном приближении для ценопопуляций пустырника из зоны ВУРСа погодные условия имели большое значение (F6;8 = 5.24, p = 0.018), а для фоновых выборок значимых различий не обнаружено (F7;8 = 0.721, p = 0.66). По длине корней значимыми оказались фактор “вегетационный сезон (год)”, а также двухфакторный эффект.
Таблица 2.
Результаты дисперсионного анализа влияния вегетационного сезона (1) и радиационной обстановки на участках произрастания материнских растений (2) на выживаемость и длину корней проростков пустырника
| Фактор | Выживаемость | Длина корня | ||||
|---|---|---|---|---|---|---|
| df | F | p | df | F | p | |
| (1) | 8 | 6.11 | ⪡0.0001 | 8 | 18.39 | ⪡0.0001 |
| (2) | 15 | 3.43 | ⪡0.0001 | 15 | 1.48 | 0.112 |
| (1) × (2) | 12 | 7.67 | ⪡0.0001 | 12 | 3.31 | 0.0002 |
Изменчивость качества семенного потомства с учетом влияния погодных условий была показана и для других видов травянистых растений из зоны ВУРСа: звездчатки злаковой, дремы белой, костреца безостого и одуванчика лекарственного [3, 11–13, 16]. Разберем детально, какие именно погодные факторы определяли качество семян пустырника (табл. 3). В фоновых популяциях обнаружено только два значимых эффекта влияния погодных условий: 1) отрицательная (–) корреляция между индексом Селянинова за апрель и длиной корня; 2) отрицательная (–) корреляция между суммой осадков за ноябрь прошлого года и длиной корня. В импактных популяциях на выживаемость проростков пустырника на стадии листообразования оказывали значимое влияние следующие факторы: 1) сумма эффективных температур за апрель и июль (+); 2) сумма осадков при эффективных температурах за апрель и август (–) текущего года; 3) сумма осадков за май (+) и август (–); 4) индекс Селянинова за апрель (–) и май (+) текущего и октябрь (–) прошлого сезона.
Таблица 3.
Значения коэффициентов корреляции и анализ уравнений линейной регрессии y = b0 ± b1x для показателей жизнеспособности семенного потомства пустырника из разных зон
| Показатель | Климатический фактор | R | p-значение | b0 ± SE | b1 ± SE |
|---|---|---|---|---|---|
| Фоновые популяции | |||||
| Длина корня | Selyaninov-4 | –0.73 | 0.0028 | 68.83 ± 5.18 | –20.8 ± 5.56 |
| Sum_P-11 | –0.69 | 0.003 | 76.03 ± 6.13 | –0.18 ± 0.05 | |
| Импактные популяции | |||||
| Выживаемость на стадии листообразования | Sum_P-5 | 0.67 | 0.0058 | 31.47 ± 9.47 | 0.55 ± 0.17 |
| Sum_P-8 | –0.69 | 0.0043 | 80.96 ± 7.11 | –0.48 ± 0.14 | |
| Sum_Tef.-4 | 0.74 | 0.0056 | 37.64 ± 6.92 | 0.65 ± 0.19 | |
| Sum_Tef.-7 | 0.67 | 0.0059 | –2.6 ± 19.5 | 0.13 ± 0.04 | |
| Sum_P-Tef.-4 | –0.99 | 0.0051 | 120.9 ± 4.25 | –74.82 ± 5.36 | |
| Sum_P-Tef.-8 | –0.70 | 0.0039 | 78.9 ± 6.53 | –0.46 ± 0.13 | |
| Selyaninov-4 | –0.98 | 0.0005 | 82.5 ± 2.07 | –96.56 ± 9.33 | |
| Selyaninov-5 | 0.65 | 0.0085 | 43.9 ± 6.5 | 14.44 ± 4.67 | |
| Selyaninov-10 | –0.99 | 0.00001 | 79.7 ± 1.53 | –36.31 ± 2.78 | |
| Длина корня | Т_mean-4 | 0.66 | 0.0080 | 28.5 ± 12.8 | 10.8 ± 3.5 |
| Sum_P-3 | –0.88 | 0.00002 | 119.6 ± 8.86 | –1.84 ± 0.28 | |
| Sum_Tef.-7 | 0.69 | 0.0045 | –16.2 ± 24.2 | 0.165 ± 0.048 | |
| Pef._9-6/Tef._5-8 | –0.65 | 0.0082 | 98.6 ± 11.8 | –563.8 ± 180.8 | |
Примечание: Sum_Tef. – сумма эффективных температур, Sum_P – сумма осадков, Selyaninov – индекс аридности Селянинова, Sum_P-Tef. – суммы осадков при эффективных температурах. Числа, включенные в показатели, индексируют месяцы, в которые проявилось влияние погодных факторов. Погодные показатели октября (Selyaninov-10) и ноября (Sum_P-11) характеризуют предыдущий сезон.
Отметим, что проростки с 2–3 листьями у пустырника чаще формировались при высоких эффективных температурах за летний период. Установлено, что этот физиологический показатель отрицательно (–) коррелировал с соотношением осадки/температура за летние месяцы текущего вегетационного сезона и положительно (+), если к осадкам текущего вегетационного сезона добавляли осадки осенне-зимнего периода прошлого года.
Для показателей “длина корня” и “наличие латеральных корней” у проростков пустырника импактной зоны значимыми оказались следующие факторы: 1) средняя температура за апрель (+); 2) сумма эффективных температур за июль (+); 3) сумма осадков за март (–); 4) сумма осадков при эффективных температурах за август (–); 5) отношение сумм осадков за различные периоды (осень прошлого–лето нынешнего года) к суммам эффективных температур за текущий вегетационный сезон (–).
Таким образом, за восемь лет основные показатели жизнеспособности семенного потомства пустырника из зоны ВУРСа были ниже, чем в фоновых популяциях. Обнаружен основной фактор (индекс Селянинова за апрель), оказывающий значимое влияние на качество семенного потомства как в фоновых, так и импактных выборках. Значение коэффициентов b0 уравнения линейной регрессии для импактных выборок было сопоставимо с таковыми для фоновых (см. табл. 3). При этом значение b1 для хронически облучаемых выборок пустырника было в 4.6 раза выше, чем для фоновых ценопопуляций, следовательно, эффект влияния данного фактора для пустырника из зоны ВУРСа был сильнее. В целом с ростом количества осадков показатели жизнеспособности семенного потомства пустырника из импактных выборок в большинстве случаев снижались.
Радиоустойчивость семенного потомства пустырника и ее связь с погодными условиями
Практически все дозы облучения, использованные в эксперименте (25, 50, 100, 150, 200, 250, 300, 350, 400 Гр), действовали однонаправленно, поэтому между показателями радиоустойчивости установлена тотальная корреляция. Выживаемость проростков на стадии листообразования и длина корней наиболее адекватно характеризовали ответ растений на провокационное облучение. Значимые факторы, влияющие на физиологический отклик проростков пустырника после предпосевного облучения в широком диапазоне доз в фоновых и импактных популяциях, приведены в табл. 4.
Таблица 4.
Значения коэффициентов корреляции и анализ уравнений линейной регрессии y = b0 ± b1x для показателей радиоустойчивости семенного потомства пустырника из разных зон
| Параметры | Климатический фактор |
R | p-значение | b0 ± SE | b1 ± SE |
|---|---|---|---|---|---|
| Фоновые выборки | |||||
| Выживаемость на стадии листообразования | Т_mean-1 | 0.70–0.83 | <0.0024 | 31.60 ± 7.48 | 1.88 ± 0.51 |
| Т_mean-8 | 0.63 | 0.009 | –120.4 ± 47.1 | 7.63 ± 2.72 | |
| Sum_P-5 | 0.63–0.74 | <0.009 | –9.34 ± 4.44 | 0.14 ± 0.04 | |
| Sum_P-11 | 0.85–0.98 | <0.0001 | –6.33 ± 3.96 | 0.28 ± 0.03 | |
| Sum_Tef.-6 | (–0.63)–(–0.83) | <0.009 | 51.91 ± 9.49 | –0.09 ± 0.01 | |
| Sum_P-Tef.-10 | (–0.69)–(–0.72) | 0.008 | 36.02 ± 9.14 | –0.75 ± 0.23 | |
| Selyaninov-4 | 0.81–0.96 | <0.00009 | –3.51 ± 1.88 | 15.25 ± 2.01 | |
| Длина корня | Т_mean-1 | 0.68–0.80 | <0.0040 | 11.62 ± 2.05 | 0.69 ± 0.14 |
| Sum_P-5 | 0.65–0.67 | <0.0064 | –4.92 ± 2.29 | 0.07 ± 0.02 | |
| Sum_P-11 | 0.75–0.96 | <0.001 | –1.71 ± 2.86 | 0.09 ± 0.02 | |
| Sum_Tef.-6 | (–0.68)–(–0.82) | <0.0039 | 27.36 ± 4.82 | –0.05 ± 0.009 | |
| Sum_Tef.-8 | 0.68 | 0.0038 | –70.51 ± 22.6 | 0.15 ± 0.04 | |
| Sum_P-Tef.-10 | –0.77 | 0.002 | 25.43 ± 4.97 | –0.53 ± 0.13 | |
| Selyaninov-4 | 0.70–0.93 | <0.0048 | –1.41 ± 0.69 | 6.15 ± 0.74 | |
| Selyaninov-9 | 0.79–0.95 | <0.006 | –14.8 ± 5.3 | 16.7 ± 4.5 | |
| Импактные выборки | |||||
| Выживаемость на стадии листообразования | Т_mean-1 | (–0.61)–(–0.92) | <0.00009 | –86.4 ± 12.2 | –5.93 ± 0.72 |
| Sum_P-4 | 0.67–0.68 | <0.0064 | –6.64 ± 3.28 | 0.26 ± 0.08 | |
| Sum_P-10 | (–0.66)–(–0.75) | <0.006 | 16.61 ± 5.76 | –0.35 ± 0.13 | |
| Sum_P-11 | –0.70 | 0.004 | 21.5 ± 7.0 | –0.41 ± 0.18 | |
| Sum_Tef.-4 | 0.75–0.92 | <0.005 | –11.75 ± 3.73 | 0.72 ± 0.99 | |
| Sum_Tef.-6 | 0.70 | 0.0033 | –137.3 ± 42.0 | 0.34 ± 0.09 | |
| Sum_P-Tef.-4 | –0.99 | 0.0063 | 79.67 ± 4.2 | –66.39 ± 5.3 | |
| Selyaninov-8 | –0.76 | 0.0009 | 39.53 ± 7.48 | –26.47 ± 6.25 | |
| Sum_P-Tef._5-8 | –0.65 | 0.008 | 54.4 ± 14.4 | –0.21 ± 0.68 | |
| Pef._10-8/Tef._5-6 | –0.67 | 0.0062 | 49.63 ± 12.28 | –123.6 ± 37.9 | |
| Pef._6-8/Tef._5-7 | –0.66 | 0.0076 | 46.28 ± 11.7 | –215.2 ± 68.3 | |
| Длина корня | Т_mean-1 | –0.70 | 0.0039 | –24.94 ± 8.29 | –1.72 ± 0.49 |
| Sum_P-4 | 0.62–0.68 | <0.0058 | –4.98 ± 2.47 | 0.19 ± 0.59 | |
| Sum_P-10 | –0.69 | 0.004 | 18.1 ± 4.52 | –0.35 ± 0.1 | |
| Sum_Tef.-4 | 0.90-0.92 | <0.00008 | –2.17 ± 0.71 | 0.13 ± 0.019 | |
| Sum_P-Tef.-4 | –0.99 | 0.0069 | 14.8 ± 1.27 | –12.3 ± 1.6 | |
| Selyaninov-4 | –0.94 | 0.0056 | 2.71 ± 0.21 | –5.1 ± 0.95 | |
| Selyaninov-10 | –0.94 | 0.005 | 2.57 ± 0.18 | –1.92 ± 0.34 | |
Примечание. Обозначения климатических факторов см. в табл. 3.
На выживаемость проростков фоновых ценопопуляций после провокационного облучения оказывали влияние следующие факторы: 1) средняя температура за январь и август (+); 2) сумма осадков за май (+) текущего и ноябрь прошлого сезона (+); 3) сумма эффективных температур за июнь (–); 4) сумма осадков при эффективных температурах за октябрь прошлого сезона (–); 5) индекс Селянинова за апрель (+). Указанные факторы оказывали аналогичное значимое влияние на длину корней, формирование 2 и более листьев, а также на рост латеральных корней. Кроме того, для этих показателей был значим фактор “сумма эффективных температур за август” (+). Позитивная корреляция отмечена также между длиной корней у проростков и индексом Селянинова за сентябрь прошлого года.
Значимое влияние на физиологический отклик (листообразование и длина корней) семенного потомства из импактных популяций оказывали следующие факторы: 1) средняя температура за январь (–); 2) сумма осадков за апрель (+) текущего, октябрь и ноябрь прошлого сезона (–); 3) сумма эффективных температур за апрель и июнь (+); 4) сумма осадков при эффективных температурах за апрель и весь вегетационный сезон (–); 5) индекс Селянинова за апрель, август и октябрь прошлого года (–); 6) соотношение осадки/температура как текущих, так предыдущих сезонов (–).
Значимые эффекты между соотношением осадки/температура как текущих, так и предыдущих сезонов и выживаемостью проростков импактной зоны имели одно и то же отрицательное направление. Это заключение подтверждают и показатели “доля проростков, имеющих два и более листа”, “наличие латеральных корней”.
Таким образом, для фоновых и импактных выборок обнаружено 4 общих погодных параметра (Т_mean-1, Sum_P-11, Sum_Tef.-6, Selyaninov-4), значимо влияющих на радиоустойчивость семенного потомства пустырника. Значения коэффициентов b0 уравнений линейной регрессии для импактных выборок были в 1.9–3.4, а коэффициентов b1 в 0.8–3.8 раза выше, чем на фоновых площадках. Следовательно, в целом влияние погодных факторов на формирование радиоустойчивости семян пустырника в выборках ВУРСа было сильнее. При этом для большинства взаимодействий “погодные условия – физиологический отклик на провокационное облучение” в фоновых выборках эффекты были положительные, а в импактных – отрицательные.
Кроме того, обнаружены временные лаги (k = 1) у импактных популяций по показателю сумма осадков за апрель (+) текущего сезона и октябрь (–) предыдущего сезона, т. е. аналогичные эффекты в фоновых выборках были отмечены на месяц позже – в мае (+) и ноябре (+) соответственно. По первому лагу значение коэффициента b0 для импактных выборок было сопоставимо с фоновыми площадками, а коэффициента b1 в 1.6–2.7 раз превышало его. По второму лагу эффект влияния осадков предыдущего сезона для пустырника из зоны ВУРСа был многократно сильнее (b0 – в 10.6 раза, а b1 – в 3.9 раза), чем за пределами зоны аварии.
Мутабильность семенного потомства пустырника
Мы рассмотрели влияние погодных условий на образование аномалий в трех группах признаков: 1) изменение формы разных органов (семядоли, листья, корни, гипокотиль, множественные изменения); 2) образование некрозов; 3) изменение цвета разных органов. В фоновых выборках разнообразие аномалий у проростков было почти в 2 раза меньше, чем в импактных, и все они положительно связаны с количеством осадков или производными этого параметра в разные месяцы (табл. 5). Так, обнаружены позитивные корреляции физиологического отклика с суммой осадков при эффективных температурах за отдельные летние месяцы и целый вегетационный сезон, суммой эффективных температур за вегетационный сезон, индексами Селянинова за апрель, июль и вегетационный сезон в целом. Значимо было и влияние отношения сумм осадков при эффективных температурах (осень прошлого–лето нынешнего года) к суммам эффективных температур за текущий вегетационный сезон. Число аномалий в развитии проростков пустырника также увеличивалось при избытке осадков в январе и марте текущего года и ноябре прошлого года.
Таблица 5.
Значения коэффициентов корреляции и анализ уравнений линейной регрессии y = b0 ± b1x для показателей мутабильности семенного потомства пустырника из разных зон
| Показатель | Климатический фактор | R | p-значение | b0 ± SE | b1 ± SE |
|---|---|---|---|---|---|
| Фоновые выборки | |||||
| Форма семядоли | Sum_P-Tef._5-8 | 0.68 | 0.0040 | –3.23± 3.64 | 0.02± 0.006 |
| Selyaninov_5-8 | 0.67 | 0.0040 | –4.90± 4.10 | 4.39± 1.28 | |
| P_5-8/T_5-8 | 0.63 | 0.0094 | –5.91± 4.93 | 42.4± 14.1 | |
| Pef._10-8/Tef._5-8 | 0.68 | 0.0040 | –4.96± 4.10 | 43.9± 3.43 | |
| Pef._6-8/Tef._6-8 | 0.67 | 0.0043 | –3.89± 3.84 | 35.9± 10.6 | |
| Корень скручен | Sum_P-Tef._6-8 | 0.63 | 0.0085 | –0.81± 0.60 | 0.003± 0.001 |
| Pef._10-8/Tef._5-6 | 0.63 | 0.0085 | –1.40± 0.78 | 3.20± 1.04 | |
| Pef._6-8/Tef._6-7 | 0.63 | 0.0091 | –1.07± 0.68 | 3.87± 1.28 | |
| Некроз семядоли | Sum_P-11 | 0.63 | 0.008 | –0.22± 3.23 | 0.08± 0.03 |
| Некроз листа | Sum_P-1 | 0.65 | 0.0064 | –0.011± 0.89 | 0.009± 0.003 |
| Sum_P-3 | 0.63 | 0.0087 | –0.011± 0.95 | 0.007± 0.002 | |
| Sum_P-Tef.-7 | 0.65 | 0.0064 | –1.44± 1.20 | 0.01± 0.003 | |
| Selyaninov-7 | 0.64 | 0.0078 | –1.72± 1.30 | 0.69± 0.22 | |
| Sum_Tef_5-8 | 0.65 | 0.0064 | –33.2± 10.87 | 0.02± 0.006 | |
| P_10-7/T_5-7 | 0.62 | 0.0097 | –2.44± 1.55 | 4.24± 1.42 | |
| Pef._10-7/Tef._6-7 | 0.63 | 0.0095 | –3.31± 1.80 | 11.9± 3.97 | |
| Pef._6-7/Tef._6-8 | 0.62 | 0.0099 | –2.90± 1.69 | 17.69± 5.93 | |
| Цвет семядоли | Selyaninov-4 | 0.76 | 0.0017 | 0.76± 1.31 | 5.68± 1.41 |
| Sum_P-11 | 0.79 | 0.0002 | 0.33± 1.11 | 0.04± 0.009 | |
| Импактные выборки | |||||
| Множественные изменения всех органов | Selyaninov-4 | 0.98 | 0.0005 | –0.03± 0.01 | 0.53± 0.05 |
| Sum_P-Tef.-10 | 0.92 | 0.009 | –0.04± 0.02 | 0.02± 0.005 | |
| Selyaninov-10 | 0.99 | 0.0001 | –0.02± 0.007 | 0.20± 0.014 | |
| Форма семядоли | Sum_P-5 | 0.69 | 0.0041 | –7.02± 4.56 | 0.28± 0.08 |
| Selyaninov_9 | –0.81 | 0.008 | 56.2± 14.0 | –47.2± 12.7 | |
| Форма листа | Sum_P-5 | 0.74 | 0.0016 | –0.68± 0.29 | 0.02± 0.005 |
| Selyaninov-5 | 0.69 | 0.0047 | –0.19± 0.21 | 0.52± 0.15 | |
| Гипокотиль скручен | Т_mean-5 | –0.71 | 0.0031 | 41.3± 9.81 | –0.10± 0.03 |
| Sum_P-3 | –0.67 | 0.0065 | 18.9± 4.21 | –0.43± 0.13 | |
| Корень скручен | Т_mean-10 | 0.83 | 0.0001 | 0.017± 0.09 | 0.075± 0.014 |
| Sum_P-5 | 0.69 | 0.0042 | –0.23± 0.19 | 0.01± 0.003 | |
| Sum_P-9 | –0.76 | 0.001 | 0.20± 0.09 | –0.001± 0.0003 | |
| Sum_P-11 | –0.67 | 0.007 | 0.93± 0.19 | –0.015± 0.005 | |
| Sum_Tef.-4 | 0.87 | 0.0002 | –0.078± 0.09 | 0.01± 0.002 | |
| Sum_Tef.-9 | –0.71 | 0.003 | 0.89± 0.16 | –0.004± 0.001 | |
| Sum_Tef.-7 | 0.72 | 0.0024 | –1.03± 0.39 | 0.002± 0.0008 | |
| Selyaninov-7 | –0.67 | 0.0059 | 0.87± 0.17 | –0.36± 0.11 | |
| Некроз листа | Pef._10-6/Tef._5-8 | –0.65 | 0.0094 | 3.39± 0.82 | –43.3± 14.2 |
| Некроз корня | Sum_P-10 | –0.65 | 0.009 | 4.76± 1.26 | –0.09± 0.03 |
| Цвет семядоли | Т_mean-9 | 0.65 | 0.009 | –26.0± 9.5 | 2.9± 0.93 |
| Цвет шейки корня | Т_mean-5 | –0.65 | 0.0089 | 23.2± 6.76 | –1.83± 0.59 |
| Sum_Tef.-6 | –0.67 | 0.0068 | 34.2± 9.83 | –0.07± 0.021 | |
| Т_mean-11 | –0.81 | 0.0003 | 1.57± 0.78 | –0.021± 0.004 | |
| Т_mean-12 | –0.73 | 0.002 | –3.00± 1.72 | –0.02± 0.005 | |
В импактных выборках отмечены аномалии, не встречавшиеся в фоновых (множественные изменения всех органов, нарушение формы листа и гипокотиля, некроз корней и изменение цвета корневой шейки). Выявлена взаимосвязь появления морфозов с разными погодными факторами, причем влияние было как положительным, так и отрицательным. Для первой группы аномалий (форма) были важны метеорологические условия за сентябрь (T–, P–, Selyaninov–) и октябрь (Tср+, P+, Selyaninov+) прошлого года, за апрель (Tэф+, Selyaninov+), май (T–, P +, Selyaninov+) и июль (Tэф+, Selyaninov–) текущего года. Кроме того, проростки со скрученными корнями формировались чаще при низких осадках за ноябрь прошлого года. Для второй группы морфозов (некрозы) были значимы метеоусловия за октябрь прошлого года (P–), а также соотношение “сумма осадков при эффективных температурах текущего и прошлого года/сумма эффективных температур за весенне-летний сезон текущего года” (–). Для третьей группы аномалий (цвет) были важны эффективные температуры за июнь текущего года (+), а также средние температуры за ноябрь и декабрь прошлого года (–).
Таким образом, для фоновых и импактных выборок обнаружено 5 общих метеорологических факторов (Sum_P-3, Selyaninov-4, Selyaninov-7, Sum_P-11 и отношение сумм осадков при эффективных температурах (осень прошлого–лето нынешнего года) к суммам эффективных температур за текущий вегетационный сезон), значимо влияющих на мутабильность семенного потомства пустырника. При этом только индекс Селянинова за апрель действовал одинаково положительно на формирование аномалий в развитии у проростков в обеих зонах. Другие указанные выше погодные условия в фоновых популяциях давали положительные эффекты, а в импактных – отрицательные (см. табл. 5). Значение коэффициента b0 для схожих, но не идентичных факторов (Pэф-10-7/Tэф.-6-7; Pэф.-10-6/Tэф.-5-8) в импактных выборках (3.39) было численно сопоставимо с противоположным по знаку b0 в фоновых (–3.31), но коэффициент b1 импактных выборок (–43.3) превышал численно b1 фоновых (11.9) в 3.6 раза, что свидетельствует о большем влиянии данных факторов и противоположной направленности их действия в условиях хронического облучения материнских растений.
ОБСУЖДЕНИЕ
Анализ влияния погодных условий текущего года на межгодовую изменчивость качества семенного потомства пустырника пятилопастного показал, что самым значимым метеорологическим фактором для всех исследованных физиологических показателей является индекс Селянинова за апрель. При этом для фоновых и импактных выборок однонаправленные изменения отмечены по жизнеспособности (–) и мутабильности семенного потомства (+). Следовательно, семенное потомство пустырника с высокими показателями жизнеспособности и низкой долей аномальных проростков формируется при дефиците осадков и повышенной температуре в начале вегетационного сезона независимо от уровня дозовых нагрузок материнских растений. Отклик на такие условия радиоустойчивости семенного потомства в фоновых выборках был позитивный.
Эти результаты хорошо согласуются с исследованием радиоустойчивости семян в условиях повышенной температуры и водного дефицита [30, 31]. В.В. Стогний [31] связывает подобные эффекты с неспецифической реакцией образования активных форм кислорода в засушливых условиях: для снижения их концентрации происходит активация системы антиоксидантной защиты, прежде всего низкомолекулярных антиоксидантов, обеспечивающих высокую устойчивость семян к облучению. Высокая радиоустойчивость может быть связана также с повышенной активностью ферментов (супероксидпероксидаза, каталаза и дисмутаза) и таких неферментных антиоксидантов, как аскорбиновая кислота и глутатион, с повышением уровня метаболитов (пролина, общего растворимого сахара и белка) [32, 33]. Кроме того, семена, сформировавшиеся в условиях водного дефицита, дольше сохраняли жизнеспособность при хранении, соответственно осадки в самые засушливые месяцы репродуктивного периода были самым сильным предиктором изменчивости [34].
Для импактных популяций была характерна иная реакция: при дефиците осадков отмечена низкая радиоустойчивость. Эти данные согласуются с исследованиями, проведенными на разных по радиоустойчивости сортах яровой пшеницы: повреждающий эффект радиации усиливался в засушливый период вегетации [29]. Прохладные и избыточно увлажненные условия формирования семян приводили к снижению жизнеспособности семенного потомства Triticum aestivum, однако после дополнительного облучения была отмечена стимуляция роста и развития проростков. Кроме того, такие условия индуцировали увеличение доли аномальных проростков относительно оптимальных погодных условий [35].
Отметим, что для популяций из зоны ВУРСа, кроме индекса Селянинова за апрель, были важны эффективные температуры за апрель, которые оказывали положительное влияние на все исследованные характеристики (жизнеспособность, мутабильность, радиочувствительность) семенного потомства пустырника пятилопастного. Можно было бы предположить, что небольшое число значимых эффектов между погодными условиями и физиологическим откликом в фоновых популяциях связано с большой вариабельностью качества семенного потомства пустырника (по выживаемости CV = 35.4%). Однако межгодовая изменчивость выживаемости проростков в зоне ВУРСа была не меньше (CV = 32.4%). Вероятно, в условиях хронического облучения выборки становятся более чувствительными и лабильными к любым изменениям окружающей среды, а в результате взаимодействия техногенного фактора и погодных условий возникают синергические эффекты [36].
Нами установлено, что сумма осадков за ноябрь прошлого года (Sum_P-11) оказалась значимой для критериев оценки мутабильности и радиоустойчивости семенного потомства пустырника из фоновых (+) и импактных (–) выборок, за исключением показателей жизнеспособности семенного потомства из зоны ВУРСа, где эффекты отсутствовали. Значимое воздействие погодных условий в эти месяцы на качество семян следующего сезона выявлялось и ранее. Так, была показана отрицательная связь между осадками сентября–октября прошлого года и числом цветоносов у одуванчика [37]; также отмечена отрицательная связь между фитомассой и осенними осадками для фоновых популяций и положительная – для импактных [38]. Однако в нашем случае осадки за ноябрь прошлого года положительно влияли на качество семенного потомства пустырника из фоновых выборок и отрицательно – из импактных. Аналогичные данные были получены при анализе отношения сумм осадков (осень прошлого–лето нынешнего года) к суммам температур за текущий вегетационный сезон. Осенние периоды с небольшим количеством осадков и высокими эффективными температурами являются благоприятными для формирования надземной и подземной фитомассы [38]. В целом высокие температуры и водный стресс в период созревания увеличивали всхожесть семян Arabidopsis thaliana [39].
В работе [40], посвященной оценке независимого и совместного действия метеофакторов и ионизирующего излучения на обилие гнездящихся птиц на Фукусиме, показано, что воздействие ионизирующего излучения было сильнее, чем осадков и температуры. Вероятно, это связано с тем, что исследован первый период после аварии, когда мощности доз велики и действие излучений проявляется наиболее сильно. В то же время исследования отдаленных последствий хронического облучения в зоне ВУРСа [3, 11–13, 16], в зоне ЧАЭС [14, 15, 41], а также приведенные в данной статье результаты свидетельствуют о том, что погодные факторы играют ключевую роль при формировании качества семенного потомства растений. В работе [15] показано отсутствие корреляция между уровнем радиационной нагрузки и долей абортивных семян, а также всхожестью семян сосны, что хорошо согласуется с нашими результатами. В этой же работе установлено, что высокая эффективная температура и низкое количество осадков в мае снижали долю абортивных семян у сосны в зоне ЧАЭС. В то же время высокие эффективные температуры в августе [15] и осадки в мае [14] приводили к снижению всхожести семян сосны. Противоположные зависимости были получены при исследовании некоторых видов травянистых растений ВУРСа: у Taraxacum officinale, Melandrium alba отмечены положительные зависимости между жизнеспособностью семенного потомства и температурой мая, июня, августа [3, 12]; у Stellaria graminea отмечены негативные зависимости качества семян от температуры в июне [16]. Исходя из сказанного выше, мы полагаем, что приоритетность значимости факторов (облучение или погодные условия), действующих на природные популяции, определяется периодом времени, прошедшим после аварии, а также видовыми особенностями исследованных объектов.
ЗАКЛЮЧЕНИЕ
Из 184 проанализированных метеорологических факторов и их производных выделены два основных – индекс Селянинова за апрель и сумма осадков за ноябрь, значимых для основных характеристик качества семенного потомства пустырника как из зоны радиоактивного загрязнения, так и фоновых территорий. Независимо от уровня дозовых нагрузок материнских растений семенное потомство с высокими показателями жизнеспособности и низкой долей аномальных проростков формировалось при дефиците осадков и повышенной температуре в начале вегетационного сезона и достаточном количестве осадков поздней осенью предыдущего года. Отклик на такие условия радиоустойчивости семенного потомства в фоновых выборках был позитивный, а в импактных – негативный. Эффект влияния погодных условий на качество семенного потомства пустырника (жизнеспособность, радиоустойчивость, мутабильность), сформировавшихся в зоне ВУРСа, проявлялся, как правило, сильнее, чем в фоновых выборках. Полученные данные о влиянии погодных условий на качество семенного потомства растений, произрастающих в условиях хронического облучения, указывают на необходимость учитывать унифицированный комплекс метеорологических параметров при прогнозировании последствий радиоактивного загрязнения экосистем.
Мы благодарим к.б.н. Э.М. Каримуллину, н.с. В.П. Гусеву, инженера Т.Е. Беляеву, м.н.с. Н.С. Шималину (ИЭРиЖ УрО РАН), бакалавров Д.Ю. Росяеву, А.С. Чиркову, Т.В. Ласкину (УрФУ) за помощь в сборе материала, подготовке и проведении экспериментов, создании электронных баз данных. Работа выполнена в рамках государственного задания Института экологии растений и животных УрО РАН (АААА-А19-119032090023-0).
Список литературы
Sparrow A.H., Schwemmer S.S., Bottino P.J. The effects of external gamma radiation from radioactive fallout on plants with special reference to crop production // Radiation Botany. 1971. V. 11. № 2. P. 85‒118.
Petin V.G., Zhurakovskaya G.P., Komarova L.N., Ryabova S.V. Synergism of environmental factors as a function of their intensity // Rus. J. of Ecology. 1998. V. 29. № 5. P. 338‒343.
Pozolotina V.N., Antonova E.V. Temporal variability of the quality of Taraxacum officinale seed progeny from the East-Ural Radioactive Trace: is there an interaction between low level radiation and weather conditions? // Intern. J. of Radiation Biology. 2017. V. 93. № 3. P. 330‒339.
Environmental protection – the concept and use of reference animals and plants. ICRP Publication 108. International Commission on Radiological Protection. 2008.
Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В. Очерк учения о популяции. М.: Наука, 1973. 277 с.
Bréchignac F., Oughton D., Mays P. et al. Addressing ecological effects of radiation on populations and ecosystems to improve protection of the environment against radiation: Agreed statements from a Consensus Symposium // J. of Environmental Radioactivity. 2016. V. 158‒159. P. 21‒29.
Alonzo F., Hertel-Aas T., Real A. et al. Population modelling to compare chronic external radiotoxicity between individual and population endpoints in four taxonomic groups // J. of Environmental Radioactivity. 2016. V. 152. P. 46‒59.
Garnier-Laplace J., Della-Vedova P., Andersson P. et al. A multi-criteria weight of evidence approach for deriving ecological benchmarks for radioactive substances // J. of Radiological Protection. 2010. V. 30. № 2. P. 215‒233.
Andersson P., Garnier-Laplace J., Beresford N.A. et al. Protection of the environment from ionising radiation in a regulatory context (protect): proposed numerical benchmark values // J. of Environmental Radioactivity. 2009. V. 100. P. 1100–1108.
Bradshaw P., Kapustka L., Barnthouse L. et al. Using an Ecosystem Approach to complement protection schemes based on organism-level endpoints // J. of Environmental Radioactivity. 2014. V. 136. P. 98‒104.
Pozolotina V.N., Antonova E.V., Karimullina E.M. Assessment of radiation impact on Stellaria graminea cenopopulations in the zone of the Eastern Ural Radioactive Trace // Rus. J. of Ecology. 2010. V. 41. № 6. P. 459−468.
Antonova E.V., Karimullina E.M., Pozolotina V.N. Intraspecific variation in Melandrium album along a radioactive contamination gradient at the Eastern Ural radioactive trace // Rus. J. of Ecology. 2013. V. 44. № 1. P. 18‒27.
Antonova E.V., Pozolotina V.N., Karimullina E.M. Variation in the seed progeny of smooth brome grass, Bromus inermis Leyss., under conditions of chronic irradiation in the zone of the Eastern Ural Radioactive Trace // Rus. J. of Ecology. 2014. V. 45. № 6. P. 508−516.
Geras’kin S.A., Vasiliev D.V., Kuzmenkov A.G. Specific Features of Scots Pine Seeds Formation in the Remote Period after the Chernobyl NPP Accident // Radiats Biol Radioecol. 2015. V. 55. № 5. P. 539−547.
Geras’kin S., Vasiliyev D., Makarenko E. et al. Influence of long-term chronic exposure and weather conditions on Scots pine populations // Environmental Science and Pollution Research. 2017. V. 24. № 12. P. 11 240‒11 253.
Antonova E.V., Pozolotina V.N., Karimullina E.M. Viability of plant seed progeny from the East-Ural Radioactive Trace: radiation and weather condition // Genetics, Evolution and Radiation: Crossing Borders, The Interdisciplinary Legacy of Nikolay W. Timofeeff-Ressovsky / Eds Korogodina V.L. et al. Springer, 2016. P. 267‒276.
Экологические последствия радиоактивного загрязнения на Южном Урале / Под ред. Соколова В.Е., Криволуцкого Д.А. М.: Наука, 1993. 336 с.
Pozolotina V.N., Molchanova I.V., Mikhaylovskaya L.N. et al. The current state of terrestrial ecosystems in the Eastern Ural Radioactive Trace // Radionuclides: Sources, Properties and Hazards / Ed. Gerada J.G. New York: Nova Science, 2012. P. 1−22.
Karimullina E.M., Mikhailovskaya L.N., Pozolotina V.N., Antonova E.V. Radionuclide uptake and dose assessment of 14 herbaceous species from the East-Ural Radioactive Trace area using the ERICA Tool // Environmental Science and Pollution Research. 2018. V. 25. № 14. P. 13 975–13 987.
Позолотина В.Н., Молчанова И.В., Караваева Е.Н. и др. Современное состояние наземных экосистем зоны Восточно-Уральского радиоактивного следа. Екатеринбург: Изд-во “Гощицкий”, 2008. 204 с.
Molchanova I., Mikhailovskaya L., Antonov K. et al. Current assessment of integrated content of long-lived radionuclides in soils of the head part of the East Ural Radioactive Trace // J. Environmental Radioactivity, 2014. V. 138 № 6. P. 238−248.
Чибилёв А.А., Чибилёв А.А. Природное районирование Урала с учётом широтной зональности, высотной поясности и вертикальной дифференциации ландшафтов // Изв. Самарского научного центра РАН. 2012. V. 14 № 1−6. P. 1660−1665.
Флора и растительность биологической станции Уральского государственного университета / Мухин В.А., Третьякова А.P., Тептина А.Ю. и др. Екатеринбург: Изд-во Урал. ун-та, 2003. 132 с.
Кайгородова С.Ю. Трансформация морфологии почв в зоне воздействия Карабашского медеплавильного завода // Изв. Оренбургского гос. аграрного ун-та. 2012. Т. 38. № 6. С. 13−17.
Иллюстрированный определитель растений Cредней России. Покрытосеменные (двудольные: раздельнолепестные) / Губанов И., Киселева К., Новиков В., Тихомиров В. М.: Тов-во научн. изд. КМК, Ин-т технологических исследований, 2004. 586 с.
Oliver E.J. The Encyclopedia of World Climatology (Encyclopedia of Earth Sciences Series). Dordrecht: Springer, 2005. 854 p.
Newcombe R.G. Interval estimation for the difference between independent proportions: Comparison of eleven methods // Statistics in Medicine. 1998. V. 17. № 8. P. 873−890.
Ульяненко Л.Н., Удалова А.А. Оценка состояния окружающей среды по реакции сельскохозяйственных растений на действие ионизирующих излучений // Радиация и риск. Бюл. Национального радиационно-эпидемиологического регистра. 2015. Т. 24. № 1.
Ульяненко Л.Н., Круглов P.В., Филипас А.P., Алексахин Р.М. Влияние ионизирующего излучения и климатических факторов на продуктивность пшеницы // Сельскохозяйственная биология. 2001. № 5. P. 69−74.
Янушкевич С.И. Влияние условий выращивания ячменя и пшеницы на эффект гамма-облучения: Автореф. дис. … канд. биол. наук. Кишинев: МГУ, 1964.
Стогний В.В. Влияние климато-географических условий на радиоустойчивость и антиоксидантный статус семян дикорастущих травянистых растений: Автореф. дис. … канд. биол. наук. Якутск, 2001.
Khan M.N., Zhang J., Luo T. et al. Morpho-physiological and biochemical responses of tolerant and sensitive rapeseed cultivars to drought stress during early seedling growth stage // Acta Physiologiae Plantarum. 2019. V. 41. № 2. Article 25.
Avramova V., Abd Elgawad H., Zhang Z. et al. Drought induces distinct growth response, protection, and recovery mechanisms in the maize leaf growth zone // Plant Physiology. 2015. V. 169. № 2. P. 1382−1396.
Zani D., Müller J.V. Climatic control of seed longevity of Silene during the post-zygotic phase: do seeds from warm, dry climates possess higher maturity and desiccation tolerance than seeds from cold, wet climates? // Ecological Research. 2017. V. 32. № 6. P. 983−994.
Antonova E.V., Khlestkina E.K. Radiosensitivity and mutability of wheat seed progeny cultivated under adverse environments // Plant Physiology and Biochemistry. 2019. V. 137. P. 162−168.
Bezel’ V.S., Pozolotina V.N., Bel’skii E.A., Zhuikova T.V. Variation in population parameters: Adaptation to toxic environmental factors // Rus. J. of Ecology. 2001. V. 32. № 6. P. 413−419.
Жуйкова Т.В. Реакция ценопопуляций и травянистых сообществ на химическое загрязнение среды: Автореф. дис. … докт. биол. наук. Екатеринбург, 2009.
Zhuikova T.V., Bezel’ V.S., Bergman I.E. et al. Dependence of hytomass of herbaceous cenoses on weather factors in anthropogenically impacted areas // Contemporary Problems of Ecology. 2018. V. 11. № 4. P. 428−437.
Abo Gamar M.I., Qaderi M.M. Interactive effects of temperature, carbon dioxide and watering regime on seed germinability of two genotypes of Arabidopsis thaliana // Seed Science Research. 2019. V. 29. № 1. P. 12−20.
Møller A.P., Mousseau T.A. Interactive effects of ionizing radiation and climate change on the abundance of breeding birds // Ecological Indicators. 2019. V. 99. P. 178−182.
Гераськин С.А., Кузьменков А.Г., Васильев Д.В. Временнáя динамика цитогенетических эффектов в хронически облучаемых популяциях сосны обыкновенной // Радиационная биология. Радиоэкология. 2018. Т. 58. № 1. P. 74−84.
Дополнительные материалы отсутствуют.