

Экология, 2020, № 6, стр. 465-470
Изменчивость экологических и морфологических характеристик темной полевки (Microtus agrestis) на Южном и Среднем Урале
М. И. Чепраков a, *, Н. Ф. Черноусова a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: cheprakov@ipae.uran.ru
Поступила в редакцию 23.05.2019
После доработки 10.02.2020
Принята к публикации 13.02.2020
Аннотация
Изменчивость экологических и морфологических характеристик темной полевки изучали на выборках, собранных в двух локалитетах Южного и трех локалитетах Среднего Урала. Полевки были отловлены в сходных биотопах – сосняках ягодниковых. В этих местообитаниях темная полевка – обычный вид. Полевки исследованных регионов характеризуются крупными размерами тела и черепа и высокой частотой дополнительной внутренней петли М1 моляра. Полученные результаты согласуются с гипотезой, в соответствии с которой предки современных темных полевок восточной сублинии цитохрома b пережили наиболее холодный период позднего плейстоцена в Уральском рефугиуме.
На протяжении последних лет изучение филогеографической структуры темной полевки (Microtus agrestis), выполненное рядом исследователей с использованием митохондриального (цитохром b) и ядерных локусов, выявило существование трех высоко генетически дифференцированных линий этого вида – португальской, южной и северной [1, 2]. Область распространения представителей северной линии охватывает Великобританию, Францию (кроме южной части), Данию, Германию, Польшу, Чехию, Румынию, страны Скандинавии и Россию [1, 2]. В пределах северной линии выделяют шесть сублиний: северобританскую, западную, французскую, центрально-европейскую, скандинавскую и восточную. Представители восточной сублинии встречаются в России, Финляндии, северной Швеции и северной Норвегии [3]. Согласно демографической модели, построенной на основе байесовского метода, в северной линии вида с начала голоцена происходит популяционный рост, продолжающийся до настоящего времени [2].
В Фенноскандии пространственное распределение особей восточной сублинии цитохрома b соответствует распределению северного подвида M. a. agrestis – они встречаются в Финляндии, северной Швеции и северной Норвегии [4]. Этот подвид характеризуется определенными эколого-морфологическими особенностями: крупными размерами тела и черепа, наличием дополнительной внутренней петли на М1 [5]. Высказана гипотеза [1], что вероятным рефугиумом, где предки представителей восточной сублинии выжили в течение последнего ледникового максимума, был Южный Урал.
Для фаун первой половины позднего плейстоцена Южного и Среднего Урала характерна относительно высокая доля темной полевки. В период позднего (ледникового) валдая темная полевка была очень редким представителем в фаунах этих регионов. В течение всего голоценового периода она входит в группу обычных видов [6–8]. В конце позднего плейстоцена темная полевка распространялась на Южном и Среднем Урале на север до 58° с.ш. В позднеледниковье, около 12 тыс. л. н., этот вид постепенно расселился на Северный Урал до 62° с.ш., а около 11 тыс. л. н. – на Полярный Урал до 67° с.ш. [9]. Эти данные позволяют заключить, что Уральский рефугиум, в котором предки современных темных полевок восточной сублинии цитохрома b пережили наиболее холодный период позднего плейстоцена [1], включал в себя не только Южный, но и Средний Урал. В позднеледниковое время началась экспансия представителей этой линии, которая в настоящее время прослежена в северном направлении.
Сопоставление морфологических особенностей у темных полевок из Южного и Среднего Урала может подтвердить гипотезу о происхождении восточной сублинии цитохрома b из Уральского рефугиума.
Цель настоящей работы – изучить изменчивость экологических и морфологических характеристик темной полевки на Южном и Среднем Урале. Для этого мы охарактеризовали биотопическую приуроченность и обилие полевок, изучили изменчивость размеров тела и черепа в зависимости от репродуктивно-возрастного статуса и пола. Исследовали морфотипическую изменчивость М1 в уральских популяциях темной полевки в связи с репродуктивно-возрастным статусом, полом, годом и районом обитания. Для оценки индивидуальной изменчивости жевательной поверхности М1 сравнивали встречаемость разных морфотипов левого и правого моляров. Сопоставили изученные морфологические параметры с таковыми из других регионов, в которых встречаются особи восточной сублинии цитохрома b, чтобы проверить, как наши данные согласуются с гипотезой о происхождении восточной сублинии цитохрома b из Уральского рефугиума.
МАТЕРИАЛ И МЕТОДЫ
На Южном Урале (SU) мелких млекопитающих отлавливали в лесных местообитаниях: в окрестностях г. Карабаш (SK, 55.51° с.ш., 60.10° в.д., 1983–1987 гг.) и Ильменском заповеднике (IR, 55.01° с.ш., 60.16° в.д., 1986 г.). На Среднем Урале (MU) – в естественном лесном насаждении, расположенном в 50 км на юго-восток от г. Екатеринбурга (окрестности пос. Двуреченск, SD, 56.60° с.ш., 61.04° в.д., 1983–1987 гг. – SD1 и 1991–2016 гг. – SD2) и в двух лесопарках г. Екатеринбурга: в Юго-Западном (SW, 56.80° с.ш., 60.53° в.д., 1990–2012 гг.) и Калиновском (на северо-востоке города – NE, 56.92° с.ш., 60.64° в.д., 1990–2016 гг.). Всего отработано 27 550 лов.-сут. Отловы проводили в летний период методом ловушко-линий (в 1983–1987 гг. – живоловками, в 1990–2016 гг. – капканчиками) с одноразовой проверкой в сутки. В качестве приманки использовали хлеб, обжаренный в подсолнечном масле. Общее количество пойманных темных полевок составило 445 экз., исследованных черепов – 332, зубов М1 – 664 (правые и левые). Различали три репродуктивно-возрастные группы: прибылые неполовозрелые, прибылые половозрелые и зимовавшие. Группы прибылых особей различали между собой по состоянию генеративной системы. В качестве возрастных критериев использовали массу и размеры тела, краниометрические и одонтологические признаки: масса тела (BM), длина тела (LB), длина хвоста (LT), кондилобазальная длина черепа (CBL), длина верхнего зубного ряда (LMT) [10, 11].
У темной полевки, как и у других грызунов зоны бореальных лесов, наблюдается закономерное изменение репродуктивно-возрастной структуры выборок в течение летнего сезона. В частности, к концу лета снижается доля зимовавших особей и возрастает доля прибылых [10]. Выборки SD1, SW и NE преимущественно состоят из особей, отловленных в середине лета, и имеют сходную долю зимовавших особей (табл. 1). Выборки полевок из Южного Урала объединили, поскольку они имеют сходную репродуктивно-возрастную структуру. В выборке SU преобладают зимовавшие особи, отловленные в начале лета, – она имеет наибольшую их долю. В выборке SD2 в большей мере представлены позднелетние прибылые особи. Ни в одной из выборок репродуктивно-возрастные группы не различаются по соотношению самцов и самок. Сами выборки также не различаются по соотношению полов.
Таблица 1.
Репродуктивно-возрастная и половая структура выборок темной полевки Южного и Среднего Урала (над чертой – экз., под чертой – %)
| Группа | Выборка | ||||
|---|---|---|---|---|---|
| SU | DSD1 | DSD2 | SW | NE | |
| Прибылые неполовозрелые Прибылые половозрелые Зимовавшие |
7/18 3/8 28/74 |
22/37 13/22 24/41 |
88/58 28/19 34/23 |
11/26 9/22 22/52 |
13/30 13/30 17/40 |
| Самцы Самки |
20/53 18/47 |
23/39 36/61 |
77/51 73/49 |
15/36 27/64 |
17/40 26/60 |
Собранные черепа полевок хранятся в Музее ИЭРиЖ УрО РАН, г. Екатеринбург.
Пакет программ StatSoft STATISTICA for Windows 6.0 использовали для анализа изменчивости размеров тела и черепа – дисперсионный анализ и t-статистику, морфотипической изменчивости М1 – логлинейный анализ и статистику χ2.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Биотопическая приуроченность и обилие темной полевки на Южном и Среднем Урале. В среднегорьях Южного Урала представлено несколько высотных поясов растительности – от горно-степного до горно-тундрового. Наибольшую площадь занимает горно-лесной пояс [6].
Согласно литературным данным [6, 12], численность темной полевки в разных биотопах различается. В пределах горно-степного пояса темная полевка обычна в пойменных луговых ассоциациях, в зарослях крупноплодных деревьев и кустарников, приуроченных к поймам ручьев и рек. Она изредка встречается в кустарниковых каменистых степях, а также в лесных колках и каменистых россыпях горно-лесостепного пояса, многочисленна в густых пойменных березняках горно-лесостепного пояса [6, 12]. В горно-лесном поясе темная полевка многочисленна в смешанных сосново-березовых лесах и на недавних вырубках. Она является обычным видом в нижней и верхней тайге и редким видом ‒ в каменистых россыпях этого пояса.
В еловых мелколесьях и березовых криволесьях подгольцового пояса и каменистых россыпях горно-тундрового пояса темная полевка редка, а обычна в травяно-моховых тундрах горно-тундрового пояса [6, 12].
В низкогорьях Среднего Урала темная полевка обычна в ельниках хвощово-осоково-сфагновых, сосняках осоково-сфагновых, ельниках-кедровниках сфагново-хвощовых, березняках осоково-травяных и на лесных полянах [13–15], редка в ельниках хвощово-папоротниковых и зеленомошно-мелкотравных, пихто-ельниках высокотравных, березняках вейниково-мелкотравных [14, 15].
Биотопы, в которых мы проводили отловы животных на Южном и Среднем Урале, сопоставимы и относятся к одному типу лесов – ягодниковым соснякам: зеленомошно-ягодниковым (IR, SK, SD) и ягодниковым (SW, NE) [16–19]. Значительное развитие травянистой растительности выступает непременным условием присутствия темной полевки в лесных местообитаниях [20]. Как известно, зеленые части травянистых растений и кустарничков преобладают в питании этого вида круглый год – их встречаемость в составе пищи колеблется по сезонам от 83 до 92% [10].
Сравнение оценок численности (ос. на 100 лов.-сут) темной полевки между регионами в синхронные периоды (за 1983–1987 гг.) выявило сходство их средних значений: 2.59 ± 0.31 (SD1, n = 70) и 2.96 ± 0.47 (SU, n = 40). Годовые значения численности колебались в сходных пределах – от 0.4 до 5.5 (SD1) и от 0 до 6.7 (SU).
В 1991–1995 гг. на Среднем Урале численность темной полевки для SD2 составляла 5.49 ± 0.49, n = 128, для SW – 2.87 ± 0.44, n = 43, для NE – 1.60 ± 0.33, n = 24. После 1995 г. она стала ниже: SD2 – 1.07 ± 0.12, n = 78, SW – 0.63 ± 0.11, n = 32, NЕ – 0.52 ± 0.09, n = 30, p < 0.01. Это снижение мы связываем с увеличением рекреационной нагрузки после 1995 г. В период 2010–2012 гг. в локалитетах SD, SW и NE проективное покрытие злаками еще составляло 10–14% [19].
Несмотря на флуктуации численности, в ягодниковых сосняках темная полевка обычна как на Южном, так и на Среднем Урале.
Изменчивость размеров тела и черепа темных полевок в зависимости от репродуктивно-возрастного статуса (три градации) и пола изучали с помощью дисперсионного анализа на объединенной выборке. Гендерные различия не были обнаружены для массы тела (BM), длины тела (LB) и длины верхнего зубного ряда (LMT) темной полевки (F1.326 < 1.1, p > 0.29). В среднем по репродуктивно-возрастным группам кондилобазальная длина черепа (CBL) оказалась выше у самок (25.15 ± 0.08), чем у самцов (24.81 ± 0.08, F1.255 = 8.6, p < 0.01), а длина хвоста (LT) – выше у самцов (30.2 ± 0.3), чем у самок (29.4 ± 0.3, F1.326 = = 5.0, p < 0.03). Различия хорошо выражены между всеми репродуктивно-возрастными группами по всем признакам (F2.326 >180, p < 0.001).
Зимовавшие темные полевки из нашей объединенной выборки (табл. 2) имеют сходную (t < 1.5, p > 0.15) массу тела (41.3 ± 0.8 г), длину тела (118.7 ± 1.0 мм) и хвоста (33.7 ± 1.0 мм) с полевками с севера европейской части России (Вологодская обл., n = 63) [21], а их пределы также совпадают: BM – 21–61, LB – 102–145, LT – 26–42 [21]. Похожие пределы изменчивости длины тела и хвоста имеют полевки с севера Сибири: LB – 101–138 и LT – 30–42, n = 78; на Алтае и Саянах: LB – 110–134 и LT – 29–41, n = 84 [22]. Пределы изменчивости кондилобазальной длины черепа и длины верхнего зубного ряда у полевок из нашей объединенной выборки совпадают (см. табл. 2) с пределами у темных полевок с Европейского Севера России: CBL – 26.0–28.2 и LMT – 6.2–7.0, n = 78, с севера Сибири: CBL – 26.2–28.3 и LMT – 6.3–6.9, n = 118 и с Алтая и Саян: CBL – 26.4–29.0 и LMT – 6.0–6.7, n = 84 [22]. Следовательно, крупные размеры тела и черепа, свойственные северному подвиду M. a. agrestis, присущи как полевкам Европейского Севера России и севера Сибири, так и полевкам Южного и Среднего Урала, Алтая и Саян.
Таблица 2.
Изменчивость средних размеров тела и черепа темных полевок Южного и Среднего Урала в зависимости от репродуктивно-возрастного состояния (над чертой – M ± SE, под чертой – пределы, в скобках – n)
| Признаки | Репродуктивно-возрастные группы | ||
|---|---|---|---|
| прибылые неполовозрелые |
прибылые половозрелые |
зимовавшие | |
| Масса тела, г | $\frac{{18.7 \pm 0.5}}{{10--24\left( {141} \right)}}$ | $\frac{{31.8 \pm 0.7}}{{25 - 52\left( {66} \right)}}$ | $\frac{{40.8 \pm 0.5}}{{25 - 61\left( {125} \right)}}$ |
| Длина тела, мм | $\frac{{90.6 \pm 0.6}}{{75 - 100\left( {141} \right)}}$ | $\frac{{106.8 \pm 0.9}}{{97 - 121\left( {66} \right)}}$ | $\frac{{118.1 \pm 0.6}}{{100 - 140\left( {125} \right)}}$ |
| Длина хвоста, мм | $\frac{{25.8 \pm 0.3}}{{18 - 34\left( {141} \right)}}$ | $\frac{{29.9 \pm 0.4}}{{24 - 37\left( {66} \right)}}$ | $\frac{{33.7 \pm 0.3}}{{25 - 41\left( {125} \right)}}$ |
| Кондилобазальная длина черепа, мм | $\frac{{23.25 \pm 0.09}}{{20.5 - 25.4\left( {93} \right)}}$ | $\frac{{24.93 \pm 0.12}}{{24.2 - 25.6\left( {59} \right)}}$ | $\frac{{26.76 \pm 0.09}}{{25.6 - 28.8\left( {109} \right)}}$ |
| Длины верхнего зубного ряда, мм | $\frac{{5.86 \pm 0.02}}{{5.5 - 6.3\left( {141} \right)}}$ | $\frac{{6.17 \pm 0.02}}{{6.0 - 6.3\left( {66} \right)}}$ | $\frac{{6.53 \pm 0.02}}{{6.3 - 7.0\left( {125} \right)}}$ |
Изменчивость жевательной поверхности М1 моляра. Основной составляющей изменчивости жевательной поверхности этого моляра у темной полевки является наличие дополнительной внутренней петли. Мы выделили семь морфотипов по степени выраженности дополнительной внутренней петли на этом моляре (рис. 1 ), которые сгруппировали в три категории: 1 – отсутствие дополнительной внутренней петли (A и B), 2 – открытая петля (C и D), 3 – замкнутая петля (E, F и G).
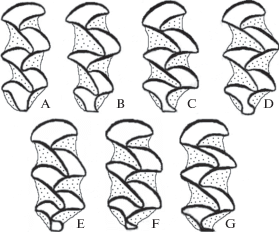
Репродуктивно-возрастное состояние (χ2 = 6.2, df = 4, p = 0.19), пол (χ2 = 4.3, df = 2, p = 0.12) и лево- и правостороннее положение зуба в челюсти (χ2 = 0.3, df = 2, p = 0.86) не влияют на распределение по категориям морфотипов М1 моляра. Выборки из Южного Урала однородны по частотам категорий морфотипов при сравнении по годам и локалитетам (χ2 = 8.0, df = 4, p = 0.09), выборки из лесопарков г. Екатеринбурга также однородны при сравнении по годам и локалитетам (χ2 = 9.3, df = 8, p = 0.32; рис. 2). В локалитете SD обнаружена неоднородность по частотам. Так, в выборке 1991 г. доля замкнутых петель составляет 78% (n = 78), а в выборке 1994 г. – 38% (n = 32, χ2 = 15.9, df = 1, p < 0.0001). В раннелетней выборке 2006 г. доля замкнутых петель равна 87% (n =30), а в позднелетней – 37% (n = 60, χ2 = 20.4, df = 1, p < 0.0001). Следовательно, в отдельном локалитете могут возникать существенные межгодовые и сезонные различия в частоте категорий морфотипов М1 моляра.
Рис. 2.
Распределение темной полевки по морфотипам М1 моляра в локалитетах Южного и Среднего Урала. Южный Урал: IR – Ильменский заповедник, SK – окрестности г. Карабаш; Средний Урал: SD – окрестности пос. Двуреченск, SW – лесопарк Юго-Западный, NE – лесопарк Калиновский.
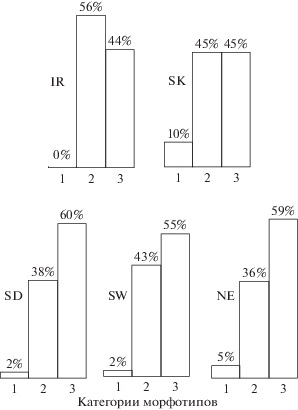
Суммарные выборки из локалитета SD (n = 418) и лесопарков г. Екатеринбурга (n = 170) сходны по частотам категорий морфотипов (χ2 = 0.83, df = 2, p = 0.67), в то время как суммарные выборки с Южного (n = 76) и Среднего (n = 588) Урала статистически различаются как по частоте первой категории морфотипов (SU – 7.9%, MU – 2.7%, χ2 = 5.6, df = 1, p < 0.02), так и по частоте третьей категории (SU – 45%, MU – 59%, χ2 = 5.5, df = 1, p < 0.02). Однако в выборках из обоих изучаемых регионов частота дополнительной внутренней петли М1 моляра показывает существенное доминирование (92–97%), что свойственно северному подвиду M. a. agrestis [5].
Наши данные по частоте дополнительной внутренней петли М1 моляра мы сравнили с данными для зимовавших темных полевок из Нижнего Приобья (87.7 ± 3.8%, n = 81) и Полярного Урала (93.0 ± 2.3%, n = 129). Они имеют высокую частоту дополнительной внутренней петли этого моляра, хотя у прибылых полевок она оказалась несколько ниже (77.3 ± 3.3%, n = 163) [23]. На севере европейской части России эта частота составляет 98 ± 1.6%, n = 78 [22], в Финляндии и северной Скандинавии она достигает 68 ± 9.9%, n = 22 [24], а на Алтае и Саянах – 51 ± 5.5%, n = 84 [22]. Подобное распределение частот данного признака, который в какой-то мере может выступать в качестве маркера восточной сублинии цитохрома b [4, 5], может свидетельствовать о том, что экспансия темных полевок из Южно-Среднеуральского рефугиума происходила не только в западном, но и в восточном направлении.
Таким образом, наши данные свидетельствуют о том, что в ягодниковых сосняках Южного и Среднего Урала темная полевка обычный вид. Мы установили высокий уровень сходства морфологических характеристик темной полевки из Южного и Среднего Урала с выборками из других регионов, в которых встречаются особи восточной сублинии цитохрома b. Полученные результаты согласуются с гипотезой о происхождении восточной сублинии цитохрома b из Южно-Среднеуральского рефугиума, где предки представителей этой сублинии пережили последний ледниковый максимум, а также дают основание предполагать, что экспансия темных полевок из этого рефугиума происходила не только в западном, но и в восточном направлении.
Работа выполнена в рамках государственного задания № АААА-А19-119031890087-7 Института экологии растений и животных УрО РАН.
Список литературы
Jaarola M., Searle J.B. Phylogeography of field voles (Microtus agrestis) in Eurasia inferred from mitochondrial DNA sequences // Mol. Ecol. 2002. V. 11. № 12. P. 2613–2621.
Paupério J., Herman J.S., Melo-Ferreira J. et al. Cryptic speciation in the field vole: a multilocus approach confirms three highly divergent lineages in Eurasia // Mol. Ecol. 2012. V. 21. № 24. P. 6015–6032.
Herman J.S., Searle J.B. Post-glacial partitioning of mitochondrial genetic variation in the field vole // Proc. R. Soc. Lond. B: Biol. Sci. 2011. V. 278. P. 3601–3607.
Jaarola M., Tegelström H. Colonization history of north European field voles (Microtus agrestis) revealed by mitochondrial DNA // Mol. Ecol. 1995. V. 4. № 3. P. 299–310.
Сиивонен Л. Млекопитающие Северной Европы. М.: Лесн. пром-сть, 1979. 232 с.
Смирнов Н.Г., Большаков В.Н., Косинцев П.А. и др. Историческая экология животных Южного Урала. Свердловск: УрО РАН, 1990. 245 с.
Смирнов Н.Г. Мелкие млекопитающие Среднего Урала в позднем плейстоцене и голоцене. Екатеринбург: Наука, 1993. 62 с.
Смирнов Н.Г. Грызуны Урала и прилегающих территорий в позднем плейстоцене и голоцене: Автореф. дис. … докт. биол. наук. Свердловск, 1994. 58 с.
Бачура О.П., Косинцев П.А. Ареалы млекопитающих на Урале в конце позднего плейстоцена // Зоол. журн. 2019. Т. 98. Вып. 8. С. 933–948.
Ивантер Э.В. Популяционная экология мелких млекопитающих таежного Северо-Запада СССР. Л.: Наука, 1975. 247 с.
Бобринский Н.А., Кузнецов Б.А., Кузякин А.П. Определитель млекопитающих СССР. М.: Просвещение, 1965. 382 с.
Большаков В.Н., Балахонов В.С., Бененсон И.Е. и др. Мелкие млекопитающие Уральских гор (экология млекопитающих Урала). Свердловск: УНЦ АН СССР, 1986. 101 с.
Бердюгин К.И., Кузнецова И.А., Шарова Л.П. Сообщества грызунов низкогорий Среднего Урала // Проблемы заповедного дела. Екатеринбург: “Екатеринбург”, 1996. С. 35–39.
Кузнецова И.А., Коурова Т.П., Жумашева А.М., Суслова В.В. Результаты долговременных учетов полевок Висимского заповедника // Исследования эталонных природных комплексов Урала. Екатеринбург: “Екатеринбург”, 2001. С. 302–306.
Марин Ю.Ф. Основные результаты учетов мелких млекопитающих на постоянных учетных линиях ловушек в Висимском заповеднике в 1982–2000 гг. // Исследования эталонных природных комплексов Урала. Екатеринбург: “Екатеринбург”, 2001. С. 337–346.
Колесников Б.П., Зубарева Р.С., Смолоногов Е.П. Лесорастительные условия и типы лесов Свердловской области. Свердловск: УНЦ АН СССР, 1973. 176 с.
Горчаковский П.Л., Золотарева Н.В., Коротеева Е.В., Подгаевская Е.Н. Фиторазнообразие Ильменского заповедника в системе охраны и мониторинга. Екатеринбург: “Гощицкий”, 2005. 192 с.
Коротеева Е.В., Вейсберг Е.И., Куянцева Н.Б. Оценка состояния лесной ценофлоры в зоне воздействия Карабашского медеплавильного комбината (Южный Урал) // Изв. Самар. НЦ РАН. 2011. Т. 13. № 1(4). С. 1005–1011.
Черноусова Н.Ф., Толкач О.В., Добротворская О.Е. Сообщества мелких млекопитающих в урбаногенно-нарушенных лесных экосистемах // Экология. 2014. № 6. С. 439–447. [Chernousova N.F., Tolkach O.V., Dobrotvorskaya O.E. Small mammal communities in forest ecosystems affected by urbanization // Rus. J. Ecol. 2014. V. 45. № 6. P. 490–497.]
Ивантер Э.В., Курхинен Ю.П., Соколов А.В. Экология темной полевки (Microtus agrestis L.) в коренных и антропогенных ландшафтах восточной Фенноскандии // Экология. 2013. № 3. С. 189–196. [Ivanter E.V., Kurkhinen Y.P. Sokolov A.V. Ecology of the field vole (Microtus agrestis L.) in indigenous and anthropogenic landscapes of Eastern Fennoscandia // Rus. J. Ecol. 2013. V. 44. № 3. P. 213–220.]
Башенина Н.В. Пути адаптаций мышевидных грызунов. М.: Наука, 1977. 355 с
Огнев С.И. Звери СССР и прилежащих стран. Т. 7. Грызуны. М.; Л.: Изд-во АН СССР, 1950. 706 с.
Бойкова Ф.И. Циклы численности и морфологические особенности темной полевки в Субарктике Западной Сибири // Численность и распределение наземных позвоночных Ямала и прилегающих территорий. Свердловск: УНЦ АН CCCP, 1981. С. 63–79.
Reichstein H., Reise D. Zur Variabilität des Molaren-Schmelzschlingenmusters der Erdmaus, Microtus agrestis (L.) // Zeitschrift für Säugetierkunde. 1965. Bd 30. S. 36–47.
Дополнительные материалы отсутствуют.