

Экология, 2020, № 6, стр. 457-464
Новые виды рыб в водоемах Европейского Северо-Востока России
А. П. Новосёлов *
Федеральный исследовательский центр комплексного изучения Арктики Уральского отделения РАН
(ФИЦКИА Уро РАН)
163000 Архангельск, наб. Северной Двины, 23, Россия
* E-mail: alexander.novoselov@rambler.ru
Поступила в редакцию 12.04.2020
После доработки 29.04.2020
Принята к публикации 06.05.2020
Аннотация
Проанализированы изменения состава ихтиофауны Европейского Северо-Востока России под воздействием биологических инвазий. Установлено, что в водоемах региона чужеродные рыбы-вселенцы появились в результате проведения акклиматизационных работ (дальневосточная горбуша, печорская пелядь в бассейне Белого моря, северодвинская стерлядь и сибирский осетр в р. Печора), направленной (судак) и случайной (головешка-ротан) интродукции, а также саморасселения каспийских (белоглазка и жерех) или балтийских (жерех) видов. Результаты анализа многолетних изменений пресноводной ихтиофауны показали, что появление чужеродных видов рыб в водоемах Европейского Северо-Востока России привело к увеличению разнообразия ихтиофауны, появлению трофической конкуренции с нативными видами, снижению рыбохозяйственного статуса водоемов лососевых рек за счет увеличения риска гибели молоди атлантического лосося и потенциальному ухудшению эпизоотической ситуации в низовье р. Северная Двина. В то же время в результате успешной акклиматизации печорской пеляди в водоемах бассейна Белого моря была разработана схема организации полносистемных озерных сиговых хозяйств с формированием собственных маточных стад пеляди, а также поликультуры с местным сигом. Натурализовавшиеся в реках Онега и Печора осетровые виды рыб со временем могут разнообразить перечень промысловых рыб.
Изменения в водных экосистемах, вызываемые природными и антропогенными факторами, по-разному влияют на гидробионтов: одни виды в меняющихся условиях испытывают угнетение, другие продолжают находиться в состоянии гомеостаза, третьи начинают расширять границы естественных ареалов. Это может быть как результатом целенаправленной интродукции видов с целью акклиматизации в новых местах, так и следствием спонтанного саморасселения.
Под биологическими инвазиями понимаются случаи проникновения живых организмов в экосистемы, расположенные за пределами их первоначального (обычно естественного) ареала. Инвазивные виды, называемые “вселенцами”, “адвентивными” или “чужеродными” видами, могут воздействовать на аборигенные популяции, виды и сообщества, приводя к необратимым изменениям. Термин “инвазивный вид” определен в Конвенции о биологическом разнообразии (1992), подразумевая натурализовавшиеся виды, наносящие вред аборигенной (нативной) фауне и экосистемам. Чужеродные виды рыб могут наносить существенный ущерб рыбному хозяйству, выступая в качестве возбудителей или переносчиков заболеваний, а также конкурентно-активных видов, вытесняющих местные промысловые виды рыб. Так, головешка-ротан Perccottus glenii Dybowski, 1877 широко распространился в Северной Евразии, вызывая угнетение многих популяций аборигенных видов беспозвоночных, рыб и амфибий [1]. В некоторых изолированных водоемах Республики Коми ротан стал доминировать по численности, потеснив золотого карася [2], в водоемах Белоруссии встречается в бассейнах всех основных рек [3]. В научной литературе есть ряд примеров результатов негативного воздействия судака на нативные виды рыб [4].
Поэтому выявление инвазивных видов и исследование их влияния на нативные сообщества и экосистемы – важные задачи. Изучение современного состояния и динамики рыбного населения северных водоемов под воздействием природных (климатические изменения) и антропогенных (техногенная нагрузка) факторов представляет несомненный научный и практический интерес.
Цель настоящей работы – попытка проведения обзора новых видов (как чужеродных, так и инвазивных), появившихся в пресноводных водоемах Европейского Северо-Востока России в результате акклиматизационных работ, целенаправленной и ненаправленной (спонтанной) интродукции, а также саморасселения.
МАТЕРИАЛ И МЕТОДЫ
Регион Европейского Северо-Востока России объединяет территорию, северная граница которой проходит по морской береговой линии западной части Белого, юго-восточной части Баренцева и юго-западной части Карского морей (включая бассейн р. Кары) (рис. 1 ). Восточная граница территории совпадает с уральским Печоро-Обским водоразделом, южная – с водоразделом Баренцева и Каспийского морей. Западная граница территории проходит по западному контуру бассейна р. Онега, т.е. практически совпадает с границей между Балтийским кристаллическим щитом и Русской платформенной равниной. Этот район включает в себя побережья морей, омывающих территорию Архангельской области и Ненецкого автономного округа, а также наиболее крупные речные бассейны Европейского Северо-Востока России – рек Онеги, Северной Двины, Мезени и Печоры.
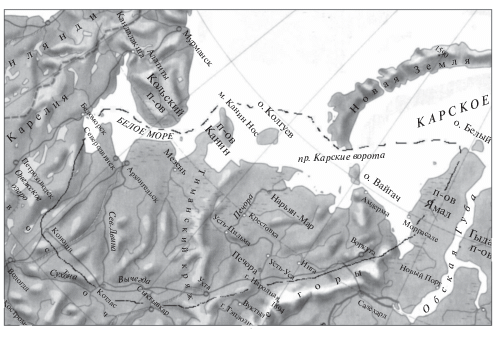
Материалом для настоящей работы послужили результаты многолетних исследований ихтиофауны Европейского Северо-Востока России за период с 1980-х гг., а также данные публикаций о появлении новых видов в составе ихтиофауны региона. Сбор материалов проводился на водоемах Онежского полуострова, озерах Архангельской области, бассейнах рек Северной Двины и Печоры (на территории Ненецкого автономного округа и Республики Коми). Систематический статус рыб представлен в соответствии с “Атласом пресноводных рыб России” [5] и выверен по справочному пособию “Рыбы России…” [6].
Новые виды отлавливали и анализировали в ходе комплексных ихтиологических съемок как активными, так и пассивными орудиями лова. Использовался закидной невод длиной 80 м с размером ячеи в кутке 16–20 мм и набор ставных жаберных сетей в количестве 10 шт. с размером ячеи от 15 до 70 мм. Анализ выловленной рыбы включал определение соотношения видов в составе контрольных уловов, распределения рыб во временном (разные годы) и пространственном (различные водные объекты) аспектах, а также биологических параметров чужеродных видов.
Биологический анализ проводился на свежем материале по методике И.Ф. Правдина [7] с учетом рекомендаций Ю.С. Решетникова [8] применительно к сиговым рыбам. У пойманных рыб измеряли промысловую длину тела (см – от вершины рыла до конца чешуйного покрова), взвешивали массу тела (г), визуально определяли пол и стадию зрелости гонад в баллах. Возраст рыб определен по чешуе, которую отбирали из 2–3 рядов над боковой линией перед спинным плавником. Определение возраста и измерение радиусов годовых колец по переднему краю чешуи проведено с помощью бинокуляра МБС-9. Темп линейного и весового роста сига анализировали по традиционным методикам [9] с использованием методических указаний М.В. Мины [10]. При этом рассчитывали разницу между величиной последнего прироста тела рыбы и величиной прироста предыдущего сезона.
Питание рыб анализировали в соответствии с “Методическим пособием по изучению питания и пищевых отношений рыб в естественных условиях” [11]. Количественный состав пищи выражали в процентном отношении веса отдельных кормовых объектов к весу содержимого желудочно-кишечных трактов. Интенсивность питания рассчитывали в виде общих индексов наполнения желудочно-кишечных трактов и выражали в продецимиллях (‱).
При рассмотрении пищевых (конкурентных) взаимоотношений чужеродных и аборигенных видов рыб анализировали степень пищевого сходства (СП) сравниваемых видов рыб [12], а также индекс перекрывания пищевых ниш (Сλ) [13]. Индекс пищевого сходства рассчитывали как сумму наименьших величин из видового состава рациона сравниваемых рыб (рационы в %): при полном совпадении индекс равен 100%, при отсутствии совпадения 0%. Для определения степени перекрывания пищевых ниш разных видов рыб рассчитывали индекс Хорна:
где xi и yі – значения отдельных компонентов в пищевых комках рыб, %.При анализе собранных материалов мы придерживались следующих общепринятых понятий в трактовке А.Ф. Карпевич [14]:
Интродукция – перенос организмов с целью введения их в новую область, водоем, культуру. Она является первым этапом процесса акклиматизации, но не обязательно заканчивается акклиматизацией интродуцента. Интродукции видов могут быть намеренными, когда чужеродный вид намеренно перемещается или выпускается за пределы его естественного распространения (ареала), или ненамеренными, когда интродукция происходит по какой-либо иной причине, связанной с деятельностью человека [15].
Акклиматизация – процесс приспособления интродуцированных особей и их потомства к новым условиям среды, а также формирования в них новой популяции.
Натурализация – конечная фаза процесса акклиматизации, когда вселенец приспособился к новым условиям, определились его ниша и взаимоотношения с аборигенами в экосистеме водоёма, установилось подвижное равновесие численности новой популяции и выявилась возможность ее использования в кормовых или промысловых целях.
Саморасселение – самостоятельное вселение водных организмов с последующей их акклиматизацией в новом водоеме.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
За последние полвека в составе ихтиофауны пресноводных водоемов рассматриваемой территории появился ряд новых видов, которые не обитали здесь ранее, в результате: 1) проведения акклиматизационных работ (дальневосточная горбуша в бассейнах Белого и Баренцева морей, печорская пелядь в Беломорском бассейне, северодвинская стерлядь и сибирский осетр в р. Печора); 2) намеренной интродукции (судак в бассейнах рек Онеги и Северной Двины); 3) ненамеренной (спонтанной) интродукции (ротан в озерных экосистемах Архангельской области и Республики Коми); 4) саморасселения каспийских (белоглазка, жерех) или балтийских (жерех) видов в р. Северная Двина.
Акклиматизация
Горбуша Oncorhynchus gorbusha (Walbaum, 1792). История акклиматизации дальневосточной горбуши на Европейском Севере России насчитывает уже более полувека. За этот период она широко распространилась в водоемах Кольского п-ва, Карелии, Архангельской области и Ненецкого автономного округа, отмечается у берегов Норвегии, Швеции, Исландии и Шпицбергена, встречается в бассейне р. Печора, на востоке доходит до бассейна р. Таз [16]. В реки Белого моря заходят многочисленные стада горбуши линии нечетных лет, сформированные от естественного нереста. Биологический эффект акклиматизации выразился в достаточно высокой численности натурализовавшейся горбуши как промыслового объекта [17].
В ходе организации и проведения акклиматизационных работ с особой остротой вставал вопрос о конкуренции горбуши с аборигенными видами – атлантическим лососем и кумжей. Для исключения такой конкуренции в 2005 г. Россия подписала конвенцию о недопущении вселения новых видов лососевых рыб в водоемы, где обитает семга Salmo salar Linnaeus, 1758.
Пелядь Coregonus peled (Gmelin, 1789). С 1972 г. проводились работы по вселению печорской пеляди в водоемы бассейна Белого моря. Собранную на р. Печора икру инкубировали на Онежском рыбоводном заводе, а полученными личинками и подрощенной молодью зарыбляли разнотипные водоемы Архангельской области. В общей сложности в 1970–1980-е гг. было выпущено более 6 млн сеголеток и около 60 млн личинок пеляди, и она успешно освоила новый для нее ареал [18–20].
В новых условиях проявилась широкая экологическая пластичность пеляди и произошло образование трех ее экологических форм – озерной, озерно-речной и речной. Типично озерная пелядь, отличающаяся большей высокотелостью, стала нереститься и нагуливать в озерах и в р. Онегу не выходила. Более низкотелая и мелкая озерно-речная пелядь нагуливала в озерах, а осенью мигрировала в реку на нерест. Типично речная пелядь с прогонистой формой тела освоила низовье р. Онега, где перешла на смешанное питание (вплоть до хищничества молодью окуня), и характеризовалась замедленным ростом как по сравнению с озерной, так и исходной материнской формой [18].
Скорость роста пеляди определялась в основном характером ее питания. Типично зоопланктонное питание пеляди в озерах с достаточной кормовой базой обусловило формирование озерной формы, превосходящей по темпу роста не только пелядь из других водоемов нового ареала, но и пелядь материнского печорского стада. Речная пелядь, перейдя преимущественно на бентосный характер питания, значительно отставала в росте как от озерной формы, так и от материнской печорской. В новых условиях возраст ее полового созревания сократился на один-два года, однако пелядь при этом характеризовалась меньшими значениями абсолютной плодовитости [18].
Характер питания пеляди зависит от кормовой базы водоемов. В озерах с достаточным развитием зоопланктона бентос практически не имел значения для питания пеляди и основу пищи составлял зоопланктон, в основном веслоногие и ветвистоусые ракообразные. В малокормных озерах и речных условиях большое значение в питании пеляди имел бентос. В новых условиях обитания конкуренция пеляди с аборигенными видами была слабой благодаря сильному расхождению спектров их питания [21].
В речных условиях ни один из проанализированных видов рыб, в том числе и пелядь, не имели планктонного характера питания. Однако пищевая конкуренция снижалась в результате того, что пелядь использовала в пищу в основном моллюсков, а местные рыбы потребляли личинок хирономид, стрекоз, ручейников и поденок. Только проходной сиг, потреблявший, как и пелядь, моллюсков, мог являться серьезным пищевым конкурентом пеляди. Однако во время нерестового хода вверх по реке сиг практически не питается, а его весенне-летнее пребывание в нижнем течении реки носит периодический характер, что исключает длительное состояние напряженности пищевых отношений между пелядью и сигом [10]. В озерах пелядь не конкурировала с представителями местной ихтиофауны, поскольку она заняла трофическую нишу планктофага, недоиспользовавшуюся до ее вселения. При общем питании ветвистоусыми ракообразными пелядь потребляла в основном босмин, а местные уклея, плотва и мелкий окунь питались исключительно дафниями. Для бентофагов (сига и леща), а также эврифага ерша зоопланктон являлся лишь случайной пищей.
Результаты вселения печорской пеляди в разнотипные водоемы Архангельской области показали ее широкие адаптивные возможности и могут рассматриваться как один из примеров положительных результатов акклиматизационных работ. Возможности ее хозяйственного использования в качестве объекта пастбищного выращивания в полносистемных сиговых озерных хозяйствах ограничиваются в настоящее время не экологическими, а лишь социально-экономическими факторами [18].
В водоемах Европейского Северо-Востока России осетровые виды рыб, акклиматизированные в новых условиях обитания, включают северодвинскую стерлядь и сибирского осетра.
Стерлядь Acipenser ruthenus Linnaeus, 1758 – единственный вид осетровых рыб, распространенный на Европейском Северо-Востоке России в бассейне р. Северная Двина. Долгое время появление здесь стерляди связывали со строительством каналов [22, 23]. Сторонники этой гипотезы объясняли проникновение стерляди в Двинский бассейн со стороны р. Вычегды функционированием Екатерининского канала, а именно прорывом шлюза высоким весенним паводком 1810 г. [24]. В то же время находка остатков стерляди в отложениях бассейна р. Онеги, датированных II–III тысячелетием до н. э., привела к предположению о проникновении ее также и в р. Северная Двина, где она, в отличие от онежской, сохранилась до наших дней [25, 26]. В настоящее время стерлядь встречается в реках Сухоне, Юге (с Лузой), Вычегде (с Сысолой), самой Двине и ее притоках Baгe и Пинеге [27].
В целях расширения ареала и акклиматизации северодвинскую стерлядь неоднократно выпускали в реки Печора (1933–1961 гг.), Мезень (1960–1963 гг.), Онега (1961–1968 гг.), Шуя (1968–1982 гг.). В первые годы после проведения акклиматизационных работ эффекта от них не наблюдалось, поскольку зарыбление проводилось разновозрастными особями и характеризовалось малыми объемами. В зарыбленных водоемах (реки Печора, Мезень, Онега) молодь стерляди встречалась единично, а взрослые особи, в том числе и половозрелые, попадали в орудия лова крайне редко. В настоящее время случаи поимки стерляди в р. Печора перестали быть единичными, встречается она и в ее притоке – р. Уса. Установлено, что ее нагул происходит в основном в нижнем и частично в среднем течении р. Печоры, а численность лимитируется лишь наличием естественных нерестилищ [28]. Примерно такая же ситуация наблюдается и в р. Онеге, где был отмечен случай поимки стерляди массой более 10 кг.
Сибирский осетр Acipenser baerii Brandt, 1869. В 1956 г. около сотни разновозрастных осетров, выловленных в р. Обь, были выпущены в р. Печора и в том же году несколько экземпляров были выловлены различными орудиями лова в рр. Уса и Колва [29]. До 2005 г., несмотря на активное рыболовство в р. Печора и ее притоках, а также регулярные ихтиологические исследования, осетр не встречался. В июле 2005 г. на разных участках главного печорского русла были пойманы 2 экз. осетра (массой 19.6 и 2.0 кг). Возраст крупного осетра составил 22 года, мелкого – 6 полных лет [30]. Осенью того же года еще 1 экз. осетра попал в плавную сеть в нижнем течении р. Печоры. В 2007–2019 гг. случаи поимки 4–7-летних осетров в нижнем течении р. Печоры стали регулярными, и все выловленные особи идентифицированы как сибирский осетр – Acipenser baerii Brandt.
Судя по хронологии событий, пойманные в р. Печора осетры появились в результате уже естественного воспроизводства интродуцированных взрослых особей. Об этом также свидетельствует и факт поимки сеголетка осетра при промысле ряпушки в 2006 г. в районе с. Усть-Цильма [20], т.е. уже можно говорить о завершившемся процессе натурализации вида в новых условиях обитания. Во всяком случае наличие в настоящее время в р. Печоре сибирского осетра следует считать достоверным фактом, что дает основание для его включения в состав ихтиофауны р. Печора в статусе редкого вида [30, 31].
Намеренная интродукция
Обыкновенный судак Stizostedion lucioperca (Linnaeus, 1758). Естественный ареал судака в водоемах Архангельской области ограничивается лишь рядом озер Балтийского бассейна. Известно, что судак не входил в состав рыбного населения водной системы р. Онега не только в прошлом веке, но и в тот период, когда в Онежском бассейне обитали рыбы теплолюбивого комплекса – до I тысячелетия до н.э. [25, 32]. В настоящее время он появился в р. Онега в результате вселения в оз. Воже (соседняя с Архангельской областью Вологодская область), которое проводилось в целях улучшения качественного состава озерной ихтиофауны [33]. Открытость озерно-речной системы позволила интродуцированному судаку через р. Свидь проникнуть в оз. Лача, являющееся истоком одной из крупных рек бассейна Белого моря – Онеги. После 2003 г. судак стал отмечаться в уловах в среднем течении р. Онега, где расположены основные нерестово-выростные угодья (НВУ) атлантического лосося. Установлено, что пик миграции покатников лосося совпадает по времени с окончанием нереста судака и началом его интенсивного питания. В 2001 г. в желудках нескольких выловленных судаков было отмечено до 5 экз. смолтов сёмги. На наш взгляд, с увеличением численности судака в Онежском бассейне его воздействие на аборигенные виды будет только усиливаться [34].
Известны примеры негативного влияния судака на нативные виды рыб. Так, в результате его вселения в оз. Балхаш один из местных промысловых видов – балхашский окунь (Perca schrenki) – был включен в Красную книгу МСОП [35], а в результате интродукции судака в озера Норвегии и увеличения его численности отмечалось значительное снижение в них численности плотвы.
В р. Северная Двина судак появился в результате направленной интродукции (вселения) в р. Сухону из оз. Кубенское, а затем распространился практически по всей реке вплоть до участков дельты и приустьевого взморья (губа Сухое море). В настоящее время можно говорить о произошедшей натурализации судака в Северодвинском бассейне, поскольку как в промысловых, так и контрольных орудиях лова отмечаются как производители (массой до 4 кг), так и разновозрастная молодь.
Непреднамеренная интродукция
Интродукция рыб за пределы естественных ареалов может иметь негативные последствия и привести к изменениям в водных экосистемах [4]. Северные водоемы малоустойчивы к воздействию внешних факторов природного и антропогенного характера [35].
Головешка-ротан Percottus glenii Dybowski, 1877. В конце 1990-х гг. этот вид был завезен аквариумистами в оз. Плесецкое, расположенное на закрытой территории космодрома “Плесецк” в Архангельской области в бассейне р. Северная Двина. Обнаружен также на территории Республики Коми в изолированных прудах в районе г. Сыктывкара в бассейне р. Вычегда [2]. Известно, что в новых условиях ротан переходит на хищное питание и обладает высоким темпом роста [36]. Как показали результаты проведенных нами ранее исследований [37], в оз. Плесецкое ротан действительно имел высокий темп роста. При значительной плотности популяции масса его разновозрастных особей колебалась от 1 до 469 г, при этом ротан характеризовался преимущественно хищным характером питания. В его рационе рыба (в т.ч. и особи своего вида) являлась доминирующим компонентом, составляя более 80% по весу от содержимого желудочно-кишечных трактов.
Саморасселение
В последние десятилетия в р. Северная Двина появились новые виды рыб, не обитавшие здесь ранее, – это обитатели бассейнов Каспийского и Балтийского морей, а именно белоглазка и жерех. Мигранты могли появиться в Северодвинском бассейне двумя путями: через реки Вычегду и (или) Сухону. В первом случае они по р. Каме и ее притокам (Вятке и Южной Кельтме) дошли до Екатерининского канала, который как судоходный уже не функционирует, но в годы большой водности миграции рыб по нему вполне возможны. Далее по р. Северная Кельтма эти виды могли проникнуть в р. Вычегда, и затем через р. Малая Северная Двина заселить весь Двинский бассейн. Второй путь возможного саморасселения каспийских видов пролегает через Рыбинское водохранилище, реки Шексна и Паразовица, по Паразовицкому каналу в оз. Кубенское и далее в р. Сухона, дающую при слиянии с р. Вычегда начало р. Малая Северная Двина. Кроме того, через Паразовицкий канал возможна миграция жереха и из водоемов Балтийского бассейна. Этот путь пролегает из Онежского озера через р. Вытегра в Волго-Балтийский канал, затем через р. Ковжа в оз. Белое, в р. Паразовица и далее по уже рассмотренной схеме проникновения в р. Сухона каспийских видов [38].
Белоглазка Abramis sapa (Pallas, 1814) является второстепенной промысловой рыбой нижнего течения р. Волга. Впервые была отмечена в р. Вычегда в 1971 г., затем появилась в р. Северной Двине, быстро увеличивая свою численность. В промысловой статистике долгое время включалась в состав уловов как молодь леща или густеры. В последнее десятилетие она стала встречаться практически по всей р. Северная Двина, распространившись к настоящему времени вплоть до дельтовой части реки и участков приустьевого взморья. Ситуация представляется весьма проблематичной, поскольку, являясь солоноватоводным видом, белоглазка может создать серьезную пищевую конкуренцию ценным промысловым видам аборигенного комплекса – карповым рыбам (лещ), а также северодвинскому сигу на его кормовых биотопах в дельтовой части реки и приустьевом взморье [38].
Анализ питания и пищевой конкуренции белоглазки с местными видами показал, что между ними действительно сложились достаточно напряженные взаимоотношения. Индекс пищевого сходства [12] белоглазки и сига составил 62.8%, индекс перекрывания пищевых ниш Сλ [13] – 89.9%. Это стало следствием того, что они в основном питаются сходными видами корма. Основу рациона обоих видов составляют водные личинки насекомых, суммарная доля которых составляет у белоглазки 70%, у сига 58%, а доминирующими кормовыми объектами были личинки хирономид – 53% у белоглазки и 50% у сига. Оба вида рыб активно потребляли водную растительность, доля которой в пищевом спектре белоглазки составляла 21%, сига – 10%. Моллюски присутствовали в желудочно-кишечных трактах обоих видов, но сиг потребляет их более интенсивно (18%) по сравнению с белоглазкой (1%). Такая же картина наблюдается и при сравнении характера питания белоглазки и леща (СП = 57.8%, Сλ = 84.1%). Это происходит в результате их питания личинками хирономид (соответственно 53 и 86%) и моллюсков (соответственно 1 и 10%) [39].
Обыкновенный жерех Aspius aspius (Linnaeus, 1758). Хищный представитель карповых рыб, появившийся в Северодвинском бассейне вслед за белоглазкой. В р. Вычегде является редким видом [31, 40 ]. В р. Северной Двине численность жереха также пока невелика, единично встречается на участках среднего течения реки, а также в устьевой части и на участках приустьевого взморья.
НЕКОТОРЫЕ АСПЕКТЫ ТРАНСФОРМАЦИИ ВОДНЫХ ЭКОСИСТЕМ ПРИ ПОЯВЛЕНИИ ИНВАЗИВНЫХ ВИДОВ
Успешно освоив новую среду обитания, адвентивные виды являются реальным фактором трансформации водных экосистем, воздействуя на популяции, виды и сообщества аборигенных фаун. Конкретное воздействие каждого чужеродного вида трудно прогнозируемо ввиду большого числа связанных с ним параметров [41]. Применительно к водоемам рассматриваемого нами региона экспертно можно гипотетически обозначить следующие возможные последствия:
1. Изменение видового разнообразия. В результате саморасселения в бассейн р. Северная Двина ряда рыб каспийского и балтийского комплексов (белоглазка, жерех) расширился видовой состав ихтиофауны. В стабильной экологической обстановке они способны освоить имеющиеся свободные экологические ниши, не приводя к депрессии аборигенной ихтиофауны, однако при дальнейшем потеплении и, как следствие, эфтрофировании речных бассейнов теплолюбивые вселенцы могут получить преимущества при воспроизводстве как фитофильные виды и дать вспышку численности. Поэтому вполне вероятна депрессия видов рыб лососево-сигового комплекса в результате ухудшения условий естественного воспроизводства. Иными словами, происходящая экспансия инвазивных видов рыб в бассейны северных рек представляет угрозу для сохранения естественной биологической сбалансированности видового состава ихтиофауны.
2. Усиление трофической конкуренции. Прежде всего может произойти обострение пищевой конкуренции рыб-вселенцев с промысловыми представителями нативной фауны. Так, появление белоглазки в бассейне р. Северная Двина привело к возникновению конкуренции с лещем и сигом. В условиях общей загрязненности речного бассейна и серьезного прессинга рыболовства современное состояние сига оценивается как депрессивное. Поэтому возникшую пищевую конкуренцию с саморасселившейся в бассейне р. Северная Двина белоглазкой следует расценивать как дополнительный негативный фактор, повышающий степень экологического риска для популяции северодвинского сига.
3. Снижение рыбохозяйственного статуса водоемов. При вселении хищных видов рыб возможно снижение рыбохозяйственного статуса водоемов или потеря отдельных видов, населяющих сопредельные водные объекты. Так, с появлением судака в р. Онега появился дополнительный фактор риска для молоди атлантического лосося, поскольку пик миграции покатников семги совпадает по времени с окончанием нереста судака и началом его интенсивного питания. В этой связи при дальнейшем распространении судака по Онежской речной системе и его экологическом прогрессе вполне вероятно снижение продуктивности как самой р. Онега (по лососю), так и промысловых озер в бассейне (по ряпушке).
4. Ухудшение эпизоотической ситуации на водоемах. Проникновение чужеродных видов в водоемы Севера может повлечь за собой и санитарно-биологическую опасность. Если в дельте Северной Двины еще десятилетие назад карповые виды (лещ, язь и плотва), пораженные обыкновенным лентецом, встречались единично, то сейчас это обычное явление. Конкретная роль в этом процессе инвазивных южных видов пока нами не выявлена и требует изучения.
ЗАКЛЮЧЕНИЕ
Попав в новые условия, инвазивные виды могут угнетать или вытеснять аборигенные виды из-за своей биологической агрессивности, более высокой жизнеспособности и большего адаптивного потенциала. Поэтому появление новых видов в водоемах Севера уже само по себе предполагает серьезную проблему. В водоемах Европейского Северо-Востока России чужеродные рыбы-вселенцы появились в результате проведения акклиматизационных работ (дальневосточная горбуша, печорская пелядь в бассейне Белого моря, северодвинская стерлядь и сибирский осетр в р. Печора), направленной (судак) и случайной (головешка-ротан) интродукции, а также саморасселения каспийских (белоглазка и жерех) или балтийских (жерех) видов.
Появление адвентивных видов рыб в пресноводных водоемах Европейского Северо-Востока России привело к увеличению разнообразия ихтиофауны, возникновению трофической конкуренции с нативными видами, снижению рыбохозяйственного статуса водоемов лососевых рек за счет увеличения риска гибели молоди атлантического лосося и потенциальному ухудшению эпизоотической ситуации в низовье р. Северная Двина. Однако появление новых видов не всегда ведет к негативным последствиям. В результате успешной акклиматизации печорской пеляди в водоемах бассейна Белого моря была разработана схема организации полносистемных озерных сиговых хозяйств с формированием собственных маточных стад пеляди, а также поликультуры с местным сигом. Натурализовавшиеся в реках Онега и Печора осетровые виды рыб со временем могут разнообразить перечень промысловых рыб.
Работа выполнена за счет средств целевой субсидии на выполнение государственного задания “Исследование закономерностей формирования пресноводной ихтиофауны Европейского Северо-Востока России в условиях меняющегося климата и воздействия антропогенных факторов” (№ 0332-2019-0001), № гос. регистрации – АААА-А19-119011690119-9.
Список литературы
Решетников А.Н. Современный ареал ротана (Odontobutidae, Pisces) в Евразии//Российский журн. биологич. инвазий. 2009. № 1. С. 22–34.
Бознак Э.И. Головешка-ротан Percottus glenii (Eleotridae) из бассейна р. Вычегды // Вопр. ихтиологии. 2004. Т. 44. № 5. С. 712–713.
Лукина И.И. Распространение ротана (Perccottus glenii Dybowski, 1877) в Беларуси // Российский журн. биологич. инвазий. 2011. № 2. С. 114–118.
Павлов Д.С., Савваитова К.А., Соколов Л.И., Алексеев С.С. Редкие и исчезающие животные. Рыбы. М.: Высшая школа, 1994. 333 с.
Атлас пресноводных рыб России. Под ред. проф. Решетникова Ю.С. М.: Наука, 2003. Т. 1. 378 с.; Т. 2. 252 с.
Романов В.И. Ихтиофауна России в системе рыб мировой фауны: Учебное пособие. Томск: Издательский Дом ТГУ, 2014. 410 с.
Правдин И.Ф. Руководство по изучению рыб. М.: Пищепромиздат, 1966. 376 с.
Решетников Ю.С. Экология и систематика сиговых рыб. М.: Наука, 1980. 301 с.
Чугунова Н.И. Руководство по изучению возраста и роста рыб. М.: Изд-во АН СССР, 1959. 164 с.
Мина М.В. Рост рыб (методы исследования в природных популяциях) // Рост животных. Зоология позвоночных: итоги науки и техники. М.: ВИНИТИ, 1973. Т. 4. С. 68–115.
Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях. М.: Наука, 1974. 254 с.
Шорыгин А. А. Питание и пищевые взаимоотношения рыб Каспийского моря. М.: Пищепромиздат, 1952. 286 с.
Horn H. Measurement of overlap in comparative ecological studies // Amer. Natur. 1966. № 100. P. 419– 424.
Карпевич А.Ф. Теория и практика акклиматизации организмов. М.: Пищевая пром-ть, 1975. 432 с.
Decision VI/23 COP6 of the Convention on Biological Diversity: Hague, 2002. [Электронный ресурс]: http://www.biodiv.org/decisions
Богданов В.Д., Кижеватов Я.А. Горбуша (Oncorhynchus gorbuscha Walbaum, 1792) – новый вид водных биологических ресурсов в Ямало-Ненецком национальном округе // Вестник Астраханского гос. технич. ун-та. Сер. Рыбное хоз-во. 2015. № 3. С. 7–14.
Боркичев В.С., Студенов И.И., Крылова С.С., Шаров А.А. Промысловое использование горбуши в Белом море // Биологические ресурсы Белого моря и внутренних водоемов Европейского Севера. Сыктывкар, 2003. С. 18–20.
Новоселов А.П. Морфо-экологическая изменчивость печорской пеляди при акклиматизации в водоемах Северо-Запада СССР: Автореф. дис. … канд. биол. наук. М., 1984. 24 с.
Новоселов А.П. Развитие товарного сиговодства ̶ путь повышения продуктивности озер Архангельской области // Рыбное хозяйство. 1991. № 3. С. 44– 47.
Новоселов А.П., Решетников Ю.С. Пелядь в новых местах обитания // Биология сиговых рыб. М.: Наука, 1988. С. 78–114.
Новоселов А.П. Пищевые отношения интродуцированной пеляди Coregonus peled (Gmelin)(Coregonidae) с аборигенными представителями рыбной части сообществ в озерных и речных условиях Архангельской области // Вопр. ихтиологии. 1987. Т. 25. Вып. 3. С. 458–465.
Догель В.А. Влияние акклиматизации рыб на распространение рыбной эпизоотии// Изв. ВНИОРХ. 1939. Т. 21. С. 112–115.
Иоганзен Б.Г. Стерлядь бассейна р. Оби // Труды Томского гос. ун-та. 1946. Т. 97. С. 67–72.
Остроумов Н.А. Рыбы среднего и нижнего течения Печоры // Докл. АН СССР. 1948. Т. 59. № 8. С.1497–1500.
Никольский Г.В. К истории ихтиофауны бассейна Белого моря // Зоол. журн. 1943. Т. 22. Вып. 1. С. 27–32.
Берг Л.С. О стерляди в бассейне Белого моря // Природа. 1945. № 6. С. 66.
Новоселов А.П. К вопросу о распространении стерляди на Европейском Северо-Востоке и перспективах ее искусственного воспроизводства в бассейне р. Северной Двины // Биологические ресурсы Белого моря и внутренних водоемов Европейского Севера. Петрозаводск, 1999. С. 263–266.
Захаров А.Б., Крылова В.Д., Осипова Т.С. Итоги и перспективы интродукции северодвинской стерляди Acipenser ruthenus в бассейн Печоры // Вопр. ихтиологии. 1998. Т. 38. № 6. С. 825–829.
Соловкина Л.Н. Рыбные ресурсы Коми АССР. Сыктывкар, 1975. 168 с.
Захаров А.Б., Туманов М.Д., Шалаев С.Н. Сибирский осетр Acipenser baerii в реке Печора // Вопр. ихтиологии. 2007. Т. 47. № 2. С. 196–201.
Захаров А.Б., Бознак Э.И. Инвазийные виды в крупных речных системах Европейского Северо-Востока России // Проблемы изучения и охраны животного мира на Севере. Сыктывкар, 2009. С. 259–263.
Никольский Г.В. Список рыб из неолита бассейна р. Онеги // Бюл. МОИП. 1935. Вып. 3. С. 113–118.
Зуянова О.В. О результатах пробной интродукции судака в оз. Воже // Научн. тр. ГосНИОРХ. Л., 1989. Вып. 293. С. 80–83.
Студенов И.И., Новоселов А.П. О негативном экологическом эффекте при саморасселении судака Stizostedion lucioperca (Linnaeus, 1758) в бассейне р. Онеги // Чужеродные виды в Голарктике (Борок-2). Рыбинск-Борок, 2005. С. 174–175.
Павлов Д.С. Подходы к охране редких и исчезающих видов рыб // Вопр. ихтиологии. 1992. Т. 32. Вып. 5. С. 3–19.
Еловенко В.Н. Морфо-экологическая характеристика ротана Percottus glenii Dyb. в границах естественного ареала и за его пределами: Автореф. дис. … канд. биол. наук. М., 1985. 24 с.
Новоселов А.П., Фефилова Л.Ф., Еловенко В.Н. Биологические параметры и питание ротана Percottus glenii Dybowski, 1877, случайно вселенного в оз. Плесецкое (Архангельская область) // Чужеродные виды в Голарктике (Борок-2). Рыбинск-Борок, 2005. С. 159–160.
Новоселов А.П., Студенов И.И. О появлении каспийских видов белоглазки Abramis sapa (Pallas, 1814) и жереха Aspius aspius (Linnaeus, 1758) в бассейне р. Северной Двины // Вопр. ихтиологии. 2002. Т. 42. № 8. С. 615–622.
Новоселов А.П., Студенов И.И. О питании и пищевых взаимоотношениях саморасселившейся белоглазки Abramis sapa (Pallas, 1814) и аборигенного сига Coregonus lavaretus (Linnaeus, 1758) в бассейне р. Cеверной Двины // Естественные и инвазийные процессы формирования биоразнообразия водных и наземных экосистем. Ростов-на-Дону, 2007. С. 232–234.
Бознак Э.И. Ихтиофауна реки Вычегды (морфология, биология, зоогеография): Автореф. дис. … канд. биол. наук. Санкт-Петербург, 2003. 22 с.
Салахутдинов А.Н., Шакирова Ф.М. Возможные последствия от вселения чужеродных видов в Куйбышевское водохранилище // Чужеродные виды в Голарктике (Борок-2). Рыбинск-Борок, 2005. С. 26–27.
Дополнительные материалы отсутствуют.