

Экология, 2021, № 2, стр. 103-111
Изменение распространения широколиственных древесных видов в центральной части Южного Урала со второй половины ХХ в.
Н. И. Федоров a, *, В. Б. Мартыненко a, С. Н. Жигунова a, О. И. Михайленко b, Г. В. Шендель a, Л. Г. Наумова c
a Уфимский институт биологии Уфимского федерального исследовательского центра РАН
450054 Уфа, просп. Октября, 69, Россия
b Уфимский государственный нефтяной технический университет
450062 Уфа, ул. Космонавтов, 1, корп. 1, Россия
c Башкирский государственный педагогический университет им. М. Акмуллы
450008 Уфа, ул. Октябрьской революции, 3-а, Россия
* E-mail: fedorov@anrb.ru
Поступила в редакцию 30.05.2020
После доработки 10.07.2020
Принята к публикации 10.08.2020
Аннотация
С использованием тематических карт распространения широколиственных древесных видов Tilia cordata Mill., Quercus robur L., Acer platanoides L., Ulmus glabra Huds. в центральной части Южного Урала, созданных с использованием картографических материалов лесоустройств, проведенных в 70-е годы ХХ в. и начале XXI в., проанализировано изменение их распространения после прекращения экстремально холодных зим в конце 70-х годов ХХ в. Аномальные зимние температуры наиболее сильно повлияли на распространение A. platanoides и U. glabra. В настоящее время наблюдается увеличение распространения всех широколиственных пород и сдвиг границ их распространения на восток на левобережье р. Белой.
Как известно, Уральский хребет стал естественной физико-географической границей для распространения многих видов неморального комплекса, что во многом связано с континентальностью климата. Хребет является преградой на пути влажных и теплых атлантических воздушных масс. По этой причине на западном макросклоне и в его предгорьях климат более влажный и теплый, он более благоприятен для формирования широколиственных лесов и сопутствующих им вторичных лугов. На восточном макросклоне климат более континентальный, что обусловило господство гемибореальных светлохвойно-мелколиственных лесов западно-сибирского типа и степных сообществ. В центрально-возвышенной части Южного Урала (ЮУ) широко представлены темнохвойные бореальные и смешанные широколиственно-темнохвойные леса. Стык разных лесных биомов породил экотонный эффект регионального масштаба, который проявляется во взаимопроникновении в растительные сообщества видов трех флоро-ценотических комплексов – неморального, бореального и гемибореального, и повышении за счет этого видового богатства сообществ лесов [1‒5].
В направлении с запада на восток в Южно-Уральском регионе из состава насаждений исчезают Quercus robur L., Acer platanoides L., Ulmus glabra Huds. и Tilia cordata Mill. Далее на восток от Уральских гор на Кузнецком Алатау есть изолированный липовый остров [6]. Таким образом, на ЮУ проходит восточная граница основных видов лесообразователей неморальных широколиственных лесов европейской части России.
Одним из выдающихся специалистов в изучении широколиственных лесов ЮУ был П.Л. Горчаковский [1, 2, 7], труды которого актуальны и востребованы в настоящее время. Прошло 40 лет с опубликования его фундаментальной монографии “Растения европейских широколиственных лесов на восточном пределе их ареала” [7], в которой он обобщил результаты 20-летних исследований распространения T. cordata, Q. robur, A. platanoides и U. glabra на Урале. В дальнейшем эти данные для ЮУ были детализированы Г.В. Поповым [3], который создал тематические карты распространения древесных видов на основе материалов лесоустройств лесхозов Башкирии, проведенных в начале 70-х годов ХХ в.
Границы широколиственных лесов на ЮУ с 70-х годов ХХ в. не изучали, но на примере хвойных и мелколиственных пород было показано, что на распространение древесных видов на Урале оказывают сильное влияние не только лесохозяйственная деятельность, но и изменение климата [8], которое проявляется прежде всего в увеличении сомкнутости ранее разреженных лесных сообществ и “подъеме” березовых и хвойных лесов по склонам, что приводит к смещению границы лес‒тундра [9‒15].
Известно, что на распространение A. platanoides, Q. robur и U. glabra, обладающих пониженной зимостойкостью, существенно влияли повторяющиеся до конца 70-х годов ХХ в. аномальные зимние температуры. Среди холодных зим во второй половине ХХ в. выделялась зима 1968/69 г., когда в январе–феврале температура ниже –30°С держалась в течение 26 дней, а в течение 9 дней – ниже ‒40°С, а также зима 1978/79 г., когда температура доходила до –47…‒48 °С. На состояние деревьев также влияли весенние заморозки. Так, в мае 1969 г. в период начала распускания листьев температура дважды опускалась ниже –10°С. Отсутствие сильных морозов, начиная с 80-х годов ХХ в., создало более благоприятные условия для произрастания широколиственных видов.
Цель данной работы ‒ анализ изменения распространения широколиственных древесных пород на восточной границе их ареала в центральной части ЮУ, в том числе на территории геопарка “Торатау”, с 60‒70-х годов ХХ в.
МАТЕРИАЛ И МЕТОДЫ
Объект исследования ‒ широколиственные древесные породы (Tilia cordata Mill., Quercus roтbur L., Acer platanoides L., Ulmus glabra Huds.), произрастающие в предгорных и низкогорных широколиственных лесах западного макросклона ЮУ (территория геопарка “Торатау”) и заходящие в зону распространения сосново-березово-лиственничных лесов в среднегорной части ЮУ. Для анализа распространения этих пород в начале 70-х годов ХХ в. нами были оцифрованы тематические карты их распространения по Республике Башкортостан, созданные Г.В. Поповым [3] на основе таксационных описаний о присутствии и доминировании древесных видов в составе лесной растительности, а также данных лесников об изолированных находках широколиственных видов.
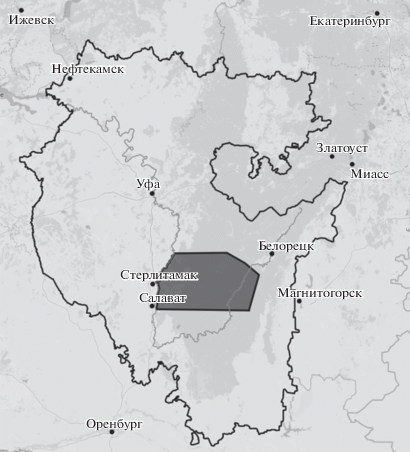
Для характеристики современного распространения использовали оцифрованные материалы лесоустройств Авзянского, Макаровского, большей части Гафурийского и, частично, Бурзянского лесхозов, проведенных в 2003‒2005 гг. Эта территория расположена между 53°15′ и 54°03′ с.ш. и 56°40′ и 58°00′ в.д. и занимает площадь около 9.5 тыс. км2 (рис. 1). Созданная ГИС-карта включает 59 540 полигонов, соответствующих выделам кварталов лесоустройств, для которых в атрибутивную таблицу была добавлена информация таксационных описаний о долевом участии древесных видов в древостое, их бонитете, возрасте и др. На основе ГИС-карты были созданы тематические карты современного распространения анализируемых видов. Информация на этих картах была дополнена данными о не отмеченных ранее местообитаниях широколиственных видов, выявленных в ходе экспедиционных работ в 2018‒2019 гг. Оцифровку растровых картографических материалов и создание тематических ГИС-карт проводили в программе QGIS 2.18.
РЕЗУЛЬТАТЫ
Тематические ГИС-карты распространения широколиственных древесных видов (T. cordata, А. platanoides, U. glabra, Q. robur) на западном макросклоне и в центральной части ЮУ в начале 70-х годов ХХ в. и начале ХХI в. приведены на рис. 2‒5. На основе их анализа установлено, что восточная граница массового распространения широколиственных лесов проходит по системе хребтов Зильмердак – Баштин – Большой Калу и в целом совпадает с областью массового распространения наиболее холодоустойчивого вида T. cordata.
Рис. 2.
Изменение распространения Tilia cordata Mill. с начала 70-х годов ХХ в.: а – распространение в начале 70-х годов ХХ в., б – в начале XXI в., дополненное данными 2014–2019 гг. (новые местообитания отмечены звездочками); черный цвет – доминирование, серый – присутствие в древостое, изолированные местообитания отмечены треугольниками; 1 – зоны появления вида в древесном ярусе.

Рис. 3.
Изменение распространения Quercus robur L. с начала 70-х годов ХХ в.: а – распространение в начале 70-х годов ХХ в., б – в начале XXI в., дополненное данными 2014–2019 гг. (новые местообитания отмечены звездочками); черный цвет – доминирование, серый – присутствие в древостое, изолированные местообитания отмечены треугольниками; 1 – зоны появления вида в древесном ярусе.
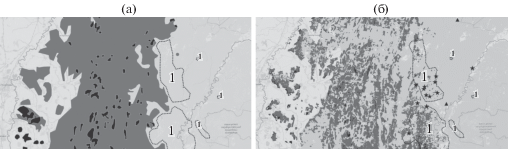
Рис. 4.
Изменение распространения Acer platanoides L. с начала 70-х годов ХХ в.: а – распространение в начале 70-х годов ХХ в., б – в начале XXI в., дополненное данными 2014–2019 гг. (новые местообитания отмечены звездочками); черный цвет – доминирование, серый – присутствие в древостое, изолированные местообитания отмечены треугольниками; 1 – зоны появления вида в древесном ярусе, 2 – зоны выпадения вида из древесного яруса.
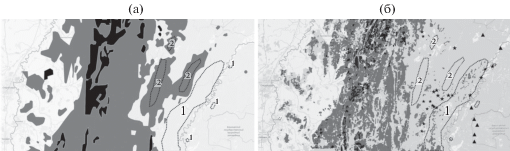
Рис. 5.
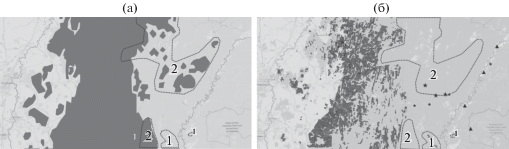
Изменение распространения Ulmus glabra Huds. с начала 70-х годов ХХ в.: а – распространение в начале 70-х годов ХХ в., б – в начале XXI в., дополненное данными 2014–2019 гг. (новые местообитания отмечены звездочками); черный цвет – доминирование, серый – присутствие в древостое, изолированные местообитания отмечены треугольниками; 1 – зоны появления вида в древесном ярусе, 2 – зоны выпадения вида из древесного яруса.
T. cordata ‒ наиболее распространенный древесный вид на западном макросклоне ЮУ, образующий как монодоминантные, так и смешанные с другими видами древостои. В центральной части ЮУ липа также наиболее распространена по сравнению с другими широколиственными видами. В целом восточная граница распространения T. cordata на тематической карте Г.В. Попова совпадает с данными П.Л. Горчаковского, однако последний указывал на наличие нескольких изолированных местообитаний (куртин) липы на левобережье р. Белой, которых нет на карте Г.В. Попова. Их отсутствие в начале 70-х годов ХХ в. может быть связано с проведением в этот период сплошных рубок. Сравнение современного распространения T. cordata с картографическими данными по распространению этого вида в начале 70-х годов ХХ в. показало, что в целом доминирование липы в древесном ярусе сохранилось на тех же территориях, но при этом произошел сдвиг границы распространения лесов с участием вида на восток, в том числе на левобережье р. Белой на 10–12 км (рис. 2). Продвижению липы способствуют ее успешное семенное размножение под пологом сосново-березовых лесов и выборочные рубки сосны и березы [16].
Другие широколиственные древесные породы менее распространены, чем T. cordata, и реже образуют чистые древостои. До начала интенсивной эксплуатации лесов вторым по значимости лесообразующим видом был Q. robur. В настоящее время в увалисто-холмистой полосе западного макросклона ЮУ леса с доминированием низкобонитетного дуба встречаются преимущественно на крутых инсолируемых склонах и вершинах увалов. Более высокобонитетные дубняки на относительно пологих склонах были вырублены и заместились преимущественно на липово-кленово-ильмовые леса, в которых дуб представлен в основном единичными крупными деревьями и подростом. В центральной части ЮУ он встречается на прогреваемых верхних частях склонов хребтов в составе сложных сосняков, в древостой которых могут входить также липа и осина. На распространение дуба существенное влияние оказали аномально низкие зимние температуры, продолжавшиеся до конца 70-х годов ХХ в. Ослабление крупных деревьев дуба в первом ярусе привело к их постепенному усыханию, и, согласно данным лесоустройств, в среднегорной части района исследования (Авзянский лесхоз) с 1989 г. по 2003 г. площади лесов с доминированием дуба снизились на 23%. В целом по Республике Башкортостан с 1966 г. по 2016 г. площади лесов с доминированием дуба и клена уменьшились почти в 3 раза [17].
Из рис. 3 видно, что усыхание дуба на западном макросклоне ЮУ и в его среднегорной части привело не только к уменьшению площади лесов с доминированием этого вида, но и его отсутствию в древостое в ряде мест, где в начале 70-х годов ХХ в. его участие было небольшим. Отсутствие с начала 1980-х годов сильных морозов привело к частичному восстановлению позиций вида и расширению его границ в южной части района исследований на восток на 10–15 км, в том числе за счет выхода в основной ярус сохранившегося крупного подроста. Причем при лесоустройстве в 2003 г. дуб был уже отмечен в составе древостоя на левобережье, а во время наших экспедиций в 2018–2019 гг. – на правобережье р. Белой, где этот вид не был обнаружен П.Л. Горчаковским и не отмечен на тематической карте Г.В. Попова.
Вид А. platanoides доминирует в смешанных широколиственных лесах чаще, чем дуб, особенно в восточной части горных широколиственных лесов западного макросклона ЮУ. При этом клен менее распространен в среднегорной части ЮУ по сравнению с дубом, что связано с его более ранним сезонным развитием и большим повреждением поздними весенними заморозками. Восточная граница распространения А. platanoides на тематической карте Г.В. Попова совпадает с данными П.Л. Горчаковского.
Из рис. 4 видно, что площади лесов с доминированием клена на западном макросклоне ЮУ, как и площади дубняков, сильно уменьшились. В отличие от дуба сокращение площадей лесов с доминированием клена после морозных зим в конце 70-х годов ХХ в. произошло преимущественно на восточной границе распространения вида. После прекращения экстремально холодных зим наблюдается увеличение участия клена в смешанных древостоях (что особенно выражено в перестойных сосново-березовых лесах), наиболее заметное на восточной границе распространения вида, где она сместилась на восток на 8–10 км. Кроме того, уже при лесоустройстве 2003 г. отмечались небольшие участки лесов с участием клена на левобережье р. Белой. Их наличие было подтверждено нами во время экспедиций 2018–2019 гг. Появление таких участков связано с увеличением плодоношения клена и его внедрением под полог разреженных перестойных сосново-березовых лесов, а также формированием подроста на вырубках с последующим выходом части подроста в древесный ярус, особенно при выборочной рубке хвойных пород [18].
На западном макросклоне и в центральной части ЮУ наименьшее распространение среди широколиственных древесных видов имеет U. glabra. Этот вид практически не образует чистые древостои и обычно встречается во втором и реже в первом ярусах смешанных широколиственных лесов. Граница распространения U. glabra на тематической карте Г.В. Попова совпадает с данными П.Л. Горчаковского. Из рис. 5 видно, что площади лесов с участием ильма на западном макросклоне ЮУ уменьшились, а восточная граница массового распространения этого вида в средней и северной частях анализируемой территории на момент лесоустройства 2003 г. сдвинулась на запад на 30–40 км, что стало следствием не только ослабления деревьев под влиянием морозных зим, но и вспышек численности насекомых-вредителей. Так, после холодной зимы в 1976 г. наблюдались вспышки насекомых-вредителей (заболонника большого ильмового Scolytus scolytus F. и заболонника струйчатого Scolytus multistriatus Мarsch.), обусловивших массовое распространение у ильма “голландской болезни” (графиоза), возбудителем которой является гриб Graphium ulmu Schwarz. В целом по сравнению с 60-ми годами ХХ в. площадь ильмовых лесов в Республике Башкортостан сократилась в 2 раза [19]. Тем не менее ильм встречается достаточно часто в подросте и даже выходит во второй ярус древостоя в ряде мест, где он отсутствовал в середине ХХ в., в том числе и на левобережье р. Белой.
Широколиственные виды присутствуют в сообществах смешанных сосново-березовых лесов, травяной ярус которых наряду с гемибореальными видами класса Brachypodio pinnati-Betuletea pendulae Ermakov, Koroljuk et Latchinsky 1991 содержит типичные виды неморального комплекса класса Carpino-Fagetea Jakucs ex Passarge 1968. При этом по флористическому составу нижних ярусов участки леса, на которых в древесном ярусе выявлены ранее не встречавшиеся широколиственные виды, не отличаются от участков экотонных смешанных лесов, в которых широколиственные виды присутствовали в основном древесном ярусе в 60–70-х годах ХХ в. Это свидетельствует о том, что расширение распространения широколиственных древесных видов в настоящее время происходит преимущественно на местах их прежнего произрастания.
ОБСУЖДЕНИЕ
Современная граница распространения широколиственных видов на ЮУ формировалась после окончания среднего суббореального теплового максимума [20], когда средняя годовая температура упала на 1–2.5°С [21]. При ксерофитизации и похолодании климата в центральной части ЮУ происходили постепенное выпадение дуба и сужение распространения ильма и клена до границ наиболее прогреваемых участков. Почти сплошной массив широколиственных и хвойно-широколиственных южно-уральских лесов распался на широколиственные леса западного макросклона и низкогорий ЮУ и единичные изолированные местообитания широколиственных пород восточнее границы основного распространения, которые находятся в окружении экотонных смешанных сообществ с доминированием в древесном ярусе светлохвойных и мелколиственных видов деревьев [22]. В зависимости от комплекса условий произрастания, и прежде всего температурного режима, в составе экотонных сообществ присутствуют липа, клен и ильм в подросте или в виде единичных деревьев во втором ярусе. В сохранении данных видов в этих местообитаниях при изменении климата ведущую роль играли вегетативное размножение и способность к произрастанию под пологом сосново-березовых лесов. Наиболее морозоустойчивая липа за счет способности к интенсивному возобновлению порослью от пня распространена более широко и сохраняет свое присутствие в подросте значительно восточнее других видов, в том числе на территории Башкирского государственного заповедника (см. рис. 2).
Рассматриваемый нами интервал времени, за который произошли описанные изменения распространения широколиственных древесных видов, условно делится на два периода. В первый из них, длившийся до конца 80-х годов ХХ в., были часты экстремально холодные зимы и весенние заморозки, что в сочетании со вспышками численности насекомых-вредителей и заболеваний деревьев ослабляло широколиственные древесные виды. Из-за экстремальных климатических явлений и вспышек численности непарного шелкопряда (Lymantria dispar (L.)) происходило усыхание значительной части деревьев дуба в широколиственных и смешанных лесах ЮУ [23, 24]. При этом снижение площадей дубовых лесов наблюдалось не только на ЮУ, но и в целом по Европе [25] и в Северной Америке [26]. Во второй половине XX в. было несколько вспышек распространения “голландской болезни” вязовых, в результате которых на ЮУ, как и во всем мире [27–29], произошла массовая гибель деревьев видов рода Ulmus, в том числе U. glabra.
Весенние заморозки во время распускания почек или формирования молодых неокрепших листьев и побегов также оказывают существенное влияние на состояние широколиственных деревьев, причем не только на дуб и ильм, но и на деревья ранне-вегетирующего клена остролистного [30–34]. От зимних морозов и весенних заморозков из рассматриваемых видов в наименьшей степени пострадала липа. Перечисленные факторы играли решающую роль в динамике лесов и взаимодействовали с более постепенными аутогенными изменениями их структуры и состава [35]. Как и в широколиственных лесах Европы [36, 37], при усыхании дуб и ильм постепенно замещались произрастающими с ними теневыносливыми видами – липой и кленом, а в смешанных лесах – осиной и березой [18].
Во второй период, продолжающийся в настоящее время, в связи с потеплением климата прекратились аномальные зимние морозы и снизилась частота весенних заморозков [8], что существенно улучшило условия произрастания широколиственных деревьев. Это оказало влияние не только на структуру древесного яруса широколиственных лесов, но и на их распространение на восточной границе ареала, где в настоящее время наблюдается увеличение плодоношения у липы, клена и ильма, а также их выход из подроста в древесный ярус. Данные изменения совпадают с мировыми тенденциями расширения ареалов широколиственных видов [38, 39].
Одним из факторов, определяющих скорость продвижения древесных видов, является интенсивность семенного размножения и эффективность распространения семян. На открытых местообитаниях основная масса семян T. cordata может переноситься на расстояние до 80 м, а видов A. platanoides и U. glabra – до 200 м [40]. Семена ильма могут также переноситься водой [41]. Основной перенос семян этих видов происходит из наиболее прогреваемых участков в верхних частях склонов в их нижние части, где они образуют мелкий подрост, выходящий при благоприятных условиях в крупный подрост и второй древесный ярус. Желуди дуба распространяются птицами и млекопитающими и в Европе легко внедряются под полог монокультур сосны [42], однако в центральной части ЮУ для массовой инвазии этого вида недостаточно плодоносящих деревьев. В целом наиболее эффективно семенное размножение реализуется у клена остролистного.
Наблюдаемое увеличение площадей лесов с участием липы – одного из основных лесообразователей среднеголоценовых лесов Европы [43] и ЮУ, в настоящее время происходит за счет выхода крупного подроста в основной древесный ярус в экотонных сообществах. Этому значительно способствуют и выборочные рубки сосны.
Вследствие интенсивного семенного размножения клена его теневыносливый подрост легко внедряется под полог лесов с доминированием других пород. Мы наблюдали обильный подрост этого вида в Белорецком районе (в березняках в окрестностях дер. Тукан и на вырубках сосняков северо-восточнее пос. Авзян), а также южнее Башкирского госзаповедника под пологом нарушенных выпасом перестойных липняков. Пополнение древесного яруса этим видом происходит так же, как и у Acer rubrum L. в лесах Европы, за счет появления одиночных деревьев в окнах полога древостоя и выхода в древесный ярус пережившего период подавления подроста при рубках леса [44].
Ильм, наиболее пострадавший не только от морозов, но и насекомых-вредителей, на ЮУ восстанавливается медленнее липы и клена, хотя и достаточно распространен в крупном подросте, который часто выходит во второй ярус. Дуб восстанавливается еще медленнее, что в значительной степени связано с ослабленным плодоношением этого вида. Можно ожидать постепенное восстановление его позиций на наиболее прогреваемых ксерофитных участках. В мезофитных сообществах его восстановление будет происходить гораздо медленнее и только в комплексе с сопутствующими видами (липы, ильма).
При сохранении современных тенденций изменения климата в центральной части ЮУ будет увеличиваться распространение широколиственных видов на восточной границе их ареалов, что соответствует данным о расширении подходящей среды обитания для широколиственных видов [45, 46]. Однако влияние потепления климата на широколиственные породы деревьев неоднозначно – оно может проявляться в изменении сроков сезонного развития от начала периода вегетации до полного развития [47]. Сдвиг на более ранние сроки начала вегетации увеличивает риск повреждения распускающихся листьев заморозками, что в наибольшей степени будет оказывать влияние на клен. Потепление климата также ускоряет темпы роста, плодовитость, зимовочную выживаемость и увеличение численности насекомых-вредителей [48, 49], что может провоцировать новые вспышки “голландской болезни” у ильма. Распространению широколиственных древесных видов могут способствовать также выборочная вырубка хвойных пород и бактериальная водянка березы (Erwinia multivora Sch.-Parf.), которая при потеплении климата может продвинуться в среднегорную часть ЮУ.
ЗАКЛЮЧЕНИЕ
Таким образом, можно констатировать, что восточные границы распространения широколиственных древесных видов в центральной части горно-лесной зоны ЮУ в течение XX в. сдвигались на запад после серии экстремально холодных зим, а в настоящее время в связи с потеплением климата постепенно смещаются на восток. На западном макросклоне ЮУ липа получила определенные преимущества по сравнению с дубом, кленом и ильмом из-за большей морозоустойчивости и способности к вегетативному размножению.
Изменение ареала древесных пород происходит более медленными темпами, чем изменение климата [50]. На ЮУ это в значительной степени связано с конкуренцией с березой повислой и достаточно долго растущей до возраста рубки сосной. О потенциале изменения границ распространения широколиственных древесных видов в значительной степени можно судить по удаленности от основного ареала изолированных местообитаний [51–53], среди которых есть обнаруженное М.И. Пряхиным местонахождение липы и ильма на восточном склоне хребта Крыкты (отроги Южно-Уральских гор в Зауралье) [7]. До момента смыкания границ массового распространения широколиственных древесных видов с их восточными изолированными местообитаниями можно говорить об изменениях растительности в пределах ее флюктуации в голоцене. Тем не менее происходящие изменения растительности обусловливают необходимость их мониторинга и коррекции в подходах к ведению лесного хозяйства.
Работа выполнена в рамках государственного задания Минобрнауки России № 075-00326-19-00 по теме № АААА-А18-118022190060-6.
Список литературы
Горчаковский П.Л. Основные проблемы исторической фитогеографии Урала // Тр. Ин-та экологии растений и животных Урал. фил. АН СССР. Свердловск, 1969. Вып. 66. 286 с.
Горчаковский П.Л. Широколиственные леса и их место в растительном покрове Южного Урала. М.: Наука, 1972. 146 с.
Попов Г.В. Леса Башкирии. Уфа: Башкир. кн. изд-во, 1980. 114 с.
Martynenko V.B., Mirkin B.M., Muldashev A.A. Syntaxonomy of Southern Urals forests as a basis for the system of their protection // Rus. J. of Ecology. 2008. V. 39. № 7. P. 459–465.
Миркин Б.М., Мартыненко В.Б., Широких П.С., Наумова Л.Г. Анализ факторов, определяющих видовое богатство сообществ лесов Южного Урала // Журн. общ. биол., 2010. Т. 71. № 2. С. 131–143.
Особо охраняемые природные территории России: современное состояние и перспективы развития / Авт.-состав.: В.Г. Кревер, М.С. Стишов, И.А. Онуфреня. М.: WWF России, 2009. 459 с.
Горчаковский П.Л. Растения европейских широколиственных лесов на восточном пределе их ареала. Свердловск, 1968. 207 с.
Оценочный доклад об изменениях климата и их последствиях на территории Российской Федерации [Электронный ресурс] - 2008. Режим доступа: http://climate2008.igce.ru/v2008/htm/index00.htm
Шиятов С.Г., Терентьев М.М., Фомин В.В. Пространственно-временная динамика лесотундровых сообществ на Полярном Урале // Экология. 2005. № 2. С. 83–90.
Шиятов С.Г. Динамика древесной и кустарниковой растительности в горах Полярного Урала под влиянием современных изменений климата. Екатеринбург: УРО РАН, 2009. 216 с.
Елсаков В.В., Марущак И.О. Спектрозональные спутниковые изображения в выявлении трендов климатических изменений лесных фитоценозов западных склонов Приполярного Урала // Компьютерная оптика. 2011. Т. 35. № 2. С. 281–286.
Им С.Т., Харук В.И. Климатически индуцированные изменения в экотоне альпийской лесотундры плато Путорана // Исследование Земли из космоса. 2013. № 5. С. 32.
Елсаков В.В., Щанов В.М., Бирюкова В.С. и др. Изменения предгорных лесов бассейна р. Щугор по материалам спутниковой съемки периода 1986–2016 гг. // Лесные экосистемы в условиях изменения климата: биологическая продуктивность и дистанционный мониторинг. Йошкар-Ола, 2016. С. 51–57.
Моисеев П.А., Шиятов С.Г., Григорьев А.А. Климатогенная динамика древесной растительности на верхнем пределе ее распространения на хребте Большой Таганай за последнее столетие. Екатеринбург: УрО РАН, 2016. 136 с.
Моисеев П.А., Гайсин И.К., Бубнов М.О., Моисеева О.О. Динамика древесной растительности на участках остепненных склонов Южного Крака в последние 80 лет // Экология. 2018. № 2. С. 157–162. https://doi.org/10.7868/S0367059718020105
Давыдычев А.Н., Горичев Ю.П., Кулагин А.Ю., Сулейманов Р.Р. Возобновительные процессы под пологом широколиственно-темнохвойных лесов Южного Урала // Лесоведение. 2011. № 2. С. 51–61.
Лесной план Республики Башкортостан [Электронный ресурс] – 2012. Режим доступа: http://docs.cntd.ru/document/935110326. (дата обращения: 30.06.2020).
Жигунова С.Н., Федоров Н.И., Михайленко О.И. Восстановительные сукцессии на сплошных вырубках сосново-березовых лесов центральной части Южного Урала // Научные ведомости Белгородского гос. ун-та. Серия: Естественные науки. 2013. № 3 (146). С. 30–35.
Гайнанов С.Г. Сохранение и воспроизводство ильмовых на Южном Урале // Вестник Башкирского гос. аграрного ун-та. 2013. № 4. С. 114–117.
Хотинский Н.А. Радиоуглеродная хронология и корреляция природных и антропогенных рубежей голоцена // Новые данные по геохронологии четвертичного периода / Ред. Пуннинг Я.-М.К., Иванова И.К., Кинд Н.В., Чичагова О.А.. М.: Наука, 1987. С. 39–45.
Novenko E., Tsyganov A., Volkova E. et al. The Holocene palaeoenvironmental history of Central European Russia reconstructed from pollen, plant macrofossil and testate amoeba analyses of the Klukva peatland // Tula region. Quat. Res. 2015. V. 83. P. 459–468.
Мартыненко В.Б., Широких П.С., Миркин Б.М. Синтаксономия коренных лесов Южно-Уральского региона и их горных аналогов // Разнообразие и динамика лесных экосистем России. В 2-х кн. М., 2013. С. 67–129.
Сергеев Л.И. Зимостойкость лесообразующих пород и лесовозобновление в Башкирии // Охрана, рациональное использование и воспроизводство лесных ресурсов Башкирии: Tез. докл. науч.-практ. конф. Уфа, 1974. С. 30–34.
Кучеров С.Е., Кучерова С.В. Усыхание дуба на юго-восточной границе ареала в связи с воздействием повреждающих факторов // Известия Самарского научного центра РАН. 2013. Т. 15. № 3-4. С. 1341–1343.
Gagen M., Matthews N., Denman S. et al. The tree ring growth histories of UK native oaks as a tool for investigating Chronic Oak Decline: An example from the Forest of Dean // Dendrochronologia. 2019. V. 55. P. 50–59.
Ma W., Liang J., Cumming J.R. et al. Fundamental shifts of central hardwood forests under climate change // Ecol. Model. 2016. V. 332. P. 28–41.
Tomlinson I, Potter C. “Too little, too late”? Science, policy and Dutch Elm Disease in the UK // J. of Historical Geography. 2010. V. 36. P. 121–131. https://doi.org/10.1016/j.jhg.2009.07.003
Fisher M.C., Henk D.A., Briggs C.J. et al. Emerging fungal threats to animal, plant and ecosystem health // Nature. 2012. V. 48. P. 186–194.
Nitschke C.R., Nichols S., Allen K. et al. The influence of climate and drought on urban tree growth in Southeast Australia and the implications for future growth under climate change // Landsc. Urban Plan. 2017. V. 167. P. 275–287. https://doi.org/10.1016/j.landurbplan.2017.06.012
Cannell M.G.R. Spring frost damage on young Picea sitchensis. 1. Occurence of damaging frosts in Scotland compared with western North America // Forestry. 1984. V. 57. № 2. P. 159–175.
Rammig A., Jönsson A.M., Hickler T. et al. Impacts of changing frost regimes on Swedish forests: Incorporating cold hardiness in a regional ecosystem model // Ecological Modelling, 2010. V. 221. P. 303–313. https://doi.org/10.1016/j.ecolmodel.2009.05.014
Augspurger C.K. Reconstructing patterns of temperature, phenology, and frost damage over 124 years: spring damage risk is increasing // Ecology. 2013. V. 94. P. 41–50. https://doi.org/10.1890/12-0200.1
Muffler L., Beierkuhnlein C., Aas G. et al. Distribution ranges and spring phenology explain late frost sensitivity in 170 woody plants from the Northern Hemisphere // Global Ecol. Biogeogr. 2016. V. 25. P. 1061–1071. https://doi.org/10.1111/geb.12466
Bascietto M., Bajocco S., Mazzenga F., Matteuccid G. Assessing spring frost effects on beech forests in Central Apennines from remotely-sensed data // Agricultural and Forest Meteorology. 2018. V. 248. P. 240–250.
Von Oheimb G., Brunet J. Dalby Söderskog revisited: long-term vegetation changes in a south Swedish deciduous forest // Acta Oecol. 2007. V. 31. P. 229–242. https://doi.org/10.1016/j.actao.2006.12.001
Abrams M.K., Orwig D.A., Dockry M.K. Dendroecology and successional status of two contrasting old-growth oak forests in the Blue Ridge Mountains, U.S.A. // Can. J. Forest Res. 1997. V. 27. P. 994–1002.
Reif A., Gärtner S.M. Die natürliche Verjüngung der laubabwerfenden Eichen – arten Stieleiche (Quercus robur L.) und Traubeneiche (Quercus petraea Liebl.) – eine Literaturstudie mit besonderer Berücksichtigung der Waldweide // Waldokologie Online. 2007. Bd 5. S. 79–116.
Putnam R.C., Reich P.B. Climate and competition affect growth and survival of transplanted sugar maple seedlings along a 1700 km gradient // Ecol. Monogr. 2017. V. 87. P. 130–157.
Gao W.-Q., Liu J.-F., Xue Z.-M. et al. Geographical patterns and drivers of growth dynamics of Quercus variabilis // Forest Ecology and Management. 2018. V. 429. P. 256–266.
Евстигнеев О.И., Мурашев И.А., Коротков В.Н. Анемохория и дальность рассеивания семян деревьев Восточно-Европейских лесов // Лесоведение. 2017. №1. С. 45–52.
Venturas M., Nanos N., Gil L. The reproductive ecology o: Ulmus laevis Pallas in a transformed habitat // Forest Ecology and Management. 2014. V. 312. P. 170–178.
Woziwoda B., Krzyżanowska A., Dyderski M.K. et al. Propagule pressure, presence of roads, and microsite variability influence dispersal of introduced Quercus rubra in temperate Pinus sylvestris forest // Forest Ecology and Management. 2018. V. 428. P. 35–45. https://doi.org/10.1016/j.foreco.2018.06.033
Grant M.J., Waller M.P., Groves J.A. The Tiliadecline: vegetation change in lowland Britain during the mid and late Holocene // Quat. Sci. Rev. 2011. V. 30. P. 394–408.
Hart J.L., Buchanan M.L., Clark S.L., Torreano S.J. Canopy accession strategiesand climate-growth relationships in Acer rubrum // Forest Ecology and Management. 2012. V. 282. P. 124–132. https://doi.org/10.1016/j.foreco.2012.06.033
Корзухин М.Д., Цельникер Ю.Л. Модельный анализ современных ареалов лесных древесных пород на территории России и их вариаций при возможных изменениях климата // Проблемы экологического мониторинга и моделирования экосистем. 2010. Т. 23. С. 249–268.
Rustad L., Campbell J., Dukes J.S. et al. Changing Climate, Changing Forests: The Impacts of Climate Change on Forests of the Northeastern United States and Eastern Canada. Newtown Square, PA: U.S. Department of Agriculture // Forest Service, Northern Research Station, 2012. 48 p.
Park H., Jeong Su-J., Ho Ch.-H. et al. Nonlinear response of vegetation green-up to local temperature variations in temperate and boreal forests in the Northern Hemisphere // Remote Sens. Environ. 2015. V. 165. P. 100–108.
Wang X.-Y., Zhao C.-Y., Jia Q.-Y. Impacts of climate change on forest ecosystems in Northeast Chin //Advances in Climate Change Research. 2013. V. 4. P. 230–241. https://doi.org/10.3724/SP.J.1248.2013.230
Butenschoen O., Scheu S. Climate change triggers effects of fungal pathogens and insect herbivores on litter decomposition // Acta Oecologica. 2014. V. 60. P. 49–56.
Brecka Aaron F.J., Shahi C., Chen Han Y.H. Climate change impacts on boreal forest timber supply // Forest Policy and Economics. 2018. V. 92. P. 11–21. https://doi.org/10.1016/j.forpol.2018.03.010
Keppel G., Van Niel K.P., Wardell-Johnson G.W. et al. Refugia: identifying and understanding safe havens for biodiversity under climate change // Global Ecol. Biogeogr. 2012. V. 21. P. 393–404.
Шевченко Н.Е., Смирнова О.В. Рефугиумы флористического разнообразия темнохвойных лесов Северного Урала как маркеры природной растительности восточно-европейской тайги // Экология. 2017. № 3. С. 171–177.
Kołaczek P., Gałka M., Apolinarska K. et al. A multi-proxy view of exceptionally early postglacial development of riparian woodlands with Ulmus in the Dniester River valley, western Ukraine // Rev. of Palaeobotany and Palynology. 2018. V. 250. P. 27–43.
Дополнительные материалы отсутствуют.