

Экология, 2021, № 2, стр. 143-152
Популяционная структура соболя Байкальской горной страны: анализ генетических и фенотипических характеристик
В. Г. Монахов a, *, М. Н. Ранюк a, М. В. Модоров a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: mon@ipae.uran.ru
Поступила в редакцию 04.04.2020
После доработки 23.07.2020
Принята к публикации 10.08.2020
Аннотация
Изучена генетическая и фенотипическая (по окраске меха) структура 14 выборок соболя Байкальской горной страны (БГС). Проанализировано 257 особей по 11 микросателлитным локусам и более 12 тыс. шкурок вида по 7 стандартным категориям окраски. Популяционная структура соболя региона неоднородна как в фенотипическом, так и в генетическом аспектах. В Забайкальском крае, Улетовском, Петровск-Забайкальском и Красночикойском районах обитает изолированная популяционная группировка соболя с темным мехом. Хребет Хамар-Дабан на юге оз. Байкал населяет популяция соболя, имеющая высокое генетическое разнообразие и самую светлую в изученном регионе окраску, что скорее всего обусловлено ее контактами с восточно-саянской популяцией. Остальные выборки БГС демонстрируют относительно высокое генетическое разнообразие и промежуточные параметры окраски мехового покрова, что может быть результатом массовых внутрирегиональных транслокаций и миграций. Результаты сравнительных тестов позволяют предположить, по крайней мере для семи выборок, опосредованные связи между фенотипической и генетической структурой популяций соболя в изученном регионе.
Соболь – важнейший ресурс таежных лесов Байкальского региона. В настоящее время он стал основным пушным видом не только Прибайкалья, но и всей страны. Шкурки зверька пользуются устойчивым спросом на внешнем рынке. Основная доля из примерно полумиллиона ежегодно поставляемых на экспорт шкурок соболя отнесена к баргузинскому, одному из самых ценных в пушной торговле, кряжу. При определении качества шкурки основное внимание уделяется качеству меха и особенно – окраске. Окраска меха у млекопитающих также и важная черта фенооблика вида, имеющая адаптивный характер. Выявление полиморфизма окраски и закономерностей его динамики в популяциях важно не только для науки, но и для практики. Такие вопросы, как рациональное использование ресурсов вида, их мониторинг, планирование мероприятий по охране, классификация и стандартизация пушной продукции и мн. др., трудно решать без знания закономерностей внутривидовой изменчивости, в частности окраски меха.
Около века назад численность зверька находилась на катастрофическом уровне из-за перепромысла, возникла угроза потери ценнейшего вида. И в те годы на данную проблему было обращено пристальное внимание правительства страны. Именно в Прибайкалье в 1916 г. был учрежден первый заповедник России, основной целью которого стала организация охраны соболя [1], а также положено начало созданию целой сети ООПТ (семь заповедников, четыре национальных парка, многочисленные заказники и памятники природы). Кроме того, для ускорения заселения пустующих участков ареала решено было применить интродукцию. Племенной материал в основном (61.4% [2]) брали из байкальского региона – Иркутской области и Бурятии: около 12 тыс. зверьков. Результаты реинтродукции оказались успешными, современный ареал соболя был восстановлен практически до его исторических границ, чему во многом способствовали вновь образованные реакклиматизированные популяции потомков интродуцентов из Байкальского региона [2]. Возможно, что одним из факторов успешной реинтродукции байкальского соболя в районы Западной и Восточной Сибири стало разнообразие населения соболя на территории Байкальской горной страны и соответственно генетическое разнообразие выпущенных животных. И хотя генетических тестов при отлове животных для последующих выпусков не делали, однако обращали пристальное внимание на морфологические характеристики зверей, а именно на качество меха, самый важный в экономическом смысле признак [2].
На окраску меха соболя обращалось внимание в видовых систематических сводках начиная с самых первых в XVII–XVIII вв. Изменчивость окраски меха зверька изучали в основном в географическом аспекте, при обосновании пушного стандарта, в таксономических публикациях и при оценке результатов реинтродукции [3–10]. Для соболя Прибайкалья разными авторами были описаны следующие подвиды (вариететы): Mustela zibellina var. baicalensis Dybowski, 1922; Martes z. princeps Birula, 1922; M. z. vitimensis Timofeev and Nadeev, 1955; M. z. obscura Timofeev and Nadeev, 1955 [11], некоторые из которых считаются синонимичными [12]. Последний автор подтверждает только M. z. princeps – баргузинский соболь (для Баргузинского хр. и Забайкалья) и M. z. obscura – чикойский соболь (для Красночикойского, Улетовского, Хилокского, Кыринского и части Петровск-Забайкальского р-нов Читинской области [1, c. 247], ныне Забайкальского края).
Сравнительно недавно получили распространение исследования генетических основ изменчивости соболя [13–16], однако они до сих пор немногочисленны, а Байкальский регион в этом аспекте специально вообще не освещался. Сейчас появляется возможность сочетать классические зоологические исследования с генетическими тестами. Мы решили использовать такую возможность, проведя изучение нескольких прибайкальских популяций соболя по окраске мехового покрова, впервые получив их генетическую характеристику с помощью маркеров ядерной ДНК. При этом авторы исходили из предположения возможного несовпадения структурных классификаций, полученных двумя методами (нулевая гипотеза).
Соответственно основной целью данного исследования ставился анализ популяционной структуры населения соболя Байкальской горной страны. Для достижения цели были поставлены следующие задачи: 1) изучить популяционную генетическую структуру соболя на основе изменчивости микросателлитных локусов яДНК; 2) выявить популяционную фенотипическую структуру на основе изменчивости окраски меха соболя Байкальской горной страны, которого принято относить [1–4, 9] к одному из наиболее ценных по меховым качествам.
МАТЕРИАЛ И МЕТОДИКА
Район исследования
По определению [17] Байкальская горная страна (БГС) на западе граничит с Восточным Саяном, на юге – со степями Монголии, на востоке ее граница проходит по междуречью рек Олёкма и Зея, с севера она прилегает к Среднесибирскому плоскогорью и Приленскому плато.
В анализ популяционной структуры соболя БГС были включены особи из 14 районов Иркутской и Амурской областей, Республики Бурятия и Забайкальского края (рис. 1 ). Взяты выборки из следующих районов: Кабанский (хр. Хамар-Дабан), Северо-Байкальский, Баунтовский и Баргузинский – Республика Бурятия, Красночикойский, Петровск-Забайкальский, Чернышевский, Улетовский, Тунгокоченский и Тунгиро-Олёкминский – Забайкальский край. Иркутская область представлена Ольхонским, Казачинско-Ленским районами и нижним бассейном р. Витим (в пределах Бодайбинского и Мамско-Чуйского р-нов). Восточную окраину БГС представляет выборка из бассейна р. Нюкжа (приток р. Олёкма).
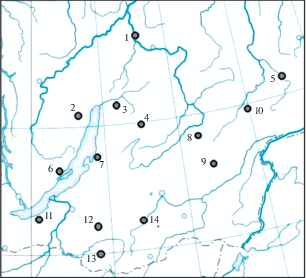
Генетический анализ
Для исследования генетической структуры населения соболя Байкальской горной страны в анализ были включены данные по 257 особям из 14 географических выборок (табл. 1, рис. 1 ). Образцы тканей предоставлены охотниками, добывавшими соболя на легальной основе. ДНК выделяли из образцов кожи и мышц при помощи наборов для выделения ДНК из тканей животных DNA-extran-2 KIT (Синтол, Россия). Все особи были генотипированы по 11 микросателлитным локусам, разработанным ранее для соболя (Mzf51, Mzf56; [18]), американской куницы Martes americana, росомахи Gulo gulo, американского барсука Taxidea taxus (Ma1, Ma3, Ma8, Ma15, Ma19; [19]), каменной куницы Martes foina (Mf8.7, Mf8.8; [20]), американской норки Neovison vison и горностая Mustela erminea (Mvis72, Mvi2243; [21, 22]).
Таблица 1.
Результаты анализа 14 выборок соболя БГС на основании аллельного состава 11 микросателлитных локусов яДНК
| Выборка (район) | Обозначение | Регион | n | Na | Ne | Ho | He |
|---|---|---|---|---|---|---|---|
| 1. Река Витим (низовья) | ВИТ | Иркутская обл. | 16 | 6.18 | 4.29 | 0.61 | 0.72 |
| 2. Казачинско-Ленский | КЗЛ | Иркутская обл. | 17 | 5.91 | 3.70 | 0.68 | 0.69 |
| 3. Северо-Байкальский | СБА | Бурятия | 25 | 6.91 | 4.09 | 0.69 | 0.70 |
| 4. Баунтовский | БАУ | Бурятия | 5 | 4.55 | 3.71 | 0.73 | 0.63 |
| 5. Река Нюкжа | НЮК | Амурская обл. | 21 | 6.64 | 3.68 | 0.65 | 0.70 |
| 6. Ольхонский | ОЛХ | Иркутская обл. | 7 | 5.09 | 3.30 | 0.61 | 0.67 |
| 7. Баргузинский | БАР | Бурятия | 14 | 6.09 | 3.73 | 0.62 | 0.69 |
| 8. Тунгокоченский | ТКЧ | Забайкальский край | 14 | 6.18 | 3.88 | 0.64 | 0.69 |
| 9. Чернышевский | ЧЕР | Забайкальский край | 7 | 5.18 | 3.71 | 0.62 | 0.69 |
| 10. Тунгиро-Олёкминский | ТОЛ | Забайкальский край | 11 | 6.55 | 4.46 | 0.70 | 0.70 |
| 11. Хр. Хамар-Дабан | ХМН | Бурятия | 54 | 7.45 | 4.42 | 0.67 | 0.71 |
| 12. Петровск-Забайкальский | ПЗА | Забайкальский край | 25 | 5.09 | 3.15 | 0.58 | 0.60 |
| 13. Красночикойский | ЧИК | Забайкальский край | 18 | 4.82 | 2.74 | 0.54 | 0.58 |
| 14. Улетовский | УЛЕ | Забайкальский край | 23 | 4.36 | 2.58 | 0.46 | 0.54 |
Амплификацию выполняли в общем объеме 10 мкл с добавлением 10× TaqBuffer с (NH4)SO4 (Thermo Scientific), Taq ДНК полимеразой (SibEnzyme, Россия), 5 мМ MgCl2, 0.2 мМ dNTP, 8.6 мМ (1%) ДМСО и 20–200 нг геномной ДНК. Концентрация праймера колебалась от 0.4 до 11 мкл, ПЦР проводили с использованием T100 Thermal Cycler (Bio Rad).
Амплификацию проводили при 94°C в течение 3 мин, затем 4 цикла денатурации при 98°C в течение 30 с, отжиг при 59°C в течение 120 с, 72°C в течение 90 с, следующие 20 циклов проводили денатурацию при 90°C в течение 30 с, отжиг при 59°C в течение 120 с, затем 72°C в течение 75 с и при окончании 68°C в течение 30 мин. Скорость нагрева от 59 до 72°С составляла не более 0.3°С/с.
На следующем этапе продукты ПЦР смешивали с формамидом и размерным стандартом S-550 Lyz (Гордиз, РФ) в соответствии с рекомендациями производителя. После 5-минутной денатурации с последующим быстрым охлаждением ПЦР-продукты определяли с использованием генетического анализатора ABI 3130 (AppliedBiosystems), а длину фрагмента оценивали с помощью программного обеспечения GeneMapper v. 3.7.
Для расчета показателей аллельного разнообразия и гетерозиготности, генетических дистанций FSt и анализа матрицы генетических дистанций методом главных координат использовали GenALEx 6.5 [23, 24]. Для оценки генетической структуры применяли байесовскую кластеризацию, реализованную в программе STRUCTURE 2.3.4 [25], для оценки наиболее вероятного числа кластеров использовали Structure Harvester [26].
Фенотипический анализ
Для перечисленных выше выборок получены данные об окраске меха соболя из двух источников. В монографии В.Д. Петренко [27] приведены результаты сортировки шкурок соболя по цветовым категориям за 1996–2003 гг., из которой для целей нашей работы привлечены сведения о 1122 экз., касающиеся выборок под номерами 1–3, 5–7, 9, 13 (см. табл. 2). Данные за 2016–2019 гг. (11 529 шкурок) получены в аукционной компании “Русский соболь” (г. Иркутск) из актов сортировки экспортных партий шкурок. Суммарный объем изученных по окраске меха выборок составил 12 651 экз. (см. табл. 2).
Таблица 2.
Цветовой состав изученных выборок соболя за 1996–2019 гг. (доли групп тeмных, средних и светлых шкурок, % ± SE)
| Выборка (район) | n | Темные | Средние | Светлые | Балл окраски |
|---|---|---|---|---|---|
| 1. Река Витим (низовья) | 3267 | 31.0 ± 0.81 | 53.3 ± 0.87 | 15.7 ± 0.64 | 3.77 ± 0.020 |
| 2. Казачинско-Ленский | 269 | 7.8 ± 1.64 | 51.3 ± 3.05 | 40.9 ± 3.00 | 2.87 ± 0.063 |
| 3. Северо-Байкальский | 971 | 17.2 ± 1.21 | 63.2 ± 1.55 | 19.6 ± 1.27 | 3.44 ± 0.033 |
| 4. Баунтовский | 640 | 30.9 ± 1.83 | 52.2 ± 1.97 | 16.9 ± 1.48 | 3.67 ± 0.044 |
| 5. Река Нюкжа | 889 | 23.2 ± 1.42 | 51.4 ± 1.68 | 25.4 ± 1.46 | 3.45 ± 0.040 |
| 6. Ольхонский | 525 | 12.0 ± 1.42 | 49.9 ± 2.18 | 38.1 ± 2.12 | 2.99 ± 0.049 |
| 7. Баргузинский | 82 | 11.0 ± 3.45 | 47.6 ± 5.52 | 41.5 ± 5.44 | 2.91 ± 0.113 |
| 8. Тунгокоченский | 2917 | 30.5 ± 0.85 | 57.5 ± 0.92 | 12.0 ± 0.60 | 3.81 ± 0.019 |
| 9. Чернышевский | 549 | 25.0 ± 1.85 | 63.9 ± 2.05 | 11.1 ± 1.34 | 3.69 ± 0.043 |
| 10. Тунгиро-Олёкминский | 121 | 28.9 ± 4.12 | 58.7 ± 4.48 | 12.4 ± 3.00 | 3.76 ± 0.098 |
| 11. Хр. Хамар-Дабан | 2155 | 7.7 ± 0.57 | 38.7 ± 1.05 | 53.6 ± 1.07 | 2.64 ± 0.023 |
| 12. Петровск-Забайкальский | 68 | 32.4 ± 5.67 | 52.9 ± 6.05 | 14.7 ± 4.29 | 3.74 ± 0.139 |
| 13. Красночикойский | 135 | 38.5 ± 4.19 | 56.3 ± 4.27 | 5.2 ± 1.91 | 4.13 ± 0.076 |
| 14. Улетовский | 63 | 23.8 ± 5.37 | 69.8 ± 5.78 | 6.3 ± 3.07 | 3.83 ± 0.109 |
| Итого | 12 651 | 23.7 ± 0.38 | 52.8 ± 0.44 | 23.6 ± 0.38 | 3.48 ± 0.019 |
Таблица 3.
Интродукции соболя в районах БГС в середине ХХ в.
| Локалитеты выборок | Регион | n (2630) |
Годы выпусков |
Происхождение племенного материала (районы) |
|---|---|---|---|---|
| Хамар-Дабан | Иркутская обл. (Слюдянский) | 86 | 1939–1952 | Бодайбинский, Баргузинский, Киренский |
| Ольхонский | Иркутская обл. | 87 | 1941–1949 | Бодайбинский, Киренский |
| Хамар-Дабан | Бурятия (Кабанский, Селенгинский) | 141 | 1940–1950 | Прибайкальский |
| Баунтовский | Бурятия | 223 | 1950–1954 | Северо-Байкальский, Баргузинский |
| Тунгокоченский | Забайкальский край | 584 | 1950–1958 | Кабанский, Бодайбинский, Северо-Байкальский |
| Тунгиро-Олёкминский | Забайкальский край | 905 | 1953–1958 | Верхнебуреинский (Хабаровский край) |
| Нюкжа (Джелтулакский, ныне Тындинский) | Амурская обл. | 604 | 1953–1958 | Верхнебуреинский, Зейско-Учурский (Хабаровский край) |
Классификацию шкурок по цвету проводили дипломированные специалисты-товароведы пушно-мехового сырья. В качестве методической базы использовали общесоюзный “Стандарт на шкурки соболя невыделанные” ОСТ НКЗаг-414, по которому все шкурки вида делятся на 7 цветовых категорий, подобно их выделению у американской куницы Martes americana [28]. Мы объединили исходные цветовые категории в три: к группе “тeмные” относили шкурки категорий “головки” (2 категории) и “подголовка высокая”, к “средним” ‒ “подголовка нормальная” и “воротовой темный”, “светлыми”считали “воротовые нормальные” и “меховые”.
Ранее было предложено [3] оценивать колорографическую структуру популяций соболя при помощи интегрального показателя – индекса (балла) окраски, который вычисляется по формуле средней взвешенной, когда самой светлой категории “меховые” присваивается балл 1, а самой тёмной “головке высокой” – балл 7. Этот показатель мы использовали для сравнения наших данных с литературными. Для каждой выборки были рассчитаны групповые средние значения, стандартные отклонения и стандартные ошибки. Все расчеты произведены в пакете Statistica 6 (Statsoft).
РЕЗУЛЬТАТЫ
Генетическая изменчивость
Генетические различия 14 выборок оценили при помощи разработанного нами мультиплекса из 11 микросателлитных локусов [29]. В большинстве случаев при сравнении частот генотипов наблюдаемое соотношение генотипов не отличалось от ожидаемого по Харди–Вайнбергу, выявленные отклонения не носили однонаправленного характера.
В исследованных выборках соболя (см. табл. 1) показатель среднего числа аллелей на локус (Na) колебался от 4.36 (Улетовский р-н) до 7.45 (Хамар-Дабан); самые низкие значения выявлены для животных Улетовского и Красночикойского районов. В остальных изученных районах БГС показатели аллельного разнообразия несколько выше при наибольших значениях в выборках Хамар-Дабана, Северобайкальского и Тунгиро-Олёкминского районов.
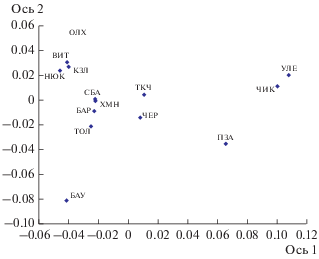
Значения генетических дистанций FSt для всех межгрупповых сравнений колебались от 0.016 до 0.124 (в среднем 0.051, см. Приложение, табл. S1 ). Наибольшее генетическое сходство проявили выборки Улетовского и Красночикойского р-нов (FSt = 0.016), а наибольшие различия (FSt более 0.09) с ними показали популяции Ольхонского, Казачинско-Ленского, Баунтовского р-нов, бассейнов Нюкжи, Витима. По результатам ординации матрицы FSt (табл. 2) методом главных координат вдоль оси 1, на которую приходится 40% объясняемой дисперсии, дифференцируются выборки Улетовского, Красночикойского и Петровск-Забайкальского р-нов (рис. 2). Вдоль оси 2 выделяется выборка Баунтовского района, но так как в анализ из этого района было включено всего 5 особей, большинство значений FSt для данной выборки недостоверны (см. Приложение, табл. S1 ).
Рис. 2.
Результаты анализа значений генетических дистанций FSt 14 выборок соболя методом главных координат. Доля объясняемой дисперсии для оси 1 – 40%, для оси 2 – 15%.
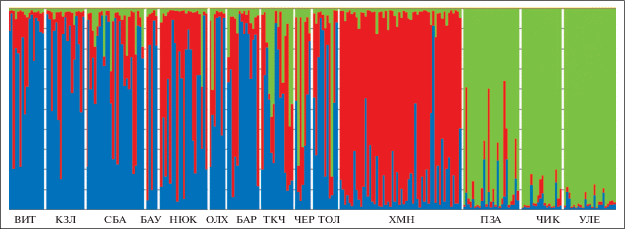
По результатам анализа генетической структуры населения соболя Байкальской горной страны байесовским методом кластеризации 14 исследуемых выборок формируют 3 основных кластера (рис. 3). Соболи, обитающие в южной части Байкала, на хребте Хамар-Дабан, по генетическим признакам отличаются от всех остальных особей (на рисунке они обозначены красным цветом). Выборки Красночикойского, Улетовского и Петровск-Забайкальского р-нов также формируют отдельный кластер (обозначены зеленым цветом). Остальные выборки образуют третий более разнородный кластер (обозначены преобладающим синим цветом).
Рис. 3.
Результаты кластеризации в программе STRUCTURE (K = 3) исследованных выборок соболя БГС. Одна вертикальная полоса соответствует одной особи (обозначения выборок см. в табл. 1).
Колорографическая структура
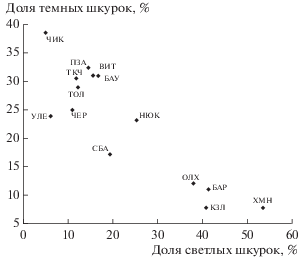
Анализ колорографической структуры изученных выборок также показал их неоднородность. Структура каждой из популяций по окраске меха приведена в табл. 2. Самую низкую (менее 5%) долю светлых шкурок (соответственно – наивысшую долю средних и темных категорий окраски) демонстрируют соболи Красночикойского, Улетовского, Чернышевского, Тунгокоченского, Тунгиро-Олёкминского и Петровск-Забайкальского районов. Эти группировки составляют ядро популяций соболя БГС с наиболее ценным мехом. Самая светлая окраска меха у зверьков из Ольхонского, Казачинско-Ленского, Баргузинского районов и Хамар-Дабана. Распределение исследованных выборок по окраске меха показано на рис. 4. Рассчитанный балл окраски колеблется от 2.64 (Хамар-Дабан) до 4.13 (Красночикойский район).
Рис. 4.
Классификация выборок соболя БГС на основе признаков окраски меха (обозначения выборок см. в табл. 1).
Попарное сравнение выборок по значению индекса окраски с вычислением критерия t-Стьюдента (см. Приложение, табл. S2 ) показало, что выборки Хамар-Дабана и Красночикойского р-на статистически высоко значимо (p < 0.001) отличаются от всех остальных в регионе. Две выборки (Нюкжа и Северо-Байкальск) проявляют сходство (p > 0.05) и образуют отдельную группу со средними параметрами окраски. Группу со светлой окраской меха образуют три выборки Ольхонского, Баргузинского и Казачинско-Ленского р-нов, которые сходны между собой (p > 0.05), но статистически отличны от других. Остальные выборки образуют пятую группу с темным мехом. В целом распределение по долям темных и светлых шкурок и интегральному параметру окраски меха получилось довольно сходным.
ОБСУЖДЕНИЕ
Исследований генетической структуры соболя по микросателлитным локусам не очень много. Известно несколько таких работ [18, 19, 30, 31], однако ни одна из них не была посвящена соболю Байкальской горной страны. В единственной работе [32] была предпринята попытка сравнения на основе мтДНК двух выборок Прибайкалья (11 экз.) и одной из Сихотэ-Алиня (10 экз.). Изученные особи разделились на две клады, в которых примерно поровну присутствовали как дальневосточные, так и прибайкальские экземпляры. Авторы не выявили значительных различий как между регионами, так и между двумя прибайкальскими выборками из соседних районов верхней Лены.
При генетическом анализе популяционной структуры соболя выявлено (см. рис. 2, 3 ), что на территории БГС обитает достаточно изолированная популяция, локализованная в Красночикойском, Улетовском и Петрово-Забайкальском районах (“чикойская” – по определению В.В. Тимофеева и В.Н. Надеева [1]). Для выборок из этих районов характерны самые низкие показатели генетического разнообразия: значения генетических дистанций (FSt) между ними и выборками из других районов БГС самые высокие из выявленных (см. табл. 1S, рис. 2).
Еще одна обособленная популяция соболя обитает в юго-западной части БГС, на хребте Хамар-Дабан. Зверьки из этого района демонстрируют генетическое своеобразие (см. рис. 3), однако в отличие от “чикойской” популяции обладают высокой генетической изменчивостью (см. табл. 1) и средними значениями FSt при парном сравнении с другими выборками (см. табл. 1S, рис. 2 ). Скорее всего, данная популяция не столько изолирована от соседних байкальских популяций соболя, сколько несет в себе генетический след мигрантов из соседних Саянских гор (с запада), что отличает ее от других популяций БГС, но в то же время повышает генетическое разнообразие внутри самой популяции. Не случайно Г.И. Монахов и В.В. Тимофеев [33] объединили Хамар-Дабанский очаг с соболем Восточного Саяна.
Выборки из остальных районов БГС демонстрируют похожие показатели как по генетическому разнообразию (см. рис. 3 ), так и по значениям генетических дистанций при парном сравнении (см. табл. S1 , рис. 2 ). Кроме миграционной активности соболя, это можно объяснить влиянием многочисленных межрайонных транслокаций соболя внутри БГС [2]. Эти факты относятся к 6 локалитетам из 14 изученных (табл. 3). В остальных изученных нами районах интродукционных работ не проводилось, за исключением Баргузинского, где в 1930 г. был произведен выпуск двух соболей, отловленных в Баргузинском заповеднике [2].
Общее количество переселенцев за период 1939–1958 гг. по данным [2] составило 2630 особей. Основная доля интродуцентов (1489, или 56.6%) пришлась на два района Забайкальского края и бассейн Нюкжи (23%), где расселены зверьки в основном из бассейна р. Бурея, а на остальные 4 района (табл. 3) пришлось 20.4%. Таким образом, проведенные в 1939–1958 гг. региональные интродукции связывают между собой половину из исследованных нами выборок (Витим, Хамар-Дабан, Ольхонский, Баунтовский, Северо-Байкальский, Баргузинский и Тунгокоченский р-ны). Несомненно, проведенные транслокации в пределах БГС способствовали перемешиванию генофонда в регионе, что и отражается на результатах наших тестов (см. рис. 2, 3 ).
Таким образом, по результатам анализа изменчивости аллельного состава 11 микросателлитных локусов яДНК нами выделены три популяционные группы: 1) выборки Улетовского, Петровск-Забайкальского и Красночикойского районов Забайкальского края, которые представляют собой одну популяцию, некоторым образом изолированную от соседних; 2) выборка Хамар-Дабана, которая также демонстрирует генетические отличия от остальных выборок соболей Прибайкалья и высокое генетическое разнообразие; 3) все остальные исследованные выборки, которые не показывают значительных различий между собой, что может быть результатом транслокаций и миграционной активности соболя даже в условиях достаточно сложного горного ландшафта (рис. 5а).
Рис. 5.
Распределение генетических признаков (а) и групп окраски меха (б) в 14 выборках M. zibellina БГС: а – доля генетических кластеров по результатам байесовской классификации по 11 микросателлитным локусам; б – доля особей с темной (черный цвет), средней (темно-серый цвет) и светлой (светло-серый цвет) окраской меха.
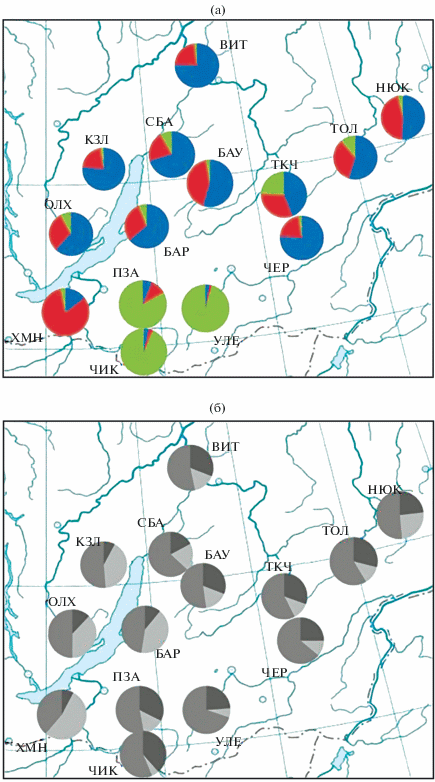
Изучению структуры байкальских популяций по окраске меха посвящен ряд работ, каждая из которых отличалась от предыдущей возрастающим количеством материала и числом выборок. Первые репрезентативные данные по окраске меха байкальских соболей опубликованы К.М. Еремеевой [3], которая привела данные о колорографической структуре соболя северной части Прибайкалья в 1946–1948 гг. и рассчитала для нее средний балл (3.9). Н.Н. Бакеев [5] привел цветовую характеристику трех популяций из БГС за 1951–1966 гг.: Витимской, Северо-Байкальской и Хамар-Дабанской, отметив для всех постепенное снижение среднего балла окраски (3.46–3.07; 3.57–3.07; 3.4–2.89 соответственно). Более подробно провел анализ окраски популяций Прибайкалья Г.И. Монахов [6]. Изучив данные о более чем 120.5 тыс. шкурок, он привел интегральную характеристику Хамар-Дабанской (3.08), Баргузинской (3.29) и суммарную для Витимской и Мамско-Чуйской (2.96) популяций. Наиболее подробно колорографическая структура соболя Прибайкалья описана В.Г. Монаховым [10] с привлечением данных по 238 тыс. шурок за 1950–1985 гг. Рассчитанный им средний балл окраски составил: Хамар-Дабан ‒ 3.08, Баргузин ‒ 3.17, Витим ‒ 3.18.
Для настоящего исследования нами привлечен материал, который позволяет дать колорографическую характеристику изучаемым группировкам соболя за последние 20 лет, соответствующие периоду сбора генетического материала (см. табл. 2, рис. 4 ) как в сравнении с собственными и литературными сведениями в ретроспективе, так и с полученными нами данными по генетической структуре на основе 11 микросателлитных локусов.
Анализ колорографической структуры популяций Прибайкалья на материале нового тысячелетия при сравнении с данными за 1950–1985 гг. [10] показал, что в некоторых из них произошли заметные изменения долей цветовых групп. Например, осветление меха произошло на Баргузинском хребте – доля светлых шкурок возросла на 13.5% (p = 0.05), а индекс окраски снизился на 0.26 (p = 0.05; см. табл. 2). Для выборки хр. Хамар-Дабан также характерно заметное снижение индекса окраски на 0.44 балла (p < 0.001) и рост доли светлых шкурок на 25.5% (p < 0.001). Вероятной причиной этому является возможность контактов с расположенной западнее популяцией Восточного Саяна, где обитает более светлый соболь.
Однако в нижнем бассейне Витима в этот же период произошло потемнение меха соболя – снижение доли светлых шкурок на 11% (p < 0.001) и рост индекса окраски на 0.6 балла (p < 0.001) ([34], табл. 2). Увеличение индекса окраски зафиксировано также для соболя Красночикойского р-на – 0.8 балла (p < 0.001) при росте доли темных шкурок на 30% (p < 0.001). Потемнение меха отмечается и для соболя бассейна р. Нюкжа, Тунгиро-Олёкминского, Улетовского, Петровск-Забайкальского, Тунгокоченского и Чернышевского районов.
Основной очаг соболей с темным меховым покровом (суммарный индекс окраски 3.93 балла) расположен вдоль хребтов Яблонового, Малханского, Даурского, Хэнтейского и Черского. Во всем ареале вида для этого (“чикойского”) очага характерна самая темная окраска меха. По нашим данным [10], наиболее близки к данной группировке лишь соболи из бассейна р. Бурея (индекс окраски 3.37). Высокое качество меха и темную окраску соболя Южно-Забайкальской области отметили В.В. Тимофеев и В.Н. Надеев [1] и по этой причине выделили его в отдельный подвид M. z. obscura – чикойский соболь [1, с. 47]. Эти авторы обнаружили, что данный очаг со всех сторон изолирован степными и лесостепными участками [1, с. 246], которые препятствуют контактам с соседними популяциями.
Несколько светлее мех у соболя из популяций Нижнего Витима, Баунтовского, Чернышевского, Тунгокоченского и Тунгиро-Олёкминского районов (совокупный индекс окраски 3.77). Северобайкальские и зверьки из бассейна р. Нюкжа имеют балл окраски 3.44–3.45, а в остальных популяциях доля светлых зверьков составляет 38–53% при среднем индексе окраски ниже 3.0 балла (см. табл. 2).
Таким образом, по результатам анализа изменчивости окраски меха соболя в пределах БГС можно сделать вывод о преобладающей доле особей со средней окраской меха во всех изученных выборках вида (52.8%, табл. 2). Отличия между исследованными выборками проявляются в доле светлых и темных особей. Так, в отдельную группу можно выделить выборки Хамар-Дабан, Казачинско-Ленский, Баргузинский и Ольхонский районы с наименьшим баллом окраски и самой высокой долей светлых особей среди исследованных (41–53%). Остальные выборки характеризуются значительно меньшей долей светлых и большей долей темных особей, среди которых выделяется выборка Красночикойского района, в которой самая высокая доля особей с темной окраской меха (39%, табл. 2, рис. 5 б).
Полученная нами морфологическая структура популяций не в полной мере отражает генетическую структуру. Тем не менее выборки, проявляющие самые высокие морфологические различия – Хамар-Дабанская и три другие со светлым мехом (Ольхонский, Баргузинский, Казачинско-Ленский р-ны), а также Красночикойская вместе с Улетовской и Петровск-Забайкальской попадают в разные генетические кластеры (см. рис. 2 ‒5). Кроме того, обнаружена значимая связь между индексом окраски и собственным значением 1-й оси анализа генетических дистанций FSt методом главных координат (r = 0.558, F1,12 = 5.5, p = 0.038, β = 0.066).
Н.Н. Бакеев [35] выделял в данном регионе витимскую и баргузинскую географические популяции. По нашим данным, выборки из данных районов действительно заметно различаются по окраске, однако генетические данные не подтверждают изолированность данных популяций. Г.И. Монахов и В.В. Тимофеев [33] разделяли исследуемую территорию на четыре природных района: Патомское и Северо-Байкальское нагорья, Северо-Западное и Северо-Восточное Прибайкалье, Восточный Саян и Хамар-Дабан и Южное Забайкалье. Географически такой вывод основан на наличии разделяющих эти территории остепненных участков, долин рек и межгорных впадин [33, с. 85, рис. 20 ] , что предполагает выделение как минимум трех географически изолированных природных районов. Результаты настоящей работы подтверждают наличие отдельных популяций только на хр. Хамар-Дабан и в Южном Забайкалье (выборки Красночикойского, Улетовского и Петровск-Забайкальского районов). Выборки из Северо-Западного и Северо-Восточного Прибайкалья по нашим данным демонстрируют различия в окраске меха (на северо-западе выше доля светлых особей и, соответственно, ниже доля темных), однако между выборками из этих районов существуют генетические различия (см. рис. 5).
Результаты проведенных сравнительных тестов позволяют предположить, по крайней мере для семи из 14 изученных выборок, наличие опосредованных связей между фенотипической и генетической структурами популяций соболя в изученном регионе, т.е. нами получено свидетельство в пользу сходства структуры соболя БГС по микросателлитным маркерам и составу группировок на основе долей окрасочных морф, что опровергает выдвинутую нулевую гипотезу.
Таким образом, популяционная структура соболя Байкальской горной страны неоднородна как в фенетическом, так и генетическом аспектах. В Забайкальском крае, в Улетовском, Петровск-Забайкальском и Красночикойском районах обитает достаточно изолированная популяционная группировка соболя с темным мехом. Хребет Хамар-Дабан на юге Байкала населяет отличная от других районов популяция соболя, имеющая высокое генетическое разнообразие и самую светлую в изученном регионе окраску, что, скорее всего, обусловлено ее контактами с расположенной западнее восточно-саянской популяцией. Остальные выборки Байкальской горной страны демонстрируют относительно высокое генетическое разнообразие и промежуточные, по сравнению с предыдущими группировками, показатели окраски мехового покрова, что может быть результатом массовых внутрирегиональных транслокаций и миграций.
Авторы благодарят за предоставленую возможность для сбора материалов Ю.Е. Вашукевича, Е.К. Кондратьева, И.А. Шашкова, В.И. Романова, С.В. Щербакова (г. Иркутск). Работа выполнена согласно государственному заданию Института экологии растений и животных УрО РАН и при частичной поддержке РФФИ (проект № 17-04-00759).
Авторы заявляют об отсутствии конфликта интересов. Ни одного животного не было добыто специально для целей данного исследования. Животные отловлены легально с использованием гуманных методов добычи.
Список литературы
Тимофеев В.В., Надеев В.Н. Соболь. М.: Заготиздат, 1955. 404 с.
Тимофеев В.В., Павлов М.П. Соболь // Акклиматизация охотничье-промысловых зверей и птиц в СССР. Ч. 1. Киров: Волго-Вятское кн. изд-во, 1973. С. 51–105.
Еремеева К.М. Географическая изменчивость окраски соболей // Тр. Московского пуш.-мех. ин-та. 1952. Т. 3. С. 81–89.
Герасимова М.А. Качество меха баргузинского соболя, акклиматизированного в Томской области // Тр. ВНИИ живот. сырья и пушнины. 1958. Вып. 17. С. 139–160.
Бакеев Н.Н. Географическая изменчивость окраски меха соболя и ее динамика // Тр. ВНИИОЗ. Киров, 1976. Вып. 26. С. 26–54.
Монахов Г.И. Географическая изменчивость и таксономическая структура соболя фауны СССР // Тр. ВНИИОЗ. Киров, 1976. Вып. 26. С. 54–86.
Монахов Г.И., Бакеев Н.Н. Соболь. М.: Лесн. пром-сть, 1981. 240 с.
Монахов В.Г. Соболь Урала, Приобья и Енисейской Сибири: результаты реакклиматизации: Екатеринбург: Банк культурной информации, 1995. 154 с.
Бакеев Н.Н., Монахов Г.И., Синицын А.А. Соболь. Вятка: ВНИИОЗ, 2013. 336 с.
Монахов В.Г. Динамика размерной и фенетической структуры соболя в ареале. Екатеринбург: НИСО УрО РАН, Банк культурной информации, 2006. 202 с.
Monakhov V.G. Martes zibellina (Carnivora: Mustelidae) // Mammalian Species. 2011. V. 43 (1). P. 75 – 86. https://doi.org/10.1644/876.1
Wozencraft W.C. Order Carnivora. P. 532–628 // Mammal species of the world: a taxonomic and geographic reference. Eds. Wilson D.E. and Reeder D.M. 3-rd ed. Baltimore, Maryland: Johns Hopkins University Press, 2005.
Балмышева Н.П., Соловенчук Л.Л. Генетическая изменчивость гена цитохрома b митохондриальной ДНК соболя (Martes zibellina L.) магаданской популяции // Генетика. 1999. Т. 35. С. 1252–1257.
Рожнов В.В., Пищулина С.Л., Мещерский И.Г. и др. Генетическая структура соболя (Martes zibellina L.) Евразии: анализ распределения митохондриальных линий // Генетика. 2013. Т. 49. С. 251–258.
Каштанов С.Н., Свищёва Г.Р., Пищулина С.Л. и др. Географическая структура генофонда соболя (Martes zibellina L.): данные анализа микросателлитных локусов // Генетика. 2015. Т. 51. С. 78–88.
Шишацкая Д.И., Фрисман Л.В. Предварительные данные по генетической изменчивости соболя Среднего Приамурья (Martes zibellina L.): анализ двух микросателлитных локусов // Региональные проблемы. 2014. Т. 17 (2). С. 60–64.
Раковская Э.М., Давыдова М.И. Физическая география России. М.: Гуманит. изд. центр ВЛАДОС, 2001. Ч. 2. 304 с.
Zhu W., Wei Q., Xue S. et al. Isolation and characterization of microsatellite markers for the Sable, Martes zibellina (Mammalia: Mustelidae) // Pakistan J. Zool. 2017. V. 49. P. 1909–1912.
Davis C.S., Strobeck C. Isolation, variability, and cross-species amplification of polymorphic micro-satellite loci in the family Mustelidae // Mol. Ecol. 1998. V. 7. P. 1771–1788.
Basto M.P., Rodrigues M., Santos-Reis M. et al. Isolation and characterization of 13 tetranucleotide microsatellite loci in the Stone marten (Martes foina) // Conserv. Genet. Res. 2010. V. 2. P. 317–319. https://doi.org/10.1007/s12686-010-9217-2
Fleming M.A., Ostrander E.A., Cook J.A. Microsatellite markers for american mink (Mustela vison) and ermine (Mustela erminea) // Mol. Ecol. 1999. V 8. P. 1351–1362. https://doi.org/10.1182/blood-2006-08-044172
Vincent I.R., Farid A., Otieno C.J. Variability of thirteen microsatellite markers in American mink (Mustela vison) // Can. J. Anim. Sci. 2003. V. 83. P. 597–599. https://doi.org/10.4141/A03-001
Peakall R., Smouse P.E. GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research // Mol. Ecol. Notes. 2006. V. 6. P. 288–295.
Peakall R., Smouse P.E. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research – an update // Bioinformatics. 2012. V. 28. P. 2537–2539.
Pritchard J.K., Stephens M., Donnelly P. Inference of population structure using multilocus genotype data // Genetics. 2000.V.155. P. 945–959.
Earl D.A., von Holdt B.M. STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method // Conservation Genetics Resources. 2012. V. 4 (2). P. 359–361. https://doi.org/10.1007/s12686-011-9548-7
Петренко В.Д. Продуктивность таежных охотничьих угодий на севере Красноярского края. Красноярск: КрасГАУ, 2009. 180 с.
Obbard M.E. Fur grading and pelt identification // Wild Furbearer Management and Conservation in North America. Ontario: Min. Nat. Resources, 1987. P. 717–824.
Modorov M., Monakhov V., Mikryukov V. et al. Microsatellite multiplex assay for sable (Martes zibellina) and pine marten (Martes martes) // Mammal research. 2020. V. 65(4). P. 855‒862. https://doi.org/10.1007/s13364-020-00529-4
Каштанов С.Н., Рубцова Г.А., Лазебный О.Е. Исследование генетической структуры промышленной популяции соболя (Martes zibellina Linnaeus, 1758) по микросателлитным маркерам // Вестник ВОГиС. 2010. Т. 14. С. 426–431.
Фрисман Л.В., Шлюфман К.В., Брыкова А.Л. Генетическая структура популяции соболя (Martes zibellina) Буреинского нагорья по сборам одного охотничьего сезона // Региональные проблемы. 2019. Т. 22 (4). С. 46‒55. https://doi.org/10.31433/2618-9593-2019-22-4-46-55
Федорова Л.И., Кайгородова И.А. Микроэволюция байкальского соболя // Изв. Иркутского гос. ун-та. Серия “Биология. Экология”. 2014. Т. 7. С. 32–36.
Монахов Г.И., Тимофеев В.В. Предбайкалье и Забайкалье // Соболь, куницы, харза. М.: Наука, 1973. С. 84–95.
Монахов В.Г. Стабильность и изменчивость окраски соболя Прибайкалья // Современные проблемы охотоведения: Мат-лы нац. научно-практич. конф. Иркутск: ООО “Мегапринт”, 2019. С. 129–135.
Бакеев Н.Н. Географическая изменчивость соболей и структура населения // Соболь, куницы, харза. М.: Наука, 1973. С. 39–49.
Дополнительные материалы
- скачать ESM.docx
- Таблица S1. Значения генетических дистанций Fst при межгрупповом сравнении 14 выборок соболя Байкальской горной страны, рассчитанных по аллельному составу 11 микросаттелитных локусов
Таблица S2. Значения критерия t-Стьюдента при попарном сравнении 14 выборок соболя Байкальской горной страны по значению индекса окраски