

Экология, 2020, № 5, стр. 397-400
Опыт изучения избирательности питания Vespertilio murinus (Chiroptera, Vespertilionidae)
Е. М. Первушина a, *, О. Р. Гизуллина a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: pervushina@ipae.uran.ru
Поступила в редакцию 16.10.2019
После доработки 09.12.2019
Принята к публикации 18.12.2019
Согласно теории оптимальной фуражировки, для оптимизации соотношения затрат и выгод у особей в популяции формируются определенные стратегии пищедобывательного поведения и таким образом проявляется ответ популяции на изменения условий среды [1]. Одним из этапов пищедобывательного поведения является выбор хищником добычи. Как известно, этот выбор определяется целым рядом факторов, в том числе доступностью подходящих пищевых объектов в окружающей среде [2]. Проведение подобных исследований представляет интерес. В то же время влияние нестабильных условий среды и различных особенностей поведения самих животных усложняет изучение избирательности питания в природе и представляет собой технически сложную задачу, которая может быть выполнима на небольших выборках.
Ранее на территории России не исследовали избирательность питания у летучих мышей. Отечественные работы в области трофической биологии летучих мышей обычно сводятся к рассмотрению структуры рациона и сравнительному анализу рационов разных видов [3, 4 и др.]. За рубежом они более актуальны и представляют собой анализ рациона видов наряду с оценкой кормовых ресурсов [5–7 и др.].
Цель нашей работы – изучить избирательность питания летучих мышей на примере двухцветного кожана Vespertilio murinus Linnaeus, 1758 путем сравнения спектра питания вида и обилия его потенциальных жертв – насекомых – в окружающей среде. Интересно было выяснить, выбирают ли животные отдельные группы насекомых из массы активных ночью членистоногих или же поедают многочисленные и доступные корма.
Исследования проводили в окрестностях Научно-практического центра биоразнообразия, с. Скородумское Ирбитского р-на Свердловской обл. (57.34° с.ш., 62.43° в.д.). Территория расположена в подзоне южной тайги равнинного Среднего Зауралья [8]. Объект исследования – обычный в Среднем Зауралье перелетный вид – охотник открытых пространств, который использует кормовые стации в различных биотопах.
Материал собирали в период репродуктивной активности летучих мышей в середине июля. Ранее было показано [8], что таксономическое разнообразие и численность насекомых – потенциальных жертв летучих мышей – положительно коррелирует с температурой окружающей среды. Чтобы избежать влияния большого количества факторов, материал собирали через относительно короткий временной промежуток в разные по погодным условиям даты – 11 и 16 июля 2012 г. (табл. 1). Животных отлавливали около убежища по окончании ночного кормления (с 3:00 до 5:00 ч). Через 12 ч после поимки собирали экскременты на анализ рациона для каждой особи, затем их выпускали в природу. В даты сбора во время активности летучих мышей (с 23:00 до 5:00 ч) отлавливали насекомых с помощью световой ловушки, установленной около убежища – центра охотничьей территории колонии вида в несколько километров. Использовали ловушку механического типа с ртутной лампой Philips ML (250 Вт). Данный метод работает на открытых пространствах и позволяет учесть большинство потенциальных для питания двухцветного кожана групп насекомых, объективно показывая изменение их активности [9].
Таблица 1.
Данные отлова светоловушкой и анализа экскрементов летучих мышей
| Название таксона | 11 июля 2012 г. | 16 июля 2012 г. | ||||
|---|---|---|---|---|---|---|
| Т = 14.6–19.6°С, влажность 46–65% | Т = 22.1–26°С, влажность 69–80% | |||||
| свето-ловушка, число особей | экскременты, обилие (M ± m) |
экскременты, встречаемость (M ± m) |
свето-ловушка, число особей |
экскременты, обилие (M ± m) |
экскременты, встречаемость (M ± m) |
|
| Diptera (Nematocera) | 1534 | 28.5 ± 1.3 | 100 ± 0 | 3760 | 10.8 ± 1.6 | 95 ± 0 |
| Diptera (Brachycera) | + | 0.9 ± 0.4 | 22.8 ± 6.8 | 95 | 0.1 ± 0.02 | 5.0 ± 0 |
| Coleoptera | 42 | 11.9 ± 1.3 | 94.2 ± 7.5 | 1823 | 16.7 ± 1.1 | 97.5 ± 1.7 |
| Lepidoptera | 351 | 2.1 ± 0.6 | 48.5 ± 10.5 | 362 | 2.4 ± 0.3 | 40 ± 0 |
| Homoptera (Cicadellidae) | 4 | 3.9 ± 0.5 | 65.7 ± 10.1 | 620 | 16.3 ± 0.1 | 97.5 ± 2.5 |
| Homoptera (Aphididae) | – | 10.3 ± 1.8 | 81.4 ± 7.05 | 3 | 1.4 ± 0.6 | 20 ± 5.0 |
| Hemiptera | 4 | 4.1 ± 1.5 | 57.1 ± 10.1 | 1820 | 4.03 ± 1.2 | 55 ± 5.0 |
| Triсhoptera | 233 | 1.08 ± 0.4 | 22.8 ± 8.08 | 728 | 1.9 ± 0.3 | 37.5 ± 2.5 |
| Neuroptera (Hemerobiidae) | – | 5.4 ± 1.2 | 60 ± 13.3 | 1 | 4.1 ± 2.1 | 55 ± 20.0 |
| Neuroptera (Chrysopidae) | – | 0.4 ± 0.4 | 10 ± 8.4 | 4 | 2.2 ± 0.3 | 32.5 ± 12.5 |
| Ephemeroptera | – | – | – | 29 | – | – |
| Hymenoptera (Ichneumonidae, Braconidae) | 24 | 2.06 ± 0.5 | 42.8 ± 10.1 | 324 | 0.21 ± 0.09 | 7.5 ± 2.5 |
| Acari | 19 | 0.2 ± 0.1 | 14.2 ± 5.7 | 74 | – | – |
| Araneae | 1 | – | – | – | – | – |
| Неопределенные остатки | – | 28.5 ± 1.3 | 100 ± 0 | – | 39.5 ± 3.3 | 100 ± 0 |
| Сумма | 2212 | 9643 | ||||
Температуру воздуха и влажность измеряли с помощью электронного термогигрометра RST (Швеция).
Экскременты анализировали с помощью визуального изучения остатков насекомых. Специфика определения подробно описана нами ранее [10]. Для каждой особи случайно отбирали 5–20 экскрементов близкого размера и формы (проб). Остатки членистоногих в каждой пробе равномерно распределяли по поверхности предметного стекла (площадь 14 см2), под которое подкладывали миллиметровую бумагу, размеченную на квадраты размером 1 см2 (всего 14 квадратов в пробе). Анализ проводили с помощью стереомикроскопа Olympus SZ61 в диапазоне увеличений ×40–80.
Насекомые в пробах были представлены многочисленными мелкими фрагментами (размером 0.1 мм). Поэтому численность остатков учитывали с помощью относительной встречаемости – доли проб, в которых обнаружены остатки представителей таксона, от общего числа проб (для каждой особи) [3, 11], где одна проба – экскремент. Для оценки соотношения разных таксонов рассчитывали относительное обилие – долю квадратов (1 см2), в которых отмечен данный таксон, от суммы квадратов для всех найденных таксонов (для каждой особи) [11]. С помощью среднего арифметического и его ошибки (см. табл. 1) определяли через критерий Стьюдента значимость результатов – насколько выборочные данные характеризуют рацион вида. Подсчет квадратов является более объективной оценкой обилия, чем обычно используемая система баллов [12]. Но остается не решенной проблема учета неопределенных остатков и фрагментов одного насекомого, возможно, распределяемого на несколько проб.
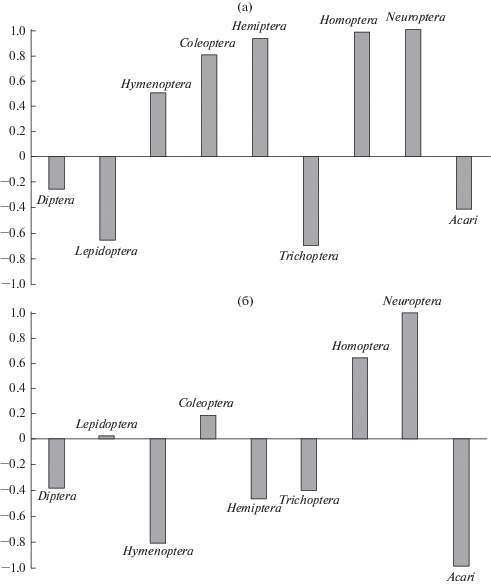
Для количественной оценки избирательности питания животных использовали индекс “элективности” Ивлева [13]:
где E – значение индекса; ri – усредненное относительное обилие i-того таксона среди всех таксонов насекомых, найденных в экскрементах; pi – доля того же таксона в сборах светоловушки (рис. 1). Индекс Ивлева принимает значения от –1 (абсолютное избегание) до +1 (абсолютное предпочтение). Значения индекса, близкие к 0, означают, что не проявляется ни избегание, ни предпочтение. Дополнительно к индексу Ивлева мы оценили связь показателей с помощью коэффициена корреляции Спирмена. Значимость различий рационов вида между датами оценивали с помощью непараметрических критериев Краскела-Уоллиса и Манна-Уитни. Для расчетов использовали число квадратов на предметном стекле, занятых фрагментами таксона. В ходе анализа мы исключили неопределенные остатки, а также Acari, которые, вероятно, попадают в экскременты с остатками насекомых-хозяев.Рис. 1.
Значения индекса Ивлева для 11.07.2012 (а) и 16.07.2012 (б): а – Т = 14.6–19.6°С, влажность 46–65%; б – Т = 22.1–26°С, влажность 69–80%.
Всего было отловлено светоловушкой около 12 000 членистоногих, изучено 80 проб экскрементов: 40 проб (7 особей) – 11.07.2012 и 40 проб (2 особи) – 16.07.2012. Математическую обработку данных проводили с использованием пакета программ Statistica for Windows 6.0 и Excel.
Результаты исследований показали, что изучаемые даты различаются по обилию насекомых (см. табл. 1). Показатели температуры и влажности 11 июля были несколько ниже, чем 16 июля, и светоловушкой было отловлено небольшое количество членистоногих (2122 особи, 10 групп). Наоборот, при более комфортных погодных условиях 16 июля (температура 26–22.1°C, влажность 69–80%) было больше групп (14) и количество членистоногих (9653 особи), что свидетельствует об оптимальности погодных условий для их активности во вторую дату.
В питании двухцветного кожана отмечены те же группы насекомых, что и в светоловушке, за исключением Ephemeroptera (см. табл. 1). Все они являются фотоксенами, т.е. привлекаются источниками искусственного света [9]. Рацион вида в разные дни был схож по составу членистоногих, однако частоты обилия и встречаемости различались. Так, заметно снижение средних значений обилия за вторую, более “теплую дату” для Diptera (Nematocera), Hymenoptera, Homoptera (Aphididae) и увеличение частот обилия для Coleoptera и Homoptera (Cicadellidae). Показатели встречаемости за вторую дату уменьшились для Diptera (Brachycera), Homoptera (Aphididae), Hymenoptera и увеличились для Neuroptera (Chrysopidae), Homoptera (Cicadellidae). Следует отметить, что Neuroptera (Hemerobiidae) стабильно присутствовали в рационе всех особей, и это свидетельствует о важности данного ресурса в питании вида. Похожие данные получены польскими исследователями [12].
С учетом ошибки среднего (см. табл. 1) за “холодную дату” значимые результаты о рационе вида получены по всем кормовым группам (p < 0.05), а за “теплую дату” ввиду малого числа особей – только для Homoptera (Cicadellidae) и Coleoptera. Поэтому сравнение рациона двухцветного кожана за разные даты мы проводили только по этим двум группам насекомых, значимые различия (p < 0.05) характерны для Homoptera (Cicadellidae).
Изначально мы предполагали, что летучие мыши поедают многочисленные и доступные группы насекомых в пределах охотничьего пространства в конкретный период времени. Однако среди обильных в отловах насекомых значение в питании вида за обе даты имели только Diptera. Для этой группы индекс Ивлева демонстрирует отрицательные значения (см. рис. 1), так как доля данного корма в среде существенно превышает его долю в питании. По сравнению с ними другие членистоногие со стабильно высокой численностью (Triсhoptera и Lepidoptera) как пищевые объекты незначительны. Похожие результаты по Diptera получены для средиземноморского нетопыря Pipistrellus kuhlii (Kuhl 1817) [11].
Кроме того, высокий уровень предпочтения двухцветным кожаном имеют немногочисленные в отловах Homoptera и Neuroptera (см. рис. 1). Для представителей отрядов Homoptera (Cicadellidae) и Neuroptera (Chrysopidae) выявлена значимая положительная связь между обилием в рационе вида и обилием в светоловушке (коэффициент корреляции R = 0.28–0.6, p < 0.05). Это свидетельствует о том, что данные кормовые объекты имеют высокий уровень предпочтения при увеличении их обилия в окружающей среде. Сюда же можно отнести и Coleoptera. Для этой группы насекомых показатели встречаемости и обилия стабильно высокие, а индекс Ивлева за обе даты положительный. В целом наши результаты согласуются с литературными данными [6, 11]. Авторами одной из работ [6] высказывается предположение, что летучие мыши избирательно поедают перечисленные группы насекомых, так как в них содержатся микроэлементы (натрий, азот, кальций, железо и калий) [14], необходимые для роста и развития млекопитающих.
Таким образом, изобилие является важным фактором, определяющим выбор летучими мышами кормовых объектов. В то же время животные поедают не только многочисленные в данный момент времени ресурсы, а выбирают определенные группы насекомых, переключаются на более широкий спектр кормов, когда численность предпочитаемых объектов питания снижена. По степени избирательности мы выделили насекомых, которые обычно присутствуют в большом количестве как в питании двухцветного кожана, так и в среде обитания, – это Diptera. Кроме того, имеются насекомые, которых летучие мыши, вероятно, поедают селективно при резком росте их обилия в окружающей среде – Coleoptera, Homoptera (Cicadellidae) и, возможно, Neuroptera (Chrysopidae). При увеличении их доли в рационе вида заметно сокращается потребление Diptera. К насекомым, в отношении которых летучие мыши проявляют избирательность независимо от численности в светоловушке, мы отнесли Neuroptera (Hemerobiidae), которые стабильно присутствуют в питании вида в небольших количествах.
Благодарим за помощь в определении насекомых сотрудников ИЭРиЖ УрО РАН Г.А. Замшину, к.б.н. А.В. Иванова, к.б.н. Н.В. Николаеву, за техническую поддержку А.А. Первушина (Свердловский областной краеведческий музей). Сбор материала и техническое оснащение поддержаны РФФИ (проект № 12-04-31257), частично Программой фундаментальных исследований УрО РАН (№ 15-3-4-49). Интерпретация результатов выполнена в рамках государственного задания Института экологии растений и животных УрО РАН.
Список литературы
Резникова Ж.И. Между драконом и яростью: Этологические и эволюционные аспекты межвидовых отношений животных (гипотезы и теории; хищники и жертвы). Ч. II. М.: Научный мир, 2000. 208 с.
Мак-Фарленд Д. Поведение животных: Психобиология, этология и эволюция. Пер. с англ. М.: Мир, 1988. 520 с.
Смирнов Д.Г., Вехник В.П. Экология питания и дифференциация трофических ниш рукокрылых (Chiroptera: Vespertilionidae) в пойменных экосистемах Самарской Луки // Изв. РАН. Сер. биол. 2014. № 1. С. 53–64.
Казаков Д.В., Махов И.А. Питание сибирского ушана (Plecotus ognevi) в различного типа стациях в Байкальской Сибири // Изв. Иркутского гос. ун-та. Сер. “Биология. Экология”. 2015. Т. 11. С. 32–42.
Jones G. Prey selection by the greater horseshoe bat (Rhinolophus ferrumequinum): optimal foraging by echolocation? // J. of Animal Ecology. 1990. V. 59. P. 587–602.
Agosta S.J., Morton D., Kuhn K.M. Feeding ecology of the bat Eptesicus fuscus: ‘preferred’ prey abundance as one factor influencing prey selection and diet breadth // J. Zool., London. 2003. V. 260. P. 169–177.
Vesterinen E.J., Ruokolainen L., Wahlberg N. et al. What you need is what you eat? Prey selection by the bat Myotis daubentonii // Molecular Ecology. 2016. V. 25. P. 1581–1594.
Первушина Е.М., Замшина Г.А., Николаева Н.В. и др. Структура потенциального энтомокомплекса в питании летучих мышей равнинного Среднего Зауралья // Сибирский экологич. журн. 2015. № 2. С. 268–279.
Горностаев Г.Н. Введение в этологию насекомых-фотоксенов (лёт насекомых на искусственные источники света) // Этология насекомых. Труды ВЭО. Т. 66. Л.: Наука, 1984. С. 101–167.
Гизуллина О.Р., Первушина Е.М. Определение фрагментов насекомых в питании двухцветного кожана в Среднем Зауралье // Вестн. Томского гос. ун-та. Биология. 2016. № 3. С. 92–106.
Goiti U., Vecin P., Garin I. et al. Diet and prey selection in Kuhl’s pipistrelle Pipistrellus kuhlii (Chiroptera: Vespertilionidae) in south-western Europe // Acta Theriologica. 2003. V. 48. № 4. P. 457–468.
Bauerova Z., Ruprecht A. Contribution to the knowledge of the trophic ecology of the parti-coloured bat, Vespertilio murinus // Folia Zool. 1989. V. 38. № 2. P. 227–232.
Ивлев В.С. Экспериментальная экология питания рыб. М.: Пищепромиздат, 1955. 252 с.
Studier E.H., Sevick S.H. Live mass, water content, nitrogen and mineral levels in some insects from South-Central Lower Michigan // Comp. Biochem. Physiol. 1992. V. 103 A. № 3. P. 579–595.
Дополнительные материалы отсутствуют.