

Экология, 2021, № 5, стр. 335-345
VS-Cambium-Developer: новый подход к моделированию функционирования камбиальной зоны хвойных под воздействием факторов окружающей среды
Д. А. Белоусова a, *, В. В. Шишов a, b, Е. А. Бабушкина a, Е. А. Ваганов a
a Сибирский федеральный университет
660041 Красноярск, просп. Свободный, 79, Россия
b Сибирский государственный университет науки и технологий им. М.Ф. Решетнева
660004 Красноярск, просп. им. газеты “Красноярский рабочий”, 31, Россия
* E-mail: daryadarya1611@gmail.com
Поступила в редакцию 02.02.2021
После доработки 20.04.2021
Принята к публикации 21.04.2021
Аннотация
Одной из фундаментальных проблем современной экологии считается оценка реакции древесных растений на воздействия окружающей среды в условиях изменяющегося климата планеты. Эффективным инструментом решения этой проблемы являются различные имитационные модели роста годичных колец деревьев. Нами предложена новая имитационная модель функционирования камбия, которая воспроизводит процесс камбиальной активности хвойных видов древесных растений в зависимости от действия ведущих климатических факторов. В основе лежит гипотеза о наличии цитоплазматического ингибитора дифференцировки клеток, функционирование которого лимитируется температурой, влажностью и освещенностью. Новый алгоритм разработан на основе имеющегося камбиального блока имитационной модели роста годичных колец Ваганова-Шашкина (VS-модели). Модель протестирована на косвенных наблюдениях функционирования камбия хвойных древесных растений в Южной Сибири (Республика Хакассия), а именно на измеренной сезонной клеточной продукции с 1964 г. по 2012 г. Предложена программная реализация новой камбиальной модели на базе технологии R Shiny, которая может быть просто адаптирована к on-line платформе VS-модели. Разработанные виджеты (средства визуализации процесса) позволяют отслеживать динамику роста камбиальной зоны с точностью до сотых долей суток.
Более 60% фитомассы древесной растительности сосредоточено в стволовой древесине, которая является результатом периодической активности латеральной меристемы – камбия [1, 2]. Поэтому создание модели, количественно описывающей его функционирование, – это не только фундаментальная задача и интегрирование нашего знания об этой ткани, но и прикладная задача, позволяющая определить инструменты управления камбиальной активностью [3–10]. В большом количестве работ [11–16] приводятся результаты, описывающие влияние на активность камбия и последующую дифференцировку его производных внутренних (гормоны роста, ассимиляты, гены, активирующие или ингибирующие процессы роста и дифференцировки) и внешних (температура, водный баланс, освещенность и др.) факторов. Сложный характер влияния факторов на активность камбия нашел отражение и в различии подходов для его количественного описания [8, 14, 17–31]. В первом приближении подходы разделяются на статистические (регрессионные модели) [21–23] и процессные (в российской терминологии – имитационные модели) [6, 18, 28, 29]. К последним, например, относится модель, в которой переход клетки в фазу растяжения лимитируется гормональным балансом и доступностью органических соединений [19, 20].
Одна из наиболее точных и одновременно сложных моделей учитывает лимитирование роста камбиальных клеток индолилуксусной кислотой (гормон роста из группы ауксинов) [24, 25]. При этом моделируется продольный перенос ауксина в пределах одного клеточного ряда. Модель учитывает конкуренцию клеток флоэмы и ксилемы, но тип деления выбирается случайным образом. Готовность клетки к делению определяется ее критическим размером [24]. Модель была успешно применена для визуального 3D-моделирования годичного кольца [25]. Главная особенность этой модели – теоретически формальное описание камбия, что позволяет моделировать повреждения ствола (атака насекомых, отсечение веток, изгиб ствола и т.д.). При этом предполагается, что моделируемые деревья растут в благоприятных внешних условиях, когда термический и водный режимы постоянны.
Новый камбиальный блок, интегрированный в модель CaBala, позволяет прогнозировать размеры трахеид, толщину клеточной стенки, угол микроволокон в клетке на основе показателей активности камбия и данных об углеводном балансе [28]. На основании этих расчетов в модели оценивается плотность древесины. Развитие камбиальных клеток лимитируется ежедневным осмотическим потенциалом роста – этот параметр связывает тургор клеточных стенок, водный и углеводный балансы. Рост каждой клетки моделируется от момента ее появления в ряду до наступления клеточной смерти (апоптоза). Модель различает радиальное и продольное растяжение клетки.
Модель XyDyS (Xylogenesis Dynamics Simulators) включает в себя ядро, функционально описывающее деление и растяжение клеток, транспорт морфогенов и два механизма для описания транспорта и граничных условий [18]. Деление и рост клеток контролируются морфогенами: ауксин, TDIF. Реакция камбиальных клеток на поступление морфогенов в ткань зависит от интенсивности и характера переноса морфогенов в ткани. Изменения в зональности морфогенетического градиента контролируются камбиальной зоной. Модель недостаточно точно рассчитывает конечные размеры клеток, так как градиент определяет зонирование независимо от роста каждой отдельной клетки [18].
“Turgor-driven growth model” дает оценку динамики роста всех тканей дерева [26]. В этой модели активность тканей лимитируется водным балансом и зависит от роста клеток, который в свою очередь лимитируется водным и температурным балансами и зависит от тургора клеточных стенок. Высокая сложность модели и высокие требования к качеству и количеству входных данных и параметров существенно затрудняют ее использование в задачах прогноза роста древесных растений в различных местообитаниях [26].
В модели, основанной на лимитировании камбиальной активности температурным и водным балансами, зависимость роста клеток камбиальной зоны от водного баланса и тургора клеточной стенки выражена через уравнение Локхарта [27]. Ферментная активность и скорость химических реакций в камбиальной зоне лимитируются температурным режимом, и при достижении пороговой температуры скорость реакций снижается за счет денатурации ферментов [27].
Все рассмотренные модели являются многомерными по параметрам и описывают, как правило, несколько процессов, происходящих как в стволовой древесине, так и в окружающей среде. Именно большое количество параметров в каждой модели и их адекватная оценка являются самой большой проблемой при попытках их использования. Потребность в создании простой и точной количественной модели функционирования камбия становится все более очевидной, поскольку значительно увеличивается физический объем измерений сезонной активности камбия и в связи с настоятельной необходимостью оценки баланса внутренних и внешних факторов в продукции новых клеток ксилемы [10, 16, 29, 32].
Новый подход к численному описанию кинетики камбия, предложенный в данной работе, основан на нескольких гипотезах о сочетании внешнего воздействия физических и биологических стрессовых факторов роста и позиционного контроля кинетики в зависимости от доступности цитоплазматического ингибитора [29], а именно:
1. Главной мишенью внешнего воздействия является камбиальная зона – зона активно делящихся клеток. Внешнее воздействие изменяет скорость роста камбиальных клеток и доступность цитоплазматического ингибитора для клеток камбиальной зоны.
2. Основные внешние факторы, лимитирующие скорость роста клеток в камбиальной зоне, – температура воздуха, содержание влаги в почве и освещенность. При расчете скорости роста клеток применяется принцип лимитирующего фактора: скорость роста определяется минимальной на данный момент частной скоростью роста, рассчитанной для каждого из факторов отдельно.
3. Доступность цитоплазматического ингибитора в клетках камбиальной зоны предопределяет объем сезонной клеточной продукции.
4. Моделируются только изменения в скорости роста клеток и их структуре, обусловленные текущими погодными условиями сезона роста (климатически обусловленные изменения прироста и структуры годичных колец) [29].
Существующий камбиальный блок модели Ваганова-Шашкина не учитывает влияние внутренних факторов на рост камбиальной зоны. Каждая клетка в камбиальном блоке характеризуется двумя переменными: ее положением в радиальном ряду и диаметром. Скорость роста клетки камбиальной зоны зависит только от скорости роста, рассчитанной в основном блоке модели Ваганова-Шашкина, которая в свою очередь зависит лишь от текущих погодных условий. Возможность деления для клетки определяется исключительно ее скоростью роста. Клетки камбия, сохранившие способность к делению в конце одного сезона, могут влиять на динамику камбия в следующем сезоне [6, 29].
Нами на основании существующего камбиального блока VS-модели [16, 29, 33] предлагается переход от моделирования роста только под влиянием внешних факторов к созданию имитационной модели, частично учитывающей основные механизмы регуляции роста камбия и продуцирование новых клеток ксилемы.
Таким образом, целью исследования является создание адекватной численной и экспериментально проверяемой модели функционирования камбия на примере функционирования камбия хвойных. Калибровка и верификация работы новой модели осуществлены на экспериментальных дендроэкологических данных лесостепной зоны юга Сибири. Усовершенствованная структура камбиального блока модели Ваганова-Шашкина позволяет минимизировать число модельных параметров, хорошо интерпретируемых как с точки зрения физиология древесных растений, так и лесной экологии для улучшения понимания функционирования камбия под воздействием различных факторов окружающей среды и более широкого охвата видов древесных растений и условий их произрастания.
МАТЕРИАЛ И МЕТОДЫ
На основе существующего камбиального блока модели Ваганова-Шашкина [29] предлагается усовершенствованная версия, в которой учитывается концентрация ингибитора роста в каждой камбиальной клетке. Отметим, что данная концентрация зависит от позиции материнской камбиальной клетки относительно инициали и меняется нелинейно в зависимости от действия климатических факторов, лимитирующих рост древесных растений.
Нами предложен простой алгоритм по расчету изменения содержания ингибитора в клетках в ответ на стрессовое воздействие климатических факторов. Прогнозирование и учет стрессовых климатических условий возможны благодаря связи новой камбиальной модели с результатами вычислений модели Ваганова-Шашкина, а именно с суточными интегральными скоростями роста [6, 29, 33]. Эта связь описывает влияние климатических условий на концентрацию биологического ингибитора роста клеток. Природа ингибитора роста в работе не уточняется. Ингибитором может быть и баланс ростовых гормонов, и доступность ассимилятов для биосинтеза новых клеток, и ограничения по скоростям биосинтеза [6, 11–16].
Основные определения
Камбий рассматривается как слой клеток, способных к неограниченному числу делений и дифференцировке [29]. В течение сезона роста камбий образует несколько слоев ксилемных и флоэмных материнских клеток (рис. 1), количество которых определяется внешними климатическими условиями, доступностью питательных веществ и контролируется гормонами. Результатом этих процессов является индивидуальная структура годичного слоя, соответствующая условиям того или иного сезона роста.
Рис. 1.
Фрагмент поперечного среза стволика сосны (а) и схема, иллюстрирующая продукцию клеток камбиальной зоной (б) (иллюстрация из работы [29]), где С – инициаль, М – материнские клетки ксилемы и флоэмы, Х – зрелые клетки ксилемы, Р – зрелые клетки флоэмы, Е – клетки в фазе растяжения.
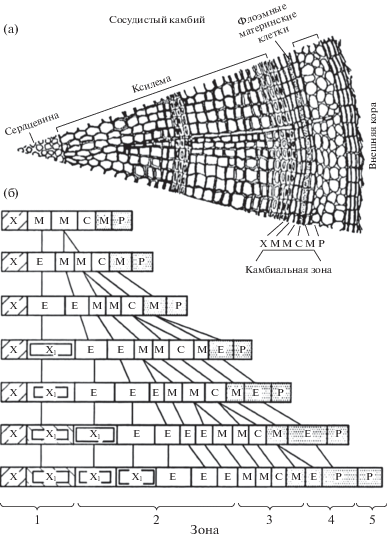
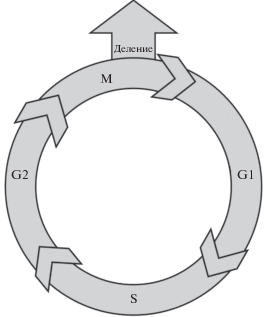
Фундаментом для понимания функционирования камбия, а именно камбиальной активности в течение сезона роста древесных растений, является клеточный цикл каждой клетки, способной к делению. Одной из основных характеристик делений клеток камбиальной зоны служит продолжительность клеточного цикла, равная промежутку времени между двумя последовательными делениями. В клеточном цикле клетка проходит четыре фазы: пресинтетическую – G1, фазу синтеза – S, предмитотическую – G2, митоз – M (рис. 2).
Рис. 2.
Диаграмма клеточного цикла. В своем развитии клетка проходит четыре фазы: пресинтетическую ‒ G1, фазу синтеза – S, предмитотическую – G2, митоз – M [29]. В результате деления образуются материнская и дочерняя клетки.
Алгоритм модели VS-Cambium-Developer. Деление инициали и материнских клеток
Модель VS-Cambium-Developer (VS-CD) воспроизводит динамику камбия для одного радиального ряда за один сезон роста (рис. 3). Длина сезона роста определяется пользователем по результатам прямых наблюдений за ксилогенезом хвойных или оценивается на основе расчетов имитационной модели роста годичных колец Ваганова-Шашкина. Периоды влияния стрессовых климатических факторов в течение сезона оцениваются VS-CD моделью на основе ежедневных интегральных скоростей роста модели Ваганова-Шашкина [5, 8, 30, 31, 34, 35]. Интегральная скорость роста определяется минимальной на данный момент частной скоростью роста, рассчитанной отдельно для каждого из лимитирующих факторов (температура воздуха, содержание влаги в почве и освещенность) [8, 29].
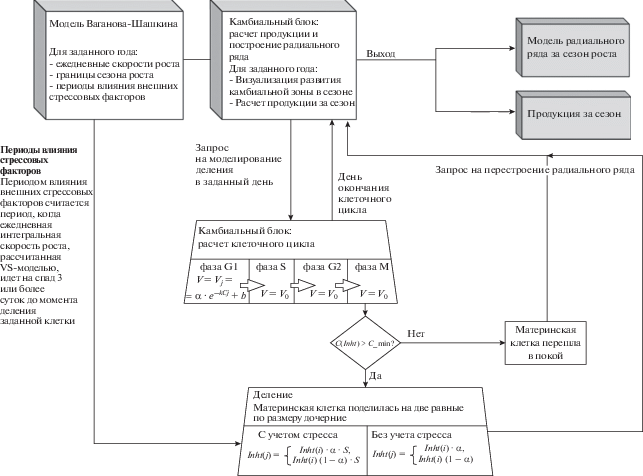
Влияние стрессовых климатических факторов учитывается при расчете клеточного цикла для каждой из материнских клеток в ряду (см. рис. 3). Для заданной материнской клетки периодом влияния внешних стрессовых (климатических) факторов считается интервал времени, когда ежедневная интегральная скорость роста последовательно уменьшается в течение 3 суток и более до момента деления самой клетки. Срок расчета стрессового периода был определен эмпирически в ходе тестирования модели на образцах сосны, собранных в лесостепи Южной Сибири. На основе экспериментов было установлено, что срок в 3 дня (срок варьировался от 1 до 6 дней) является достаточным для проявления изменений, вызванных стрессовыми условиями. Таким образом, период в 3 дня был выбран для дальнейшего моделирования сезонной продукции.
Часть входных параметров пользователь оценивает на основе визуальной параметризации, часть входных данных в VS-CD модель загружается из файла результатов расчетов модели Ваганова-Шашкина (табл. 1). Все визуальные построения новая модель производит на основании выходного массива данных, в котором хранятся сведения о каждом из делений, произошедшем за сезон (табл. 2).
Таблица 1.
Описание входных данных и параметров VS-CD модели
| Входные данные и параметры | Размерность |
|---|---|
| Параметры и данные модели Ваганова-Шашкина | |
| Длина сезона роста | сут |
| Ежедневные интегральные скорости роста | отн. ед. |
| Постоянные параметры VS-CD модели, не меняющиеся от сезона к сезону | |
| Критический диаметр клетки, при котором она переходит в фазу S из фазы G1 | мкм |
| Критический диаметр клетки, при котором она переходит в фазу G2 из фазы S | мкм |
| Критический диаметр клетки, при котором она переходит в фазу M из фазы G2 | мкм |
| Критический диаметр клетки, при котором произойдет деление | мкм |
| Минимальная концентрация ингибитора, при которой может произойти деление | отн. ед. |
| Скорость роста V0 | мкм/сут |
| Количество ингибитора в инициали | отн. ед. |
| Диаметр инициали | мкм |
| Коэффициент распределения ингибитора между дочерними клетками | отн. ед. |
| Количество ингибитора, которое передается дочерней клетке в стрессовых условиях | отн. ед. |
| Параметры VS-CD модели, характеризующие скорость роста клеток в отдельно взятом сезоне | |
| Параметр a | мкм/сут |
| Параметр b | мкм/сут |
| Параметр k | отн. ед. |
Таблица 2.
Описание выходных данных (сведения о каждом из делений, произошедшем в сезоне) VS-CD модели
| Выходные данные | Размерность |
|---|---|
| Дата появления в сезоне | сут (точность 0.001) |
| Срок прохождения материнской клеткой фазы G1 | сут (точность 0.001) |
| Срок прохождения материнской клеткой фазы S | сут (точность 0.001) |
| Срок прохождения материнской клеткой фазы G2 | сут (точность 0.001) |
| Срок прохождения материнской клеткой фазы M | сут (точность 0.001) |
| Срок прохождения материнской клеткой клеточного цикла | сут (точность 0.001) |
| Количество ингибитора, полученное дочерними клетками | отн. ед. |
| Скорость роста клетки в фазе G1 | мкм/сут |
| Концентрация ингибитора в фазе М (необходима для оценки возможности дальнейшего деления для клетки) | отн. ед. |
| Собственный уникальный идентификатор клетки | строка |
| Идентификатор материнской клетки | строка |
Алгоритм начинает расчеты с деления инициали. В течение сезона роста возможно неоднократное деление инициали [6]. Инициаль и материнские клетки делятся по одинаковым правилам. В результате деления образуются две равные по размеру клетки, в 2 раза меньше размера поделившейся клетки. Возможность нового деления любой клетки в ряду определяется только количеством ингибитора в ней. Инициаль занимает нулевую позицию в ряду, материнские клетки – позиции от 1 и далее.
В начальный момент моделирования в ряду имеется только одна инициаль, находящаяся в состоянии покоя. Пользователь задает ее размер в микронах: размер инициали возможен от 3 до 15 мкм [29, 36]. VS-CD модель рассчитывает сроки и скорости всех этапов клеточного цикла для инициали. Критерием перехода клетки из одной фазы в другую является ее размер (критические размеры переходов в клеточном цикле задаются пользователем в интервале от 3 до 15 мкм до начала работы программы). Модель рассчитывает как периоды прохождения каждой фазы отдельно, так и суммарный период прохождения клеточного цикла для каждой клетки. После прохождения клеточного цикла клетка делится на две равные по размерам.
Рост камбиальной зоны контролируется внешними климатическими условиями, которые оказывают непосредственное влияние на ежедневные интегральные скорости роста, рассчитанные VS-моделью. Количество ингибитора, получаемое дочерней клеткой, снижается в S раз (от “stress decreasing coefficient”) в периоды влияния внешних стрессовых факторов (холод, засуха и др.). Коэффициент S задается пользователем до начала работы алгоритма как параметр модели и является относительной величиной. Коэффициент может быть установлен в пределах от 0 до 1 ($0 < S < 1$); в оптимальных для роста условиях S = 1.
Инициаль в состоянии покоя хранит в себе запас ингибитора роста. Его количество задается пользователем вручную. При каждом делении ингибитор распределяется между дочерними клетками неравномерно. Пропорции ингибитора задаются пользователем и сохраняются для всех делений в пределах одного сезона. Количество ингибитора, полученное дочерними клетками от материнской или инициали, рассчитывается по формуле
(1)
$Inht\left( j \right) = ~\left\{ {\begin{array}{*{20}{c}} {Inht\left( i \right)\alpha S,\,\,\,\,\,\,\,\,\,} \\ {Inht\left( i \right)\left( {1 - \alpha } \right)S,} \end{array}} \right.$Скорость роста клетки (мкм/сут) в фазе G1 зависит от концентрации в ней ингибитора роста:
где j – номер клетки в радиальном ряду, a, b и k – параметры модели, Сj – концентрация ингибитора (относительная величина). Со скоростью Vj клетка растет только в фазе G1, в остальных фазах скорость роста постоянна и равна ${{V}_{0}}$ (скорость ${{V}_{0}}$ задается пользователем вручную от 0 до 1 до начала работы программы). В период, пока клетка проходит каждую из фаз клеточного цикла, скорость, заданная в начале фазы, не изменяется.Формула расчета концентрации ингибитора в клетке (относительная величина) имеет следующий вид:
где j – номер клетки в радиальном ряду, Сj – концентрация ингибитора (относительная величина), Dj – радиальный размер клетки (мкм), Inht(j) – количество ингибитора, полученное j-й клеткой от материнской.Каждая материнская клетка в ряду характеризуется количеством ингибитора в ней и моментом времени появления ее в сезоне. Клетка считается готовой к делению, если соблюдены два условия:
1. Концентрация ингибитора (см. формулу (3)) на момент фазы митоза больше критической (критическая концентрация задается пользователем вручную до старта программы в пределах от 0 до 1).
2. Время появления в сезоне и период прохождения клеткой клеточного цикла в сумме не превышают длину сезона роста.
Невыполнение хотя бы одного из этих условий приводит к тому, что клетка теряет способность делиться и переходит в зону растяжения.
Алгоритм расчета фаз деления материнской клетки аналогичен алгоритму для инициали.
Косвенные измерения камбиальной активности и верификация новой модели
Для верификации работы новой модели использованы образцы сосны обыкновенной (Pinus sylvestris), собранные на участке, лимитируемом влажностью почвы и температурным режимом (деревья чувствительны к засухе и холодовому стрессу) в лесостепи южной Сибири (53°43′ с.ш., 91°47′ в.д., 251 м над ур. м.) на склоне холма южной экспозиции [17]. Данные по ежегодной клеточной продукции не стандартизованы [36], моделируется реальная сезонная клеточная продукция.
Керны были извлечены из 20 зрелых неповрежденных деревьев P. sylvestris. Сбор, транспортировку, обработку кернов, измерение (с точностью 0.01 мм), кросс-датировку по ширине годичных колец проводили по стандартным методикам дендрохронологии [37] c помощью установки LINTAB и программ TSAP (Rinntech, Heidelberg, Germany) и COFECHA [38]. По фотографиям высокого разрешения срезов, полученных на микротоме по древесным образцам, измеряли следующие характеристики колец за 1969 – 2008 гг.: количество клеток, их радиальные размеры, толщина клеточной стенки для 5 рядов с последующим усреднением при помощи программного обеспечения ACR [39]. Для верификации модели необходимы данные только о сезонной кумулятивной клеточной продукции (количестве клеток за сезон).
Для моделирования индексов ежегодного прироста с помощью VS-осциллографа [8] использованы данные об изменчивости климатических переменных ближайшей метеорологической станции Минусинск (в 25 км) (53°41′ с.ш., 91°40′ в.д., 254 м над ур. м.). Период калибровки – 1969–1985 гг. (коэффициент корреляции – 0.69, синхронности – 0.88), период верификации – 1983–2008 гг. (коэффициент корреляции – 0.44, синхронности – 0.67). Источник климатических данных – интерактивная база данных ресурса meteo.ru.
Для моделирования камбиальной активности было выбрано “типичное” древесное растение, клеточная продукция которого наиболее близка к средней клеточной продукции на периоде с 1969 г. по 2008 г. для анализируемого древостоя (участка) (рис. 4). Для выбранного образца смоделированы благоприятные сезоны роста – 1985, 1986, 1993 гг. и неблагоприятные – 1995, 2001, 2002 гг.
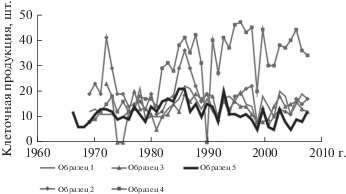
Средства визуализации результатов моделирования и параметризации в VS-CD модели
Визуальный способ ручной параметризации VS-CD модели выполнен с помощью интерфейсного элемента управления “бегунок” (см. Приложение 1 ). Каждому из параметров модели соответствует численный промежуток от минимально возможного до максимального значений. Любое изменение значения любого параметра модели, т.е. уменьшение (сдвиг бегунка влево) или увеличение (сдвиг бегунка вправо) его значения, приводит к моментальному пересчету суточной клеточной продукции для всего сезона роста и перестроению диаграмм визуализации развития камбиальной зоны. Параметры модели выбираются таким образом, чтобы минимизировать расхождение между наблюденной суточной клеточной продукцией и модельными результатами.
Программа, реализующая алгоритм VS-CD модели, позволяет представить информацию обо всех делениях за сезон в виде дерева делений, в узлах которого отображены 3 главные характеристики каждого из делений:
1. Период прохождения материнской клеткой клеточного цикла.
2. Концентрация ингибитора в клетке на момент фазы М.
3. Скорость роста клетки в фазе G1.
Алгоритм реализован с помощью языка программирования R с использованием пакета Shiny [40] и размещен по адресу: http://www.vs-genn.ru/VSCD11. Внешний вид главной страницы веб-сайта представлен в Приложении 2 .
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Моделирование выполнено с помощью визуального способа параметризации – “Бегунки” (см. Приложение 1 ). Параметры, характеризующие рост дерева и не изменяющиеся от сезона к сезону, приведены в табл. 3, параметры модели, характеризующие скорость роста клеток и результаты моделирования, – в табл. 4. Отметим, что результаты моделирования по клеточной сезонной продукции отличаются от реальной продукции не более чем на 1 клетку. Адекватность моделирования косвенно подтверждается также периодом (длительностью) первого деления инициали. Для Южной Сибири средний срок появления первой дочерней клетки в сезоне роста равен 20–25 суткам [29, 36].
Таблица 3.
Параметры модели, характеризующие рост дерева и не изменяющиеся от сезона к сезону, заданные для моделирования типичного дерева сосны обыкновенной (Pinus sylvestris)
| Параметр | Значение |
|---|---|
| Критический диаметр клетки, при котором она переходит в фазу S из фазы G1, мкм | 8.0 |
| Критический диаметр клетки, при котором она переходит в фазу G2 из фазы S, мкм | 8.2 |
| Критический диаметр клетки, при котором она переходит в фазу M из фазы G2, мкм | 8.5 |
| Критический диаметр клетки, при котором произойдет деление, мкм | 9.0 |
| Минимальная концентрация ингибитора, при которой может произойти деление, отн. ед. | 0.018 |
| Скорость роста V0, мкм/сут | 1.0 |
| Количество ингибитора в инициали, отн. ед. | 4.0 |
| Диаметр инициали, мкм | 5.0 |
| Коэффициент распределения ингибитора между дочерними клетками | 0.65 |
| Количество ингибитора, которое передается дочерней клетке в стрессовых условиях | 0.8 |
Таблица 4.
Параметры модели, характеризующие скорость роста клеток, и результаты моделирования (сравнение наблюденной и модельной продукций) роста камбиальной зоны среднерастущего дерева сосны обыкновенной (Pinus sylvestris) в лесостепи южной Сибири (деревья на участке чувствительны к холодовому стрессу и засухе)
| Год | Параметр | Реальная продукция | Модельная продукция | Срок первого деления инициали, сут | ||
|---|---|---|---|---|---|---|
| a, мкм/сут | b, мкм/сут | k, отн. ед. | ||||
| Благоприятные сезоны роста | ||||||
| 1985 | 0.124 | 0.115 | 3.35 | 21 | 22 | 25.29 |
| 1986 | 0.122 | 0.111 | 3.18 | 21 | 21 | 25.87 |
| 1993 | 0.12 | 0.13 | 3.35 | 20 | 21 | 22.70 |
| Неблагоприятные сезоны роста | ||||||
| 1995 | 0.08 | 0.12 | 4.3 | 11 | 10 | 25.47 |
| 2001 | 0.06 | 0.11 | 4.35 | 6 | 6 | 27.82 |
| 2002 | 0.06 | 0.11 | 4.35 | 5 | 5 | 27.82 |
Результаты тестирования модели согласуются с наблюдениями за продукцией ксилемы реальных деревьев, чувствительных к стрессовым климатическим условиям. Наше исследование показывает, что низкая или высокая продукция клеток ксилемы может быть ассоциирована с разными скоростями роста клеток камбиальной зоны, доступностью ингибитора клеточного роста и стрессовыми условиями среды. В благоприятные периоды роста скорость роста клеток в фазе клеточного цикла G1 практически в 2 раза выше, чем в неблагоприятные, а количество получаемого дочерними клетками ингибитора в благоприятные периоды выше, чем в неблагоприятные.
Возможной причиной отклонения моделирования клеточной продукции от реальной на 1 клетку является отсутствие учета конкуренции клеток ксилемы и флоэмы при делении камбиальных клеток, а также отсутствие учета уровня стрессовых гормонов.
Разработанная модель является постепенным переходом от процессной модели к модели, частично учитывающей основные механизмы, регулирующие активность камбия и продуцирование новых клеток ксилемы. Для дальнейшего усовершенствования работы модели и ее верификации требуются новые прямые и косвенные источники информации о ксилогенезе хвойных, охватывающие различные условия местообитаний древесных растений.
Работа выполнена в рамках проекта РФФИ № 19-04-00274 A и государственного задания в сфере науки СибГУ им. М.Ф. Решетнева (проект FEFE 2020-00104).
Авторы статьи подтверждают отсутствие конфликта интересов. В настоящей работе не содержится исследований с участием людей или животных в качестве объектов исследования.
Список литературы
Чавчавадзе Е.С. Древесина хвойных. Морфологические особенности, диагностическое значение. Л.: Наука, 1979. 190 с.
Larson P.R. The vascular cambium. Development and structure. Berlin: Springer-Verlag, 1994. 725 p.
Бабушкина Е.А., Белокопытова Л.В. Камбиальная зона – основная мишень влияния внешних факторов на формирование годичных колец хвойных // Изв. ВУЗов. Лесной журнал. 2015. № 6 (348). С. 35–45.
Bhalerao R.P., Fischer U. Environmental and hormonal control of cambial stem cell dynamics. // J. of Experimental Botany. 2017. V. 68. Issue 1. P. 79–87. https://doi.org/10.1093/jxb/erw466
Buttò V., Deslauriers A., Rossi S. et al. The role of plant hormones in tree-ring formation // Trees. 2019. V. 34. P. 315–335. https://doi.org/10.1007/s00468-019-01940-4
Anchukaitis K.J., Evans M.N., Hughes M.K., Vaganov E. An interpreted language implementation of the Vaganov-Shashkin tree-ring proxy system model // Dendrochronologia. 2020. V. 60. P. 125677. https://doi.org/10.1016/j.dendro.2020.125677
He M., Yang B., Bräuning A. et al. Recent advances in dendroclimatology in China // Earth-Science Reviews. 2019. V. 194. P. 521–535. https://doi.org/10.1016/j.earscirev.2019.02.012
Tychkov I.I., Sviderskaya I.V., Babushkina E.A et al. How can the parameterization of a process-based model help us understand real tree-ring growth? // Trees. 2019. V. 33. № 2. P. 345–357. https://doi.org/10.1007/s00468-018-1780-2
Arzac A., Llambí L.D., Dulhoste R. et al. Modelling the effect of temperature changes on plant life-form distribution across a treeline ecotone in the tropical Andes // Plant Ecology & Diversity. 2019. V. 12. №. 6. P. 619–631. https://doi.org/10.1080/17550874.2019.1655108
Popkova M.I., Shishov V.V., Vaganov E.A. et al. Contribution of xylem anatomy to tree-ring width of two larch species in permafrost and non-permafrost zones of siberia // Forests. 2020. V. 11. P.1343. https://doi.org/10.3390/f11121343
Savidge R.A. Xylogenesis, genetic and environmental regulation – a review // Iawa Journal. 1996. V. 17. № 3. P. 269–310. https://doi.org/10.1163/22941932-90001580
Uggla C., Mellerowicz E.J., Sundberg B. Indole-3-acetic acid controls cambial growth in Scots pine by positional signalling // Plant Physiology. 1998. V. 117. P. 113–121. https://dx.doi.org/10.1104%2Fpp.117.1.113
Uggla C., Magel E., Moritz T., Sundberg B. Function and dynamics of auxin and carbohydrates during earlywood/latewood transition in Scots pine // Plant Physiology. 2001. V. 125. P. 2029–2039. https://doi.org/10.1104/pp.125.4.2029
Rossi S., Deslauriers A., Anfodillo T. et al. Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length // New Phytologist. 2006. V. 170. P. 301–310. https://doi.org/10.1111/j.1469-8137.2006.01660.x
Chuine I. Why does phenology drive species distribution? // Philosophical Transactions of the Royal Society B: Biological Sciences. 2010. V. 365. № 1555. P. 3149–3160. https://doi.org/10.1098/rstb.2010.0142
Rossi S., Anfodillo T., Čufar K. et al. A meta-analysis of cambium phenology and growth: linear and non-linear patterns in conifers of the northern hemisphere // Annals of Botany. 2013. V. 112. № 9. P. 1911–1920. https://doi.org/10.1093/aob/mct243
Popkova M.I., Vaganov E.A., Shishov V.V. et al. Modeled tracheidograms disclose drought influence on Pinus sylvestris tree-rings structure from siberian forest-steppe // Frontiers in Plant Science. 2018. V. 9. P. 1144. https://doi.org/10.3389/fpls.2018.01144
Hartmann F. P., Rathgeber C.B.K., Fournier M., Moulia B. Modelling wood formation and structure: power and limits of a morphogenetic gradient in controlling xylem cell proliferation and growth // Annals of Forest Science. 2017. V. 74. № 1. P. 1–15. https://doi.org/10.1007/s13595-016-0613-y
Deleuze C., Houllier F. Connection between silviculture and wood quality through modelling approaches and simulation softwares // IAWA Journal. 1996. V.17. № 2. P. 105–112. https://doi.org/10.1163/22941932-90001438
Deleuze C., Houllier F. A simple process-based xylem growth model for describing wood microdensitometric profiles // J. of Theoretical Biology. 1998. V. 193. № 1. P. 99–113. https://doi.org/10.1006/jtbi.1998.0689
Cuny H.E., Rathgeber C.B.K., Lebourgeois F. et al. Life strategies in intra-annual dynamics of wood formation: example of three conifer species in a temperate forest in north-east France // Tree Physiology. 2012. V. 32. № 5. P. 612–625. https://doi.org/10.1093/treephys/tps039
Cuny H.E., Rathgeber C.B.K., Kiessé T.S. et al. Generalized additive models reveal the intrinsic complexity of wood formation dynamics // J. of Experimental Botany. 2013. V. 64. № 7. P. 1983–1994. https://doi.org/10.1093/jxb/ert057
Rossi S., Deslauriers A., Anfodillo T. et al. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes // Oecologia. 2007. V. 152. P. 1–12. https://doi.org/10.1007/s00442-006-0625-7
Forest L., Demongeot J. Cellular modelling of secondary radial growth in conifer trees: application to Pinus radiata (D. Don) // Bull. Math. Biol. 2006. V. 68. № 4. P. 753–784. https://doi.org/10.1007/s11538-005-9004-5
Forest L., Martín J.S., Padilla F. et al. Morphogenetic processes: application to cambial growth dynamics // Acta Biotheoretica. 2004. V. 52. № 4. P. 415–438. https://doi.org/10.1023/B:ACBI.0000046607.17817.20
Peters R.L., Steppe K., Cuny H.E. et al. Turgor – a limiting factor for radial growth in mature conifers along an elevational gradient // New Phytologist. 2021. V. 229. №. 1. P. 213–229. https://doi.org/10.1111/nph.16872
Cabon A., Fernández-de-Uña L., Gea-Izquierdo G. et al. Water potential control of turgor-driven tracheid enlargement in Scots pine at its xeric distribution edge // New Phytologist. 2020. V. 225. № 1. P. 209–221. https://doi.org/10.1111/nph.16146
Drew D.M., Downes G. A model of stem growth and wood formation in Pinus radiata // Trees. 2015. V. 29. № 5. P. 1395–1413. https://doi.org/10.1007/s00468-015-1216-1
Vaganov E.A., Hughes M.K., Shashkin A.V. Growth dynamics of conifer tree rings: images of past and future environments. Berlin: Springer-Verlag, 2006. 358 p.
Shishov V.V., Tychkov I.I., Popkova M.I. et al. VS-oscilloscope: A new tool to parameterize tree radial growth based on climate conditions // Dendrochronologia. 2016. V. 39. P. 42–50. https://doi.org/10.1016/j.dendro.2015.10.001
Tumajer J., Kašpar J., Kuželová H. et al. Forward modeling reveals multidecadal trends in cambial kinetics and phenology at treeline // Frontiers in Plant Science. 2021. V. 12. P.613643. https://doi.org/10.3389/fpls.2021.613643
Nanayakkara B., Dickson A.R., Meason D.F. Xylogenesis of Pinus radiata D. Don growing in New Zealand // Annals of Forest Science. 2019. V. 76. № 3. P. 74. https://doi.org/10.1007/s13595-019-0859-2
Vaganov E.A., Anchukaitis K.J., Evans M.N. How well understood are the processes that create dendroclimatic records? A mechanistic model of the climatic control on conifer tree-ring growth dynamics // Dendroclimatology. 2011. V. 11. P. 37‒75. https://doi.org/10.1007/978-1-4020-5725-0_3
He M., Shishov V., Kaparova N. et al. Process-based modeling of tree-ring formation and its relationships with climate on the Tibetan Plateau // Dendrochronologia. 2017. V. 42. P. 31–41. https://doi.org/10.1016/j.dendro.2017.01.002
Jevšenak J., Tychkov I., Gričar J. et al. Growth-limiting factors and climate response variability in Norway spruce (Picea abies L.) along an elevation and precipitation gradients in Slovenia // International J. of Biometeorology. 2021. V. 65. №. 2. P. 311–324. https://doi.org/10.1007/s00484-020-02033-5
Ваганов Е.А., Шашкин А.В., Свидерская И.В., Высоцкая Л.Г. Гистометрический анализ роста древесных растений. Новосибирск: Наука, 1985. 100 с.
Шиятов С.Г., Ваганов Е.А., Кирдянов А.В. и др. Методы дендрохронологии. Ч. I. Основы дендрохронологии. Сбор и получение древесно-кольцевой информации. Красноярск: КрасГУ, 2000. 80 с.
Grissino-Mayer H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA // Tree-Ring Research. 2001. V. 57. № 2. P. 205–221.
Dyachuk P., Arzac A., Peresunko P. et al. AutoCellRow (ACR) – A new tool for the automatic quantification of cell radial files in conifer images // Dendrochronologia. 2020. V. 60. P. 125687. https://doi.org/10.1016/j.dendro.2020.125687
Chang W., Cheng J., Allaire J. et al. Shiny: web application framework for R // R package version. 2017. V. 1. №. 5. P. 2017.
Дополнительные материалы отсутствуют.