

Экология, 2021, № 3, стр. 205-216
Экотопическая приуроченность неморальных реликтовых видов растений на хребте Хамар-Дабан (юг Восточной Сибири) по данным сеточного картирования
В. В. Чепинога a, b, *, М. В. Протопопова b, c, В. В. Павличенко b, c, С. В. Дудов d
a Центральный сибирский ботанический сад СО РАН
630090 Новосибирск, ул. Золотодолинская, 101, Россия
b Иркутский государственный университет
664003 Иркутск, ул. Карла Маркса, 1, Россия
c Сибирский институт физиологии и биохимии растений СО РАН
664033 Иркутск, ул. Лермонтова, 132, Россия
d Московский государственный университет им. М.В. Ломоносова
119234 Москва, Ленинские горы, 1, Россия
* E-mail: Victor.Chepinoga@gmail.com
Поступила в редакцию 01.10.2020
После доработки 12.11.2020
Принята к публикации 20.11.2020
Аннотация
Представлены результаты крупномасштабного сеточного картирования реликтовых видов растений (Anemone altaica Fisch. ex C.A. Mey., A. baicalensis Turcz., Corydalis bracteata (Steph.) Pers., Eranthis sibirica DC., Primula pallasii Lehm., Waldsteinia ternata (Steph.) Fritsch) в пределах неморального рефугиума на северном макросклоне хребта Хамар-Дабан (Республика Бурятия). Картирование проведено в нижней части бассейна р. Большой Мамай на модельном участке площадью 25 км2. Результаты исследования подтвердили общую для южно-сибирских неморальных реликтов закономерность: их приуроченность к долинам и берегам рек и ручьев. В то же время изученные виды различаются по своим экотопическим предпочтениям, в том числе по способности распространяться за пределы речной долины, диапазону предпочитаемых высот и устойчивости к нарушению растительного покрова. Кластерный анализ и оценка сопряженности видов с использованием непараметрического коэффициента конкордации Кендалла подтвердили наличие согласованности пространственного распределения модельных видов.
Большинство неморальных рефугиумов Южной Сибири в настоящее время довольно хорошо описаны и охарактеризованы [1–4 и др.], однако вопрос о встречаемости, пространственном распределении и экологических особенностях реликтовых видов, сохранившихся в этих рефугиумах, остается открытым. Некоторые исследователи считают, что реликтовые виды должны находиться в дисгармонии с современными условиями окружающей среды, поскольку концепция реликтовости предполагает редкую и спорадичную встречаемость вида в пределах своего дизъюнктивного ареала [5]. Однако в ряде случаев, напротив, отмечаются повышенное обилие и регулярная встречаемость реликтов в пределах участков их ареалов [6, 7]. Такая ситуация может объясняться тем, что каждый вид имеет свои уникальные экологические характеристики и особенности распространения. Несмотря на то, что ряд неморальных реликтов включен в федеральную [8] и/или региональные Красные книги, во многих случаях информация о распространении этих видов либо ограничена, либо недоступна, что не позволяет полноценно оценить состояние популяций или организовать их мониторинг.
Фрагментированность ареалов реликтовых видов в пределах рефугиума может указывать на его пространственную неоднородность, длительную изоляцию популяций друг от друга и наличие более мелких убежищ (микрорефугиумов), образовавшихся в периоды неблагоприятных климатических условий. Ранее нами было показано, что, несмотря на наступление благоприятных условий в голоцене, отдельные популяции реликтов на хр. Хамар-Дабан в настоящее время все еще остаются пространственно изолированными [9] и сохраняют относительную генетическую обособленность [10–13]. Для выявления степени пространственной изолированности популяций наиболее перспективен метод сеточного картирования. Этот подход является одним из наиболее эффективных в изучении географического распределения растений, а анализ пространственных данных, организованных в регулярную сетку, широко используется в исследованиях различного масштаба [14]: от локального и регионального [15–19 и др.] до континентального и глобального [20–22 и др.].
Цель данной работы – выявить экотопические и пространственные закономерности распространения реликтовых растений на северном макросклоне хр. Хамар-Дабан на основе результатов крупномасштабного сеточного картирования. Задачи исследования: 1) проведение картирования неморальных видов растений на модельном участке; 2) определение особенностей их экотопической приуроченности; 3) выделение групп сопряженности между модельными видами; 4) оценка вероятности контакта между популяциями соседних бассейнов рек.
МАТЕРИАЛ И МЕТОДЫ
Характеристика модельного участка
Неморальный рефугиум на северном макросклоне хр. Хамар-Дабан был описан Н.А. Еповой в середине ХХ в. [2]. Возможность существования здесь убежища для мезофильных неморальных видов объясняется гумидным и мягким климатом этой территории с большим количеством осадков летом и мощным снежным покровом зимой [23, 24]. Подобные условия характерны и для других неморальных рефугиумов юга Сибири [4]. В качестве модельной территории для сеточного картирования был выбран участок нижней части бассейна р. Большой Мамай (Республика Бурятия, Кабанский район) в пределах предгорной равнины и нижней полосы низкогорий. Выбор модельного участка обусловлен его положением в центральном секторе хребта (см. рис. 1), где встречается наибольшее (не менее 19 из 27) количество реликтовых видов растений [9]. В качестве границ территории картирования были приняты русла соседних с р. Большой Мамай рек – Осиновки на западе и Малого Мамая на востоке. На севере естественным рубежом считался берег оз. Байкал (456 м над ур. м.), с южной стороны граница проходила по отрогам хребта и была определена уже в ходе картирования по мере увеличения пересеченности рельефа и труднодоступности местности.
Рис. 1.
Модельный участок в низовьях р. Большой Мамай (хр. Хамар-Дабан, юг Восточной Сибири), охваченный сеточным картированием: а – Транссибирская железнодорожная магистраль; б – автомобильная трасса Р-258; в – просеки с высоковольтными линиями электропередач.
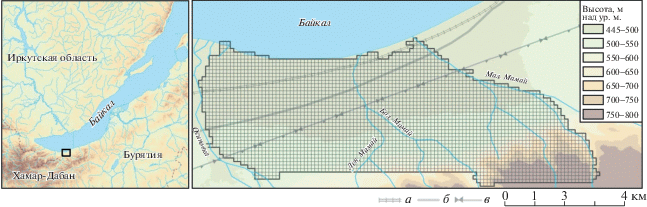
Истоки р. Большой Мамай находятся в 12 км от оз. Байкал на высотах 1500–1700 м. До самых низовий река собирает со склонов небольшие притоки. Наиболее крупным является р. Левый Мамай, который сливается с основным руслом в 1.6 км от устья. Несмотря на небольшую ширину русла, обычно не превышающую 10–12 м, в пределах предгорной равнины р. Большой Мамай образует достаточно обширную долину, изрезанную пересохшими руслами временных водотоков. Очевидно, разработанность речной долины связана с селевой активностью реки, что в целом характерно для многих водотоков северного макросклона хр. Хамар-Дабан [25] и подтверждается многократными переслоениями гумусового горизонта и грубообломочного материала в почве [26].
Река Малый Мамай берет свое начало в 9 км от оз. Байкал в пределах лесного пояса. Междуречье Большого Мамая и Малого Мамая в пределах предгорной равнины отличается пологим и спокойным рельефом. Основные крутосклонные гряды отрогов хребта заканчиваются в 3.7 км от берега оз. Байкал на высоте 550 м над ур. м. Предгорная равнина в междуречье большей частью трансформирована: преобладают вторичные березовые и хвойно-березовые леса на месте разновозрастных вырубок. В 1.3–1.4 км от устьев междуречье пересекается двумя просеками высоковольтных линий электропередач, тянущихся с запада на восток вдоль всего хребта. Кроме того, междуречье пересекают федеральная автомобильная трасса Р-258 (500–700 м от побережья оз. Байкал) и Транссибирская железнодорожная магистраль, проходящая почти по берегу озера. Приустьевая часть междуречья отделена от озера береговым валом и заболочена. Сооруженная более 120 лет назад железнодорожная насыпь способствовала обводнению, что привело к появлению водоемов непосредственно вдоль нее.
Река Осиновка имеет более протяженное (не менее 30 км) русло, достигающее в нижнем течении ширины 25–30 м. В пределах предгорной равнины русло почти не имеет притоков и “зажато” между двумя невысокими грядами, возможно, также селевого происхождения. Гряды образуют крутые обрывистые берега, которые возвышаются над руслом на высоту до 40 м и обрамляют реку почти на 3 км от створа долины. Правобережная гряда занимает около половины ширины междуречья Осиновки и Большого Мамая и образована крупноглыбовыми гранитными блоками, перекрытыми маломощной почвой. На гряде развиты разреженные темнохвойные леса с густым кустарниковым ярусом из стланиковой формы пихты (Abies sibirica Ledeb.). У автомобильной трассы гряда заканчивается. Значительная часть лесов наклонной равнины, особенно в районе железной дороги, нарушены в результате рубок и представлены различными сочетаниями смешанных хвойно-мелколиственных лесов. Заболоченность в приустьевой части реки незначительна и развита главным образом в пределах 100-метровой зоны вдоль берегового вала оз. Байкал.
Модельные виды
Для картирования была отобрана группа из шести видов весенних эфемероидов, относящихся к числу неморальных реликтов и включенных в Красную книгу Республики Бурятия [27]. Период цветения модельных видов продолжается с конца мая по начало июня, что делает их удобными объектами для проведения картирования на фоне еще неподнявшегося травостоя. Для характеристики видов использованы региональные флористические сводки [28, 29] и результаты собственных исследований [7, 9, 30, 31].
Anemone altaica Fisch. ex C.A. Mey. (ветреница алтайская) – восточно-европейский и преимущественно южно-сибирский вид. Участок его ареала на хр. Хамар-Дабан отделен от основного ареала в Западной Сибири дизъюнкцией порядка 700 км. Вид относится к группе родства европейского A. nemorosa L. и имеет серию викариантных таксонов на Российском Дальнем Востоке и в странах Восточной Азии.
Anemone baicalensis Turcz. (ветреница байкальская) – южно-сибирский эндем, описанный с хр. Хамар-Дабан. Здесь же расположен основной участок его ареала. Второй фрагмент ареала находится в Западном Саяне. В случае широкой трактовки вида ареал A. baicalensis охватывает южные регионы Российского Дальнего Востока и некоторых стран Восточной Азии, где произрастают несколько близкородственных ему таксонов.
Corydalis bracteata (Steph.) Pers. (хохлатка прицветниковая) – эндем Южной Сибири. Основной ареал вида расположен западнее 98-го меридиана, что определяет дизъюнкцию с популяциями на хр. Хамар-Дабан примерно в 600 км. Серия близкородственных видов распространена от Западной Европы до Нижнего Амура, однако в пределах хр. Хамар-Дабан родственных таксонов C. bracteata не имеет.
Eranthis sibirica DC. (весенник сибирский) – южно-сибирский эндем, описанный с хр. Хамар-Дабан. Распространен также в горах Восточного Саяна и изредка в Западном Саяне. Близкородственные виды встречаются на юге Российского Дальнего Востока и в странах Восточной Азии.
Primula pallasii Lehm. (примула Палласа) – западно-азиатский вид, доходящий на запад до Урала. В Европе замещается близким видом P. elatior (L.) Hill, в Европе и на Кавказе также имеется несколько близкородственных эндемичных таксонов. Популяции на хр. Хамар-Дабан оторваны от основного ареала на юге Западной Сибири и представляют собой крайний восточный фрагмент ареала вида.
Waldsteinia ternata (Steph.) Fritsch. (вальдштейния тройчатая) – южно-сибирский эндем, описанный с хр. Хамар-Дабан, где имеет популяции наибольшей плотности. Небольшие фрагменты ареала встречаются на отрогах Восточного Саяна и Западного Саяна. Рядом ученых W. ternata принимается в широком смысле, включая близкородственные таксоны, произрастающие на территории Центральной и Юго-Восточной Европы, Западной Сибири, Российского Дальнего Востока и стран Восточной Азии.
Методика сеточного картирования
Картирование проводили в конце мая и первой половине июня (25.05.2014; 01–04.06.2014; 13–18.06.2014; 31.05–02.06.2015; 02–05.06.2018) на модельном участке площадью 25 км2 (см. рис. 1). Максимальное расстояние с севера на юг (от 51.421° до 51.462° с.ш.) составило 4.5 км, а с запада на восток – 6.3 км (от 104.731° до 104.782° в.д.). Перепад высот составил около 500 м, варьируя от 460 м (урез воды оз. Байкал) до 700 м (водораздел рек Осиновка и Большой Мамай) и 950 м над ур. м. (водораздел рек Большой Мамай и Малый Мамай).
На модельный участок была наложена координатная сетка с размером ячейки 0.001° × 0.001°, что на местности примерно соответствовало 110 м по широте и 70 м по долготе. В общей сложности картирование видов проведено в 3261 ячейке в ходе пеших маршрутов, которые были проложены с запада на восток параллельно одной широте. Границы ячеек на местности определяли с использованием портативных GPS-навигаторов GPSMAP 64ST (Garmin, США) по смене одной тысячной доли градуса. Общая протяженность пройденных маршрутов составила более 230 км.
Встречаемость видов оценивали визуально по четырехбалльной шкале: 0 – вид отсутствует, 1 – вид встречается редко, 2 – вид встречается довольно часто, 3 – вид обычен. В пределах каждой ячейки встречаемость видов оценивали дважды каждые 35 м (сначала для первой, а затем для второй половины ячейки). Оба значения использовали для вычисления итогового показателя встречаемости видов в ячейке. Расчет проводили по следующему алгоритму: если разница между значениями показателя встречаемости вида в половинах ячейки составляла одну ступень (например, 0–1, 1–2, 2–3), то в качестве итогового принимали большее значение; если разница была равна двум (0–2, 1–3) или трем (0–3) ступеням, то итоговое значение вычисляли как большее значение минус один. Например, если в первой половине ячейки встречаемость вида была оценена как вид обычен (3), а во второй половине вид отсутствовал (0), то, учитывая разницу значений в три ступени, итоговый показатель встречаемости вида в ячейке вычисляли как 3 – 1 = 2. Для оценки сопряженности видов с помощью кластерного анализа итоговые показатели встречаемости были условно переведены в значения проективного покрытия с шагом в один порядок: 1 = 0.1%, 2 = 1%, 3 = 10%.
Одновременно с картированием проводилось визуальное наблюдение за экотопической и фитоценотической приуроченностью модельных видов. Отдельные факты наблюдений как в пределах модельного участка, так и за его пределами использованы при описании результатов исследования.
Статистический анализ сопряженности
Для подтверждения предположения о сопряженном пространственном распределении модельных видов (и соответственно о близости их экологических ниш) был использован алгоритм, описанный П. Лежандром [32]. Алгоритм включал три этапа: 1) исходные данные были организованы в виде матрицы с видами (столбцы) и ячейками (строки) картирования, на их пересечении – значение встречаемости, а на основе матрицы были рассчитаны попарные корреляции Пирсона между видами; 2) матрица корреляций была пересчитана в матрицу расстояний 1 – r, где r – коэффициент корреляции Пирсона, на основе которой, используя кластерный анализ по методу Варда, были определены предварительные группы сопряженных видов; 3) на основе исходной матрицы встречаемости видов в ячейках была проведена оценка сопряженности видов друг с другом, а также видов внутри групп с использованием непараметрического коэффициента конкордации Кендалла (W).
При расчете коэффициента W была проверена нулевая гипотеза об отсутствии сопряженности распределения. Для определения степени сопряженности распределения анализируемого вида с прочими, включенными в исследование, был также рассчитан апостериорный коэффициент конкордации Wj. Во всех случаях расчеты проводили при уровне значимости p < 0.01 с поправкой Хольма и выполняли при 999 пермутациях. Анализ данных проводили с использованием функции kendall.global пакета vegan v. 2.5-6 [33] и базового статистического пакета stats в среде R v. 3.6.1 (функции “cor”, “dist”, “hclust”). Степень конкордации (сопряженности пространственного распределения видов) оценивали по следующей условной шкале W: 0–0.4 – низкая, 0.4–0.6 – умеренная, 0.6–1 – высокая.
РЕЗУЛЬТАТЫ
Anemone altaica обнаружен в 349 ячейках, что составляет 10.7% от их общего числа (табл. 1, рис. 2). В пределах модельного участка преобладали разреженные популяции, расположенные близ русел рек и ручьев. Широкая полоса скоплений A. altaica вдоль р. Большой Мамай соответствует руслам временных водотоков, которые не были обводнены в момент картирования. Крайне спорадичны популяции вида в междуречье Осиновки и Большого Мамая, где расположена горная гряда, а водотоки редки. Этот вид встречается не только в лесном, но также и в субальпийском поясе хр. Хамар-Дабан [9]. По нашим наблюдениям, вид поднимается до высоты 1413 м над ур. м., где выше границы леса располагаются наиболее массовые популяции.
Таблица 1.
Встречаемость видов в ячейках сеточного картирования в бассейне нижнего течения р. Большой Мамай (хр. Хамар-Дабан, юг Восточной Сибири)
| Вид | Встречаемость вида | Всего ячеек | Доля (%) от общего числа (3261) ячеек | ||
|---|---|---|---|---|---|
| редко | довольно часто | обычно | |||
| Anemone altaica | 226 | 84 | 39 | 349 | 10.7 |
| Anemone baicalensis | 427 | 496 | 472 | 1395 | 42.8 |
| Corydalis bracteata | 226 | 121 | 70 | 417 | 12.8 |
| Eranthis sibirica | 494 | 177 | 54 | 725 | 22.2 |
| Primula pallasii | 44 | 3 | 0 | 47 | 1.4 |
| Waldsteinia ternata | 380 | 346 | 204 | 930 | 28.5 |
Рис. 2.
Пространственное распределение картированных видов в пределах модельного участка в бассейне нижнего течения р. Большой Мамай (хр. Хамар-Дабан, юг Восточной Сибири). Встречаемость видов: 1 – редко, 2 –часто, 3 – обычен.
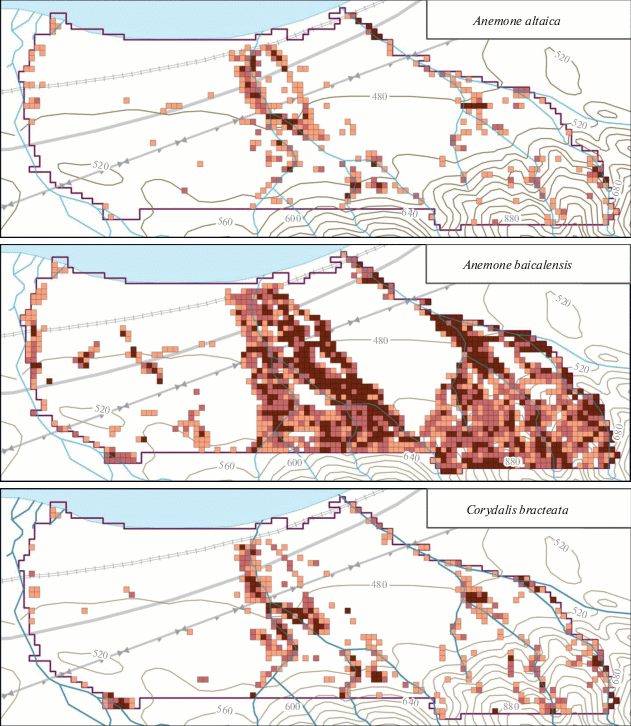
Рис. 2.
Окончание.
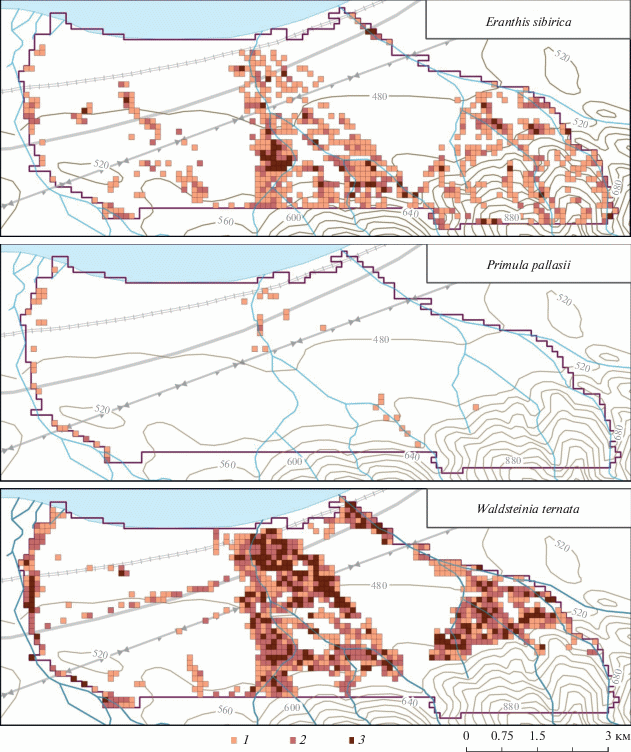
Anemone baicalensis – наиболее частый и массовый вид среди модельных. Его популяции отмечены более чем в 40% ячеек. Количество ячеек, где вид отмечен “редко”, “довольно часто” и “обычно”, почти равное (см. табл. 1). На рис. 2 видно, что на предгорной равнине популяции A. baicalensis четко указывают положение постоянных или временных водотоков. Так, в междуречье Осиновки и Большого Мамая часть популяций линейно вытянуты по направлению к Байкалу. Скопление соответствует сухому ложу временного водотока, проходящему вдоль восточного края горной гряды и отсутствующему на картах. Это пересохшее ложе опознается на картосхемах в виде скоплений популяций и других видов, распространение которых также связано с водотоками (см. рис. 2). Популяции A. baicalensis практически отсутствуют на водораздельных нарушенных пространствах со вторичными березовыми лесами (междуречье Большого Мамая и Малого Мамая), на заболоченных приустьевых участках, а также на гряде между реками Осиновка и Большой Мамай, где доминирование стланиковой пихты создает условия, не пригодные для произрастания и остальных модельных видов. С увеличением абсолютной высоты популяции A. baicalensis из бассейнов соседних водотоков постепенно смыкаются в горно-таежном поясе. На примере междуречья рек Большой Мамай и Малый Мамай видно, что смыкание популяций начинается приблизительно на отметке 600–700 м над ур. м. Именно в горно-таежном поясе, где вид встречается с высоким постоянством, Н.А. Еповой были описаны оригинальные для хр. Хамар-Дабан “пихтачи с покровом из байкальской анемоны” [34]. Непрерывное распространение A. baicalensis фактически сохраняется до высоты 1200 м над ур. м., где на северном макросклоне хребта расположен субальпийский пояс [9].
Corydalis bracteata относится к тому же приручьевому комплексу видов, что и A. altaica. Схемы распространения этих двух видов довольно схожи между собой: C. bracteata был найден в 12.8% ячеек (см. табл. 1 и рис. 2), что близко к встречаемости A. altaica – 10.7%. Однако в отличие от последнего C. bracteata не отмечается в высокогорьях – по нашим наблюдениям [9], он поднимается лишь до высоты 1064 м над ур. м. Это типичный вид лесного пояса, представленный массовыми популяциями непосредственно близ русел либо по ложам высохших водотоков, чем и объясняется небольшое число ячеек (16.8%), где встречаемость вида оценена как “обычно”.
Eranthis sibirica отмечен в 22.2% ячеек, из которых в каждой 4-й − “довольно часто”, а в каждой 13-й – “обычно” (см. табл. 1 и рис. 2). В пределах предгорной равнины E. sibirica вместе с A. altaica и C. bracteata участвует в формировании зарослей приручьевых раннецветущих эфемероидов. В то же время с увеличением абсолютной высоты E. sibirica часто удаляется от русел и стабильно (но с меньшим обилием) встречается также по склонам различной крутизны и экспозиции в составе горно-таежных лесов вместе с A. baicalensis. Следует отметить, что E. sibirica распространен как в лесном, так и в высокогорном поясах [9, 31], поднимаясь на хр. Хамар-Дабан, по нашим наблюдениям, до высоты не менее 1700 м. В среднегорьях ареал вида, очевидно, имеет непрерывный характер.
Primula pallasii обнаружен в 47 ячейках (1.4%), в большинстве из которых вид встречался “редко” (см. табл. 1). Все ячейки расположены на модельном участке вдоль русел главных водотоков (см. рис. 2), а основной ареал вида находится в высокогорьях, в субальпийском поясе, где он является одним из диагностических видов высокотравных лугов с доминированием Rhaponticum carthamoides (Willd.) Iljin s.l. [35, 36]. В низовьях рек P. pallasii оказывается, очевидно, только в случае прорастания семян, смытых вниз потоками воды. Поэтому местонахождения регулярно отмечены вдоль русел Осиновки и Большого Мамая, верхние течения которых находятся в высокогорьях. Полное отсутствие находок вдоль русла р. Малый Мамай может объясняться тем, что эта река начинается в пределах лесного пояса, т.е. ниже пределов высокогорного ареала вида.
Waldsteinia ternata обнаружен в 28.5% ячеек (см. табл. 1), из них в каждой третьей отмечен “довольно часто”, а в каждой пятой − “обычно”. Вид приурочен к долинам и берегам рек (см. рис. 2). В приустьевой части междуречья Большого Мамая и Малого Мамая W. ternata был впервые обнаружен на осоковом болоте, на верхушках кочек Carex juncella (Fries) Th. Fr. Следует отметить серию местонахождений вдоль автомобильной трассы и на просеках линий электропередач. Являясь светолюбивым видом, W. ternata густо разрастается по опушкам и вдоль дорог. Активное расселение вдоль линейных инженерных сооружений и нарушенных участков указывает на антропотолерантность вида. В целом картосхема распространения W. ternata в низовьях рек близка к паттернам распространения A. baicalensis и E. sibirica, однако в отличие от последних он является типичным лесным низкогорным видом, и его популяции обычно не поднимаются выше 600 м над ур. м. По нашим наблюдениям, на хр. Хамар-Дабан (за пределами модельного участка) популяции W. ternata отмечались на высотах от 600 до 700 м только на нарушенных участках: вдоль дорог и туристических троп.
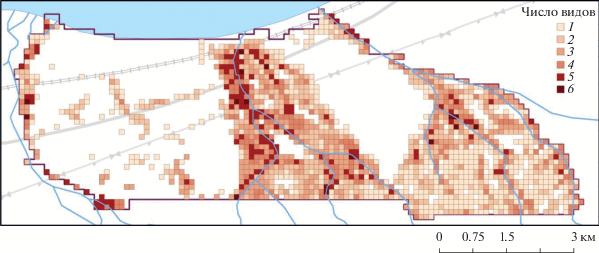
В общей сложности модельные виды были обнаружены в 1580 ячейках из 3261, что составляет 48.5%. Количество ячеек с разным числом видов и распределение непустых ячеек в пространстве представлены в табл. 2 и на рис. 3 соответственно. В трети всех непустых ячеек (30.3%) было выявлено присутствие лишь одного модельного вида – в большинстве случаев это был один из самых массовых видов: A. baicalensis или W. ternata. В каждой четвертой непустой ячейке (24.4%) было зафиксировано одновременно по два вида: чаще всего были представлены пары A. baicalensis–E. sibirica и A. baicalensis–W. ternata. Ячейки с тремя видами составили 21.6% от общего числа непустых ячеек. В большинстве этих ячеек присутствовал A. baicalensis, который сочетался с парой E. sibirica–W. ternata. Таким образом, две трети, или 76%, непустых ячеек содержали не более трех видов. Все шесть модельных видов одновременно встретились только в 17 ячейках.
Таблица 2.
Совместная встречаемость видов в пределах ячеек сеточного картирования
| Количество видов, отмеченных в одной ячейке | Количество ячеек | Доля (%) от количества непустых ячеек (1580) |
|---|---|---|
| 1 | 479 | 30.3 |
| 2 | 385 | 24.4 |
| 3 | 342 | 21.6 |
| 4 | 221 | 14.0 |
| 5 | 136 | 8.6 |
| 6 | 16 | 1.0 |
Рис. 3.
Распределение ячеек с разным числом модельных видов в пределах модельного участка в бассейне нижнего течения р. Большой Мамай (хр. Хамар-Дабан, юг Восточной Сибири).
В результате кластерного анализа (рис. 4) на первом иерахическом уровне была выделена пара строго приручьевых видов A. altaica и C. bracteata (группа I). Лесные светолюбивые виды W. ternata, A. baicalensis и E. sibirica, отличающиеся наибольшей встречаемостью и широкой экологической валентностью, формировали группу II. Редкий вид P. pallasii на ключевой территории не вошел ни в одну из обозначенных групп.
Рис. 4.
Дендрограмма кластерного анализа повидовых корреляций Пирсона методом Варда: I, II – номера групп, использованные в анализе сопряженности (см. табл. 3). Шкала расстояний приведена под дендрограммой.
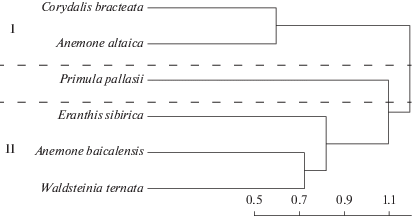
Коэффициент конкордации Кендалла (W) при рассмотрении шести видов составил 0.47, что отражает умеренную сопряженность их пространственного распределения (табл. 3). Коэффициент Wj оказался наименьшим для P. pallasii (0.13), а для остальных видов находился в диапазоне от 0.45 до 0.52. Степень сопряженности пространственного распределения видов в пределах выделенных групп характеризуется как высокая для группы I (W = 0.83; A. altaica и C. bracteata) и умеренная – для группы II (W = 0.67; W. ternata, A. baicalensis и E. sibirica).
Таблица 3.
Значения коэффициента конкордации Кендалла W для групп видов и Wj для отдельных видов внутри групп, выделенных на основе кластерного анализа
| Вид | Для всех видов | Для групп | |||
|---|---|---|---|---|---|
| Wj | W | № | W | Wj | |
| Corydalis bracteata | 0.50 | 0.47 | I | 0.83 | 0.83 |
| Anemone altaica | 0.48 | 0.83 | |||
| Primula pallasii | 0.13 | – | – | – | |
| Eranthis sibirica | 0.49 | II | 0.67 | 0.51 | |
| Anemone baicalensis | 0.52 | 0.71 | |||
| Waldsteinia ternata | 0.45 | 0.64 | |||
ОБСУЖДЕНИЕ
Известно, что состав неморальных реликтов Южной Сибири представлен видами различного происхождения, имеющими родственные виды (либо собственный ареал) в Европе (Dryopteris filix-mas (L.) Schott, Daphne mezereum L.), регионах Восточной Азии (Festuca extremorientialis Ohwi, Galium paradoxum Maxim.); некоторые обладают трансевроазиатскими (Epilobium montanum L., Galium triflorum Michx.) ареалами [4, 37]. Модельные виды, отобранные нами для картирования (см. раздел “Материал и методы”), также отличаются по происхождению, однако, видимо, происхождение не имеет заметного влияния на распространение видов на хр. Хамар-Дабан. Наблюдается закономерная и характерная для многих неморальных реликтов приуроченность к долинам и берегам рек и ручьев [9]. При этом для собственно приручьевых видов временное отсутствие открытой воды рядом с местом их произрастания не считается критическим. Репрезентативными примерами таких видов являются A. altaica и C. bracteata. За пределы речных долин выходят A. baicalensis, E. sibirica и W. ternata.
Изученные нами виды отличаются по некоторым экологическим особенностям, например по высотному распределению. Несмотря на то, что модельный участок не охватывает высотный диапазон распространения исследованных видов целиком, наши наблюдения на других участках северного макросклона хр. Хамар-Дабан позволяют составить более цельную картину об их высотном распределении. Распространение W. ternata ограничено нижней полосой лесного пояса (500–600 м), почти полностью вписываясь в район картирования. Распространение C. bracteata включает как нижнюю, так и среднюю полосы лесного пояса, не удаляясь при этом от водотоков. Диапазон распространения A. baicalensis охватывает целиком лесной пояс, а также часть субальпийского. Ареалы A. altaica и E. sibirica протягиваются через все высотные пояса хребта, однако в пределах модельного участка A. altaica ведет себя как строго приручьевой вид, в то время как E. sibirica свободно удаляется от русел водотоков. Настоящим высокогорным видом является P. pallasii, появление которого в предгорьях, очевидно, происходит только в результате смыва семян и/или вегетативных частей растений потоками воды.
Согласно нашим наблюдениям в ходе картирования, почти для всех модельных видов характерны черты, присущие антропофобным растениям, избегающим участков с нарушенным растительным покровом, и только W. ternata проявлял признаки антропотолерантности, положительно реагируя на нарушения растительности, разрастаясь на опушках леса, по придорожным канавам, вдоль троп и лесных дорог. Именно вдоль троп W. ternata проникает в горы до высоты 700 м.
Совместное отображение распространения модельных видов на одной картосхеме (см. рис. 4) однозначно указывает на их неслучайное распределение. Очевидно, что общий контур распределения непустых ячеек определяется присутствием трех наиболее массовых видов. Однако и остальные виды лишь незначительно выходят за пределы распространения массовых, группируясь в первую очередь близ водотоков. Выявленные закономерности подтверждают ранее выдвинутое предположение о том, что ареал A. baicalensis, самого массового вида среди реликтов хр. Хамар-Дабан, территориально соответствует всему неморальному рефугиуму на его северном макросклоне [38].
Кластерный анализ подтвердил наличие пространственной сопряженности распространения большинства модельных и двух групп сопряженных видов. В качестве таких групп выделилась пара приручьевых видов (A. altaica и C. bracteata) и три вида преимущественно лесных растений с более широкой экологической валентностью (A. baicalensis, E. sibirica и W. ternata). Интересно отметить положение E. sibirica, который в пределах предгорной равнины ведет себя как приручьевой вид, встречаясь совместно с C. bracteata и A. altaica. Однако по мере увеличения абсолютной высоты и пересеченности рельефа E. sibirica все чаще удаляется от водотоков и регулярно, хотя и с меньшим обилием, отмечается совместно с A. baicalensis в составе горно-таежных лесов. Результаты кластерного анализа также показали, что E. sibirica объединяется не с приручьевыми, а с лесными видами, занимающими, как и он, более широкую экологическую нишу.
По нашим наблюдениям, совместная встречаемость видов сопряженных групп сохраняется и на других участках северного макросклона хр. Хамар-Дабан. Это указывает на то, что при различиях в происхождении, экотопической приуроченности, диапазоне высотного распределения и реакции на антропогенное нарушение растительного покрова неморальные реликты обладают определенным единством, связанным с их длительным сосуществованием в рефугиуме. Одним из результатов сосуществования являются оригинальные типы лесных сообществ, выявленные на хр. Хамар-Дабан [34, 39], в состав блока диагностических видов которых в полном составе входит группа I.
Необходимо отметить, что в пределах модельного участка ячейки с наибольшим количеством видов не отличаются какой-либо вариабельностью местообитаний. Подобные ячейки, как правило, расположены в пределах предгорной равнины на выположенных участках с руслами водотоков или в непосредственной близости к ним. В таких условиях для наличия полного (или почти полного) набора модельных видов достаточно сочетание приручьевых местообитаний с лесными. Леса представлены здесь пойменными смешанными (Abies sibirica Ledeb., Populus suaveolens Fisch., Picea obovata Ledeb., Pinus sibirica Du Tour) разнотравными фитоценозами, наиболее богатыми на хр. Хамар-Дабан реликтовыми видами растений [9, 34].
Охват модельным участком территории, относящейся к бассейнам трех рек, позволил выявить наличие контактов между популяциями некоторых реликтовых видов из разных речных бассейнов. Очевидное преимущество в образовании контактов имеют массовые и с широким диапазоном распространения по высоте виды. Если приуроченность таких видов к долинам рек на предгорной равнине ограничивает вероятность контактов, то в условиях среднегорий или высокогорий контакты более вероятны уже по причине более густой речной сети. Для гумидных условий центрального сектора хр. Хамар-Дабан, где расположен модельный участок, полное или частичное объединение популяций соседних бассейнов в единую метапопуляцию может быть констатировано для A. baicalensis и E. sibirica. Наличие контактов между популяциями можно также предположить для A. altaica, C. bracteata и P. pallasii.
Иная закономерность пространственного распространения характерна для W. ternata. В отличие от остальных видов ареал W. ternata охватывает лишь нижнюю полосу лесного пояса, а популяции соседних речных бассейнов отчетливо дистанцированы друг от друга. Это позволяет предполагать, что в пределах модельного участка контакт между популяциями может быть ограничен распространением данного вида вдоль линейных инженерных конструкций (автомобильных магистралей, линий электропередач). На других участках предгорий хр. Хамар-Дабан, где расстояние между руслами рек может быть больше, случайный перенос семян человеком, очевидно, является единственным механизмом, связывающим популяции с разных бассейнов.
Таким образом, анализ данных сеточного картирования показал, что реликтовые виды хамар-дабанского рефугиума формируют популяции, плотность и границы распространения которых в первую очередь определяются их экотопическими предпочтениями и устойчивостью к нарушениям растительного покрова. Проведенное нами исследование позволило впервые для рефугиумов Южной Сибири оценить плотность популяций реликтовых растений в крупном масштабе. До сих пор данные с хр. Хамар-Дабан были ограничены единичными местонахождениями, опубликованными в работах, посвященных флористическим находкам [40‒42 и др.], либо гербарными образцами, большинство из которых собирали в одних и тех же локалитетах [7].
Помимо экологии и реакции на нарушения, распределение видов в определенных ситуациях может корректироваться также причинами, связанными с историей растительного покрова хр. Хамар-Дабан: относительной удаленностью от микрорефугиумов плейстоцена [9], пределами распространения грязе-каменных потоков во время катастрофических селевых явлений [25, 26]. Выявленные нами закономерности позволят отделить ситуации, связанные с экологией видов, и рассмотреть влияние на распространение неморальных реликтов непосредственно исторических факторов.
Анализ результатов крупномасштабного сеточного картирования позволил выявить следующие закономерности неморальных реликтовых видов растений: 1) реликты обладают определенными экологическими особенностями, связанными со своими экотопическими предпочтениями, в том числе в пределах группы модельных раннецветущих видов; 2) различия между модельными видами заключаются, в частности, в способности распространяться за пределы речной долины, в диапазоне предпочитаемых высот и реакции на нарушения растительного покрова; 3) широкая экологическая валентность некоторых модельных видов определяет возможность формирования ими единой метапопуляции, объединяющей локальные популяции из соседних речных бассейнов; 4) несмотря на различное происхождение неморальных реликтов, их длительное совместное существование в пределах рефугиума привело к возникновению групп сопряженных видов и, как следствие, оригинальных растительных сообществ.
Полученный в ходе выполнения настоящей работы опыт позволяет согласиться с утверждением о широких возможностях анализа и применения сеточных данных [14]. В соответствии с требованиями, предъявляемыми к ведению Красных книг в Российской Федерации [43] и требованиями Международного союза охраны природы при установлении категорий охраны [44], крупномасштабное сеточное картирование может быть рекомендовано для оценки плотности популяций, их состояния и организации мониторинга видов, нуждающихся в охране. Получаемые в ходе картирования количественные данные можно использовать для определения пороговых значений между категориями статуса редкости и уязвимости.
Авторы выражают глубокую признательность О.А. Аненхонову (ИОЭБ СО РАН, г. Улан-Удэ) за ценные советы и замечания по тексту рукописи статьи, а также благодарность студентам ИГУ (Э.М. Байрамова, В.А. Белова, М.А. Вилор, Е.М. Гладких, Е.Д. Золотовская, В.П. Иванова, А.Д. Коновалов, А.В. Мишина, А.В. Рудых, А.А. Толмачева, А.М. Скорнякова, А.В. Юдинцева) и Иркутского государственного аграрного университета (С.О. Быстров) за участие в полевых работах по картированию.
Работа выполнена в рамках государственных заданий ЦСБС СО РАН (рег. № АААА-А2-121011100007-6), СИФИБР СО РАН (тема № 0343-2019-006), МГУ им. М.В. Ломоносова (рег. № АААА-А16-116021660039-1), а также при поддержке РФФИ (проекты № 20-45-380009, 19-54-53014-ГФЕН, 16-34-60135-мол_а_дк, 14‑04-31350-мол_а). Часть полевых исследований, проведенных в течение полевого сезона 2018 г. в междуречье рек Большой Мамай и Осиновка, выполнена при частичной финансовой поддержке РНФ (проект № 17-74-10074).
Список литературы
Крылов П.Н. Липа в предгорьях Кузнецкого Алатау // Изв. Томского ун-та. 1891. Вып. 1. С. 3–40.
Епова Н.А. Реликты широколиственных лесов в пихтовой тайге Хамар-Дабана // Изв. Биол.-геог. НИИ при Иркут. гос. ун-те им. А.А. Жданова. Иркутск, 1956. Т. XVI. Вып. 1–4. С. 25–61.
Куминова А.В. Телецкий рефугиум третичной растительности // Изв. Вост.-Сиб. фил. АН СССР. 1957. Т. 2. С. 104–108.
Положий А.В., Крапивкина Э.Д. Реликты третичных широколиственных лесов во флоре Сибири. Томск: Изд-во Томского гос. ун-та, 1985. 158 с.
Толмачев А.И. Введение в географию растений. Л.: Изд-во Ленингр. гос. ун-та, 1974. 244 с.
Еленевский А.Г., Радыгина В.И. О понятии “реликт” и реликтомании в географии растений // Бюл. МОИП. Отд. биол. 2002. Т. 107. Вып. 3. С. 39–48.
Чепинога В.В., Мишина А.В., Протопопова М.В. и др. Новые данные о распространении некоторых неморальных реликтовых растений в предгорьях хребта Хамар-Дабан (Южное Прибайкалье) // Бот. журн. 2015. Т. 100. № 5. С. 478–489.
Красная книга Российской Федерации: растения и грибы / Под ред. Трутнева Ю.П. и др. М.: Тов-во науч. изд. КМК, 2008. 855 с.
Чепинога В.В., Протопопова М.В., Павличенко В.В. Выявление вероятных плейстоценовых микрорефугиумов на северном макросклоне хребта Хамар-Дабан (Южное Прибайкалье) // Сиб. экол. журн. 2017б. № 1. С. 44–50. [Chepinoga V.V., Protopopova M.V., Pavlichenko V.V. Detection of the most probable Pleistocene microrefugia on the northern macroslope of the Khamar-Daban Ridge (Southern Prebaikalia) // Contemp. Probl. of Ecology. 2017. V. 10, № 1. P. 38–42.]
Протопопова М.В., Павличенко В.В., Гнутиков А.А. и др. Использование генетических маркеров для оценки состояния реликтовых видов растений Байкальской Сибири // Вестн. Росс. ун-та дружбы народов. Сер. “Экология и безопасность жизнедеятельности”. 2015. № 4. С. 28–36.
Протопопова М.В., Павличенко В.В., Коновалов А.Д. и др. Перспективы использования внутренних транскрибируемых спейсеров (ITS1 и ITS2) для идентификации редких видов растений на примере рода Waldsteinia (Rosaceae) // Изв. Иркут. гос. ун-та. Сер. “Биология. Экология”. 2016. Т. 17. С. 5–11.
Протопопова М.В., Павличенко В.В., Орлова Д.А., Чепинога В.В. Филогеографическая структура Anemone baicalensis (Ranunculaceae) на основе полиморфизма пластидной ДНК (trnL-trnF) как подтверждение существования плейстоценовых микрорефугиумов на хребте Хамар-Дабан (Южное Прибайкалье) // Изв. Иркут. гос. ун-та. Сер. “Биология. Экология”. 2019. Т. 30. С. 3–15.
Protopopova M., Pavlichenko V., Gnutikov A., Chepinoga V. DNA Barcoding of Waldsteinia Willd. (Rosaceae) species based on ITS and trnH-psbA nucleotide sequences // Information Technologies in the Research of Biodiversity / Bychkov I., Voronin V. Eds. Springer Proceedings in Earth and Environmental Sciences. Cham: Springer Nature Switzerland, 2019. P. 107–115.
Серёгин А.П. Сеточное картирование флоры: мировой опыт и современные тенденции // Вестн. Тверского гос. ун-та. Сер. “Биология и экология”. 2013. Вып. 32. С. 210–245.
Firbank L.G., Ellis N.E., Hill M.O. et al. Mapping the distribution of weeds in Great Britain in relation to national survey data and to soil type // Weed Research. 1998. V. 38. P. 1–10.
Petřík P., Bruelheide H. Species groups can be transferred across different scales // J. of Biogeography. 2006. V. 33. № 9. P. 1628–1642.
Чепинога В.В., Солодянкина С.В., Иванова В.П. Распространение некоторых синантропных растений в историческом центре города Иркутска (Восточная Сибирь) // Вестн. Томского гос. ун-та. Биология. 2016. № 2(34). С. 87–100.
Чепинога В.В., Петухин В.А., Стальмакова Д.П. Результаты сеточного картирования сводки “Флора Центральной Сибири” (1979) в цифровом формате: итоги и перспективы использования // Растит. мир Азиат. России. 2017. № 3(27). С. 70–78.
Чепинога В.В., Солодянкина С.В., Иванова В.П. Особенности распространения некоторых культивируемых древесных растений в историческом центре города Иркутска (Восточная Сибирь) // Вестн. Томского гос. ун-та. Биология. 2017. № 40. С. 102–115.
Mutke J., Barthlott W. Patterns of vascular plant diversity at continental to global scales // Biologiske Skrifter. 2005. V. 55. № 4. P. 521–531.
Kreft H., Jetz W. Global patterns and determinants of vascular plant diversity // Proceed. of the National Academy of Sciences. 2007. V. 104. № 14. P. 5925–5930.
Liu Y., Su X., Shrestha N. et al. Effects of contemporary environment and Quaternary climate change on drylands plant diversity differ between growth forms // Ecography. 2018. V. 42. P. 334–345.
Ладейщиков Н.П., Филиппов А.Х., Зедгенидзе Е.Г. и др. Осадки и режим увлажнения // Структура и ресурсы климата Байкала и сопредельных пространств / Под ред. Ладейщикова Н.П. Новосибирск: Наука, 1977. С. 98–125.
Расписание погоды rp5, 2004–2020. Интернет-ресурс. Адрес доступа: URL: https://rp5.ru (последнее обращение: 12.09.2020)
Ивановский Л.Н. Особенности развития долин на южном побережье Байкала в антропогене // История развития речных долин и проблемы мелиорации земель. Сибирь и Дальний Восток. Новосибирск: Наука, 1979. С. 55–62.
Макаров С.В., Черкашина А.А. Селевой морфолитогенез в центральной части Саяно-Байкальского станового нагорья // Геогр. и прир. ресурсы. 2019. № 1. С. 96–105. [Makarov S.A., Cherkashina A.A. Debris flow morpholithogenesis in the central part of the Sayan-Baikal stanovoi highlands // Geography and Natural Resources. 2019. V. 40, № 1. P. 54–62.]
Красная книга Республики Бурятия: редкие и находящиеся под угрозой исчезновения виды животных, растений и грибов / Под ред. Пронина Н.М. 3-е изд. перераб. и доп. Улан-Удэ: Изд-во БНЦ СО РАН, 2013. 688 с.
Сосудистые растения советского Дальнего Востока. В 8-и т. / Под ред. Харкевича С.С. Л.; СПб.: Наука, 1985–1996.
Флора Сибири / Под ред. Малышева Л.И. и др. В 14-и т. Новосибирск: Наука, 1987–2003.
Чепинога В.В., Степанов Н.В., Протопопова М.В., Павличенко В.В. О распространении Waldsteinia ternata (Rosaceae) в Западном Саяне // Бот. журн. 2019. Т. 104. № 8. С. 1203–1210.
Heim R.J., Chepinoga V.V. Subalpine tall-herb vegetation patterns: a case study from the Khamar-Daban Range (southern Baikal region, Eastern Siberia) // Botanica Pacifica. 2019. V. 8. № 1. P. 39–49.
Legendre P. Species associations: The Kendall coefficient of concordance revisited // J. of Agricultural, Biological, and Environmental Statistics. 2005. V. 10. № 2. P. 226–245.
Oksanen J., Blanchet G., Friendly M. et al. vegan: Community Ecology Package. R package version 2.5-6. 2019. https://CRAN.R-project.org/package=vegan. Last accessed 12 September 2020.
Епова H.A. К характеристике пихтовой тайги Хамар-Дабана // Тр. Бурят. комплексного НИИ. Серия биол.-почв. Улан-Удэ, 1961. Вып. 4. С. 141–163.
Ermakov N., Shaulo D., Maltseva T. The class Mulgedio-Aconitetea in Siberia // Phytocoenologia. 2000. V. 30. № 2. P. 145–192.
Чепинога В.В. Ценотическое разнообразие и географические особенности высокотравных лугов хребта Хамар-Дабан // Современные проблемы биологии, экологии и почвоведения: Мат-лы междунар. науч. конф., посвящ. 100-летию высш. биол. образования в Вост. Сибири. Иркутск: изд‑во Иркут. гос. ун-та, 2019. С. 119–121.
Малышев Л.И., Пешкова Г.А. Особенности и генезис флоры Сибири: Предабайкалье и Забайкалье. Новосибирск: Наука, 1984. 264 с.
Солодянкина С.В., Истомина Е.А., Сороковой А.А., Чепинога В.В. Моделирование потенциального ареала ветреницы байкальской (Anemone baicalensis, Ranunculaceae) в Байкальском регионе // Геогр. и прир. ресурсы. 2016. № 5. С. 92–99.
Lashchinskiy N., Chepinoga V. Class Asaro-Abietetea on its eastern limit // 27th Congress of the European Vegetation Survey. Vegetation survey 90 years after the publication of Braun-Blanquet’s textbook – new challenges and concepts. Abstracts (23–26 May, 2018, Wrocław, Poland). Wrocław, 2018. P. 127.
Иванова М.М., Казановский С.Г., Киселева А.А. Находки во флоре юго-восточного (Хамар-Дабанского) побережья оз. Байкал: реликты третичной неморальной флоры и редкие виды // Turczaninowia. 2016. Т. 19. № 3. С. 94–105.
Чепинога В.В., Протопопова М.В., Павличенко В.В., Гладких Е.М. К распространению неморальных видов растений на хребте Хамар-Дабан (Южное Прибайкалье, Восточная Сибирь) // Изв. Иркут. гос. ун-та. Серия “Биология. Экология”. 2016. Т. 17. С. 30–37.
Чепинога В.В., Лащинский Н.Н., Арбузова Г.А., Гладких Е.М. Новые и редкие виды растений с хребта Хамар-Дабан (юг Восточной Сибири) // Turczaninowia. 2019. Т. 22. № 2. С. 151–159.
Методические рекомендации по ведению Красной книги субъекта Российской Федерации. М.: Министерство природных ресурсов Российской Федерации, 2006. 20 с.
Guidelines for Using the IUCN Red List Categories and Criteria. Version 14. Prepared by the IUCN Standards and Petitions Committee. 2019. Downloadable from http://www.iucnredlist.org/documents/RedListGuidelines.pdf. Last accessed 12 September 2020.
Дополнительные материалы отсутствуют.