

Экология, 2022, № 2, стр. 87-95
Влияние водных вытяжек из листьев клена ясенелистного и листьев местных видов деревьев на раннее развитие растений
Д. В. Веселкин a, *, О. С. Рафикова a
a Институт экологии растений и животных УрО РАН
620144 Екатеринбург, ул. 8 Марта, 202, Россия
* E-mail: veselkin_dv@ipae.uran.ru
Поступила в редакцию 21.10.2021
После доработки 01.11.2021
Принята к публикации 03.11.2021
- EDN: AVMAXS
- DOI: 10.31857/S0367059722020081
Аннотация
Целью работы была проверка предположения об аллелопатической активности чужеродного (инвазивного) в Евразии вида дерева Acer negundo. В двух экспериментах, выполненных методом рулонной культуры, оценили влияние водных вытяжек из летних (зеленых) и осенних (расцвеченных) листьев разных древесных растений на раннее развитие Festuca rubra, Sinapis alba и Trifolium repens. Применяли четыре варианта водных вытяжек из листьев: инвазивного Acer negundo и местных видов древесных растений Sorbus aucuparia, Prunus padus и Salix caprea. Однозначно доказать, что водные вытяжки из листьев A. negundo влияют на раннее развитие тест-растений, не удалось. Эффекты вытяжек из листьев A. negundo не отличались по силе от эффектов вытяжек из листьев местных растений Prunus padus и Salix caprea. Местный кустарник Sorbus aucuparia выраженно ингибировал раннее развитие тест-растений. В целом не получено свидетельств, что воздействия аллелопатического характера путем вымывания веществ из листьев могут быть реальным механизмом, обеспечивающим экологический успех A. negundo во вторичном ареале.
Один из вероятных механизмов экологического успеха чужеродных инвазивных растений во вторичных ареалах, посредством которого они влияют на местные растения и сообщества, – аллелопатия [1–5]. Гипотеза “нового оружия” – Novel Weapons Hypothesis, NWH [2], предполагает, что благодаря высвобождению уникальных химических соединений, к которым местные виды не приспособлены, некоторые инвазивные виды могут получать конкурентные преимущества или реализовывать средопреобразующие воздействия [2, 3]. Накоплено много подтверждений прямых аллелопатических эффектов со стороны инвазивных растений на местные растения [обзоры: 6, 7]. В мета-анализе на основе 378 публикаций [7] установлено, что аллелопатические эффекты в среднем снижали продуктивность растений на 25%. При этом влияние чужеродных видов на местные было более негативным по сравнению с влиянием местных видов на местные [7]. Однако ингибирующее аллелопатическое влияние чужеродных растений подтверждается не всегда. Например, аллелопатические эффекты не найдены в отношении следующих инвазивных видов: Euphorbia esula L. [8], Centaurea stoebe L. [8, 9], Solidago gigantea Ait. [10], Impatiens glandulifera Royle [10, 11], Erigeron annuus (L.) Pers. [10], Thymus vulgaris L. [12]. Таким образом, несмотря на преобладание результатов, подтверждающих предположение об аллелопатической активности инвазивных растений, это предположение не универсально для объяснения экологического успеха конкретных видов.
Противоречивы сведения об аллелопатии Acer negundo L., который относят к группе видов-трансформеров [13]. В отношении аллелопатической активности A. negundo опубликовано несколько подтверждений [14–17]. Однако есть также и случаи, когда наблюдаемые эффекты были неясными или отсутствовали [18–20].
Относительно мало известно о соотношении конспецифичных и гетероспецифичных (направленных соответственно на представителей своего или других видов) аллелопатических эффектов инвазивных растений [21]. Одна из гипотез, объясняющая способность чужеродных растений создавать сообщества с высоким уровнем доминирования, состоит в том, что инвайдеры могут трансформировать условия в неблагоприятную сторону для конкурирующих видов, но благоприятную для собственного развития [22]. В частности, положительное аллелопатическое влияние на прорастание собственных семян установлено для Lonicera maackii [23]. Для A. negundo показано, что его сеянцы улучшают условия роста для взрослых деревьев [24]. Для надежного суждения об аллелопатии как о механизме инвазии A. negundo необходимо накопление методически строгих результатов по проверке кон- и гетероспецифичных эффектов в экспериментах разного дизайна.
Цель настоящей работы – в экспериментах, выполненных методом рулонных культур, оценить влияние водных вытяжек из летних (зеленых) и осенних (расцвеченных) листьев A. negundo на его раннее развитие и развитие нескольких видов трав. Как наиболее вероятные на основании анализа опубликованных данных проверяли гипотезы: 1) водные вытяжки из листьев A. negundo отрицательно влияют на раннее развитие тест-растений; 2) гетероспецифичные эффекты вытяжек из листьев A. negundo более негативны по сравнению с конспецифичными эффектами.
МАТЕРИАЛ И МЕТОДЫ
При изучении аллелопатических эффектов в экспериментах с водными вытяжками рекомендуется [25–27] избегать: 1) применения в качестве единственного контроля дистиллированной воды; 2) отсутствия сравнения с воздействием местных видов; 3) измельчения растительного материала. При планировании работы мы учитывали эти требования.
Район исследования – г. Екатеринбург (56°50′ с. ш., 60°35′ в. д.), большой город с населением 1.4 млн человек, административный центр Свердловской области, расположенный в южнотаежной подзоне бореально-лесной зоны. В растительных сообществах подзоны преобладают сосновые (Pinus sylvestris L.) леса на дерново-подзолистых почвах и буроземах. Климат умеренно континентальный, зима длительная и холодная с устойчивым снежным покровом, лето короткое. Среднегодовая температура +3.0°C, средняя температура января −12.6°C, июля +19.0°C, среднегодовая сумма осадков 550–650 мм. Максимум осадков приходится на теплый сезон (май–август), в течение которого выпадает около 60–70% годовой суммы. Листья донорных растений для приготовления экстрактов собирали на территории крупного внутригородского общественного парка – Центрального парка культуры и отдыха им. В. Маяковского. Растительность парка формируют сосновые леса с примесью Betula spp., Populus tremula L., Tilia sp. В парке много декоративных посадок, в том числе интродуцентов: Ulmus laevis Pall., Malus baccata (L.) Borkh., A. negundo, Syringa vulgaris L., Cotoneaster lucidus Schltdl., Caragana arborescens Lam.
Растения-доноры. Изучали аллелопатическую активность одного инвазивного древесного вида Acer negundo и трех местных – Sorbus aucuparia L., Prunus padus L. и Salix caprea L. Все четыре вида обычны в подлеске урбанизированных лесов в районе г. Екатеринбурга и могут иметь жизненную форму кустарников, особенно S. aucuparia и P. padus, или невысоких деревьев (далее для краткости они называются деревьями или древесными растениями). Они занимают сходные местообитания и потенциально могут конкурировать друг с другом. Этим обусловлен их выбор для сравнительного исследования: A. negundo – североамериканский вид, являющийся инвазивным на большей части своего интродуцированного ареала [13, 28, 29], под его влиянием снижается разнообразие аборигенных растений [30–32]; S. aucuparia – кустарник или дерево семейства Rosaceae высотой обычно не более 7–10 м; P. padus – кустарник, иногда дерево семейства Rosaceae высотой до 10–15 м; S. caprea – невысокое (до 10 м) дерево семейства Salicaceae.
Тест-растения: Festuca rubra L. (сем. Poaceae; рыхлодерновинный многолетник); Sinapis alba L. (сем. Brassicaceae; однолетник); Trifolium repens L. (сем. Fabaceae; многолетник). Выбор видов тест-растений обусловлен следующими критериями: F. rubra и T. repens – обычные виды умеренно нарушенных местообитаний, в которые может вторгаться A. negundo; S. alba – стандартный тест-объект в исследованиях аллелопатии. Виды тест-растений – представители однодольных и двудольных, т.е. различаются таксономически. Кроме того, они образуют разные типы корневых симбиозов: S. alba – безмикоризный вид; F. rubra и T. repens образуют арбускулярную микоризу, а T. repens дополнительно вступает в симбиоз с азотфиксирующими ризобиями. Эти виды включены в настоящее исследование потому, что их реакция на преобразование условий под влиянием A. negundo ранее нами уже изучалась в эксперименте иного дизайна [19]. Все виды выращивали из покупных семян.
Сбор листьев и приготовление водных вытяжек. Свежие листья с 5 особей каждого донорного вида собирали на высоте 1.5–3 м независимо от ориентации кроны по сторонам света или внутри сообщества. Листья не измельчали, максимально сохраняя их целостность. Вытяжки готовили, заливая листья дистиллированной водой в соотношении 1 : 10 по массе [27], настаивали 24 ч в темном месте при комнатной температуре, затем фильтровали через фильтровальную бумагу. Колбы с готовыми вытяжками хранили в холодильнике при +4°C. Каждые 7 сут готовили свежие вытяжки.
Эксперимент. Для проращивания семян использовали цилиндрические прозрачные сосуды объемом 150 мл. Сосуды, полосы фильтровальной бумаги и кальки шириной 4 см и длиной 50 см предварительно 2 ч стерилизовали при 110°C. По 50 семян F. rubra, S. alba, T. repens и по 10 семян A. negundo раскладывали на полосы фильтровальной бумаги через равные промежутки на расстоянии 5–7 мм от верхнего края бумажной полосы. Поверх полосы фильтровальной бумаги размещали слой кальки и сворачивали бумагу в рулон. Один конец рулона опускали в сосуды, в которые наливали по 50 мл (треть объема сосуда) воды или водных вытяжек из листьев. Уровень жидкости в течение эксперимента поддерживали на одном уровне. Семена проращивали 21 сутки, ежедневно переставляя сосуды в случайном порядке. Подобная рандомизация позволяет избежать влияния случайных факторов. На 21-е сутки экспозиции определяли: 1) выживаемость – долю живых проростков по отношению к числу посеянных семян; 2) длину надземной части (мм) – расстояние от корневой шейки (места перехода зародышевого корешка в гипокотиль) до верхушки побега (S. alba, T. repens и A. negundo) или до верхушки самого длинного листа (F. rubra); 3) длину главного корня (мм).
Каждое сочетание параметров “вытяжка (5 вариантов) × вид тест-растения (4 варианта)” воспроизвели в 3 повторностях (сосудах). Таким образом, всего в одном эксперименте было 60 сосудов, которые экспонировали одновременно при одинаковых температуре и освещении. По этой схеме провели два эксперимента: первый – 29.07.2020–19.08.2020 г. с летними (зелеными) листьями деревьев; второй – 08.09.2020–29.09.2020 г. с осенними (расцвеченными) листьями деревьев.
Статистический анализ данных осуществляли с помощью двух- и трехфакторного дисперсионного анализа (ANOVA). Единицей наблюдения в статистическом анализе было значение признака в одном сосуде (n = 120). Признаки, измеренные в долях, предварительно подвергали арксинус-трансформации для приближения к условиям допустимости проведения ANOVA, но в таблицах и на рисунках указаны не преобразованные значения признаков. Для нивелирования видовых особенностей тест-растений использовали относительные показатели – относительную выживаемость и относительную длину корня. Их рассчитывали как отношение значения признака в данном сосуде к среднему значению соответствующего признака в трех сосудах с дистиллированной водой. Однородность дисперсий контролировали с помощью критерия Левена, а парные различия между вариантами оценивали с помощью критерия Тьюки. Чтобы оценить, какие факторы сильнее влияли на признаки раннего развития тест-растений, использовали информационный критерий Акаике (AIC). Расчеты выполнены в пакете STATISTICA 8.0 (StatSoft Inc., USA, 1984–2007). При усреднении значений через символ ± приведена стандартная ошибка (SE).
РЕЗУЛЬТАТЫ
Факторы изменчивости признаков раннего развития тест-растений. Судя по минимальным значениям AIC (табл. 1), для оптимального описания состояния тест-растений необходимо использовать все три основных предиктора: 1) вариант вытяжки – вытяжки из листьев разных растений-доноров; 2) качество листьев (летние или осенние); 3) принадлежность тест-растения к тому или иному виду. Выживаемость и морфологические характеристики различались у разных видов тест-растений и в зависимости от варианта вытяжки (табл. 2). Кроме того, выживаемость и развитие тест-растений зависели от сезона года. Статистически значимыми были также многие взаимодействия факторов. Таким образом, общая структура изменчивости признаков состояния тест-растений сложная.
Таблица 1.
Качество объяснения (значения AIC) и лучшие сочетания предикторов для объяснения изменчивости признаков раннего развития тест-растений
| Признак | AIC отдельных предикторов | Лучшее сочетание предикторов | |||
|---|---|---|---|---|---|
| вариант вытяжки | сезон | вид тест-растения | сочетание предикторов | AIC | |
| Выживаемость | 75.55 | 76.67 | –53.32 | Вариант вытяжки + сезон + вид тест-растения | –66.77 |
| Длина надземной части | 992.29 | 991.28 | 846.20 | Вариант вытяжки + сезон + вид тест-растения | 822.09 |
| Длина главного корня | 998.94 | 1033.15 | 1026.62 | Вариант вытяжки + сезон + вид тест-растения | 957.31 |
Таблица 2.
Полученные в 3-факторном ANOVA уровни значимости влияния (P) факторов “вид тест-растений”, “сезон” и “вариант вытяжки” на признаки раннего развития тест-растений
| Признак | Факторы | Взаимодействия факторов | |||||
|---|---|---|---|---|---|---|---|
| вид тест-растений (dF = 2) |
сезон (dF = 1) |
вариант вытяжки (dF = 4) |
вид тест-
растений × × сезон (dF = 2) |
вид тест-растений × × вариант вытяжки (dF = 8) |
сезон × × вариант вытяжки (dF = 4) |
вид тест-
растений × сезон × × вариант вытяжки (dF = 8) |
|
| Выживаемость | <0.0001 | 0.0137 | <0.0001 | 0.5398 | <0.0001 | 0.0029 | 0.5268 |
| Длина надземной части | <0.0001 | 0.0005 | <0.0001 | 0.0581 | 0.0002 | 0.0173 | 0.1833 |
| Длина главного корня | <0.0001 | 0.0003 | <0.0001 | 0.6163 | 0.0004 | <0.0001 | 0.0102 |
Для проверки наших рабочих гипотез важнее всего сравнение абсолютных значений признаков в зависимости от фактора “вариант вытяжки”. Поэтому далее в первую очередь обсуждаются эффекты, связанные с вариантами вытяжки, а факторы “вид тест-растений” (“группа тест-растений”) и “сезон” рассматривали как модифицирующие. В ANOVA расчеты выполняли с учетом основных факторов и их взаимодействий.
Влияние вытяжек разных растений-доноров на ранние этапы онтогенеза тест-растений. Наименьшие выживаемость и длина главного корня обычно наблюдались в вариантах с вытяжкой из листьев S. aucuparia (табл. 3), наименьшая длина надземной части – в варианте с водой или при развитии на вытяжках из листьев местных видов S. caprea и S. aucuparia. Ни выживаемость, ни морфологические параметры развития тест-растений при развитии на вытяжках из листьев Acer negundo не были минимальными по сравнению с другими вытяжками. Напротив, развитие тест-растений на вытяжках из листьев A. negundo происходило хорошими темпами – все показатели состояния тест-растений были близки к верхним значениям.
Таблица 3.
Значения признаков раннего развития (±SE) тест-растений при проращивании на разных вытяжках, усредненные для экспериментов с летними и осенними листьями
| Признак, вид тест-растения |
Вариант вытяжки | ||||
|---|---|---|---|---|---|
| вода | местные деревья | Acer negundo | |||
| Salix caprea |
Prunus padus |
Sorbus aucuparia | |||
| Выживаемость, % | |||||
| Acer negundo | 35 ± 2a | 30 ± 9ab | 10 ± 6a | 17 ± 6ab | 30 ± 4ab |
| Festuca rubra | 86 ± 2a | 83 ± 2a | 86 ± 2a | 76 ± 7a | 87 ± 3a |
| Sinapis alba | 72 ± 6a | 81 ± 5a | 85 ± 3a | 71 ± 4a | 79 ± 5a |
| Trifolium repens | 77 ± 1c | 55 ± 5b | 63 ± 3bc | 27 ± 6a | 64 ± 4bc |
| Длина надземной части, мм | |||||
| Acer negundo | 41 ± 5a | 33 ± 4a | 41 ± 6a | 46 ± 8a | 52 ± 8a |
| Festuca rubra | 58 ± 1a | 63 ± 2ab | 63 ± 2ab | 56 ± 3a | 65 ± 2b |
| Sinapis alba | 32 ± 1a | 56 ± 3c | 59 ± 3c | 47 ± 5b | 56 ± 2c |
| Trifolium repens | 13 ± 1a | 20 ± 1bc | 21 ± 1c | 17 ± 2b | 21 ± 1c |
| Длина главного корня, мм | |||||
| Acer negundo | 38 ± 13a | 18 ± 6a | 28 ± 15a | 13 ± 8a | 40 ± 12a |
| Festuca rubra | 78 ± 1d | 46 ± 4b | 56 ± 4c | 21 ± 6a | 42 ± 7b |
| Sinapis alba | 49 ± 6b | 55 ± 3b | 52 ± 6b | 25 ± 8a | 51 ± 2b |
| Trifolium repens | 70 ± 4c | 49 ± 6c | 67 ± 6b | 21 ± 6a | 51 ± 6b |
Влияние вытяжек из зеленых и расцвеченных листьев на ранние этапы онтогенеза тест-растений. Выживаемость растений, длина надземной части и главного корня летом и осенью различались в зависимости от сезона (см. табл. 1). Летом и осенью средняя относительная выживаемость при проращивании на воде, вытяжках из листьев S. caprea и A. negundo не различалась. При этом летом вытяжки из листьев S. aucuparia заметно больше снижали относительную выживаемость тест-растений, чем вытяжки из S. caprea и A. negundo (по критерию Тьюки на уровне минимум P < 0.008; рис. 1а). Вместе с тем различия между ингибирующей способностью вытяжек из летних и осенних листьев S. aucuparia были незначимы.
Рис. 1.
Средние для всех видов тест-растений относительная выживаемость (а) и относительная длина главного корня (б) при проращивании на вытяжках из зеленых летних (◇) и расцвеченных осенних (◆) листьев. Здесь и на рис. 2: water – вода; вытяжки: Sal. cap. – Salix caprea; Pru. pad. – Prunus padus; Sor. auc. – Sorbus aucuparia; Ace. neg. – Acer negundo; вертикальные линии – SE. Одинаковыми буквенными индексами показаны гомогенные по критерию Тьюки (при P < 0.05) значения.
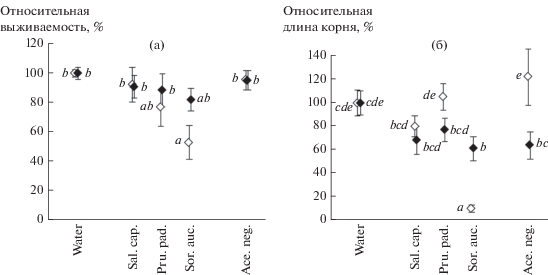
Сезонные особенности длины главного корня тест-растений проявлялись фактически исключительно в отношении вытяжек из S. aucuparia. Вытяжки как из зеленых летних, так и осенних расцвеченных листьев S. caprea, P. padus и A. negundo статистически значимо не угнетали рост корней по сравнению с водой (рис. 1б), но при этом вытяжки из расцвеченных осенних листьев в среднем обусловливали большее снижение относительной длины корня тест-растений, чем вытяжки из летних листьев. Противоположной была сезонная изменчивость влияния вытяжек из S. aucuparia: более сильное угнетающее воздействие на рост корней оказывали вытяжки из летних листьев (по критерию Тьюки парные различия между эффектами летних и осенних листьев были на уровне P = 0.0003).
Развитие тест-растений на вытяжках из листьев A. negundo ни летом, ни осенью не сопровождалось снижением роста корней в большей степени, чем на вытяжках из листьев местных деревьев.
Раннее развитие трав и A. negundo. Чтобы оценить конспецифичные и гетероспецифичные составляющие аллелопатической активности A. negundo, мы сравнили, как разные вытяжки влияли на раннее развитие травянистых тест-растений (Festuca rubra, Sinapis alba, Trifolium repens) и самого A. negundo. Для этого при проведении ANOVA фактор “вид тест-растений” заменили фактором “группа тест-растений” с двумя категориями: травы и A. negundo (рис. 2).
Рис. 2.
Средние для всех видов травянистых тест-растений (⚫) и для Acer negundo (⚪) относительная выживаемость (а) и относительная длина главного корня (б) при проращивании на разных вытяжках.
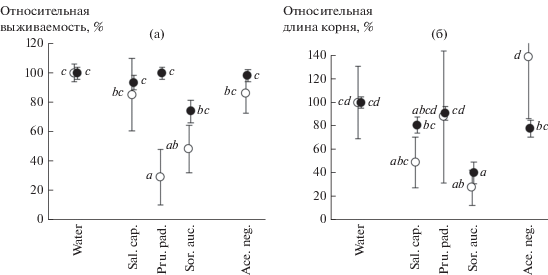
Средняя абсолютная (т.е. не относительная) в двух экспериментах выживаемость травянистых растений была в 3 раза выше (73 ± 2%), чем выживаемость A. negundo (24 ± 3%) (различия по парному критерию Тьюки P = 0.0001). Относительная выживаемость трав была самой низкой на вытяжках из листьев S. aucuparia, а A. negundo – на вытяжках из листьев S. aucuparia и P. padus. Вытяжки из листьев A. negundo не влияли каким-то особым образом ни на относительную выживаемость травянистых тест-растений, ни на относительную выживаемость A. negundo.
Средняя в двух экспериментах абсолютная (т.е. не относительная) длина главного корня у трав также была выше, чем у A. negundo, – 49 ± 2 и 29 ± 5 мм соответственно. Минимальная относительная длина корня зарегистрирована на вытяжках из листьев S. aucuparia и у трав (40 ± 9%), и у A. negundo (28 ± 15%). По парному критерию Тьюки различия относительной длины корня на воде и вытяжках из S. aucuparia достигали у трав уровня P = 0.0145, у A. negundo – P = 0.0353. Интересно, что на вытяжках из A. negundo длина корня проростков этого вида была больше, чем на воде, хотя и незначимо (относительная длина 138 ± 52%). При этом длина корней трав на вытяжках из A. negundo была меньше, чем на воде, хотя также незначимо (относительная длина 78 ± 7%). Однако в результате различия относительной длины корня трав и A. negundo при проращивании на вытяжках из A. negundo были значимы (по парному критерию Тьюки P = 0.0076). Следовательно, вытяжки из листьев A. negundo преимущественно отрицательно влияли на рост корней травянистых тест-растений и преимущественно положительно – на рост корней самого A. negundo. На примере реакций на вытяжки A. negundo можно продемонстрировать видовые особенности травянистых растений: относительная длина корня меньше всего была у F. rubra (54 ± 9%) и больше всего – у S. alba (109 ± 9%); промежуточное положение, ближе к F. rubra, занимал T. repens (71 ± 6%).
ОБСУЖДЕНИЕ
От многих экспериментов по оценке аллелопатической активности листовых вытяжек, в том числе проведенных на Acer negundo [14–18], наша работа отличается тем, что, наряду с дистиллированной водой, в качестве контролей мы использовали вытяжки из листьев местных растений. Этот подход дал неожиданные результаты. Использование местных видов деревьев для приготовления листовых вытяжек показало, что наша методика достаточно чувствительна и позволяет регистрировать отклики тест-растений, которые можно интерпретировать как указание на аллелопатическую активность. Мы наблюдали статистически надежные снижение выживаемости и изменение морфологических особенностей проростков при развитии тест-растений на одних вытяжках по сравнению с другими.
Результаты экспериментов с летними (зелеными) и осенними (расцвеченными) листьями в общем были близкими: и летом, и осенью на листовых вытяжках по сравнению с водой в среднем, хотя и не сильно, снижались выживаемость и длина корней тест-растений. При этом летом наблюдали большую дисперсию эффектов, связанных с видом растений-доноров, – от явного ингибирования до некоторого стимулирования. Осенью изменчивость эффектов вытяжек из разных видов древесных была меньше. Следует отметить, что при анализе сезонной изменчивости невозможно строго разделить эффекты, связанные, во-первых, с качеством листьев и, во-вторых, с условиями роста тест-растений. Два эксперимента – с летними и осенними листьями – провели последовательно. Хотя условия были стандартизированы, нельзя полностью исключить вероятность эффектов, связанных, например, с длиной светового дня или изменением качества семян. Однако можно отметить, что в контрольном варианте – на воде – средние значения всех признаков тест-растений летом и осенью статистически не различались. Это позволяет интерпретировать наблюдавшиеся сезонные особенности реакций тест-растений как преимущественно связанные с качеством листьев растений-доноров.
Заметный ингибирующий эффект на раннее развитие тест-растений оказал не инвазивный A. negundo, чего мы ожидали, а местный фоновый вид Sorbus aucuparia. При проращивании на вытяжках из ее листьев минимальными или близкими к минимальным были выживаемость и развитие корней. Ингибирующее воздействие S. aucuparia качественно не менялось в зависимости от того, проводился эксперимент летом или осенью, хотя ингибирующий эффект летних листьев был сильнее. Также ингибирующее воздействие S. aucuparia однозначно видно по реакции тест-растений, относящихся к жизненной форме и трав, и деревьев. Вытяжки из S. aucuparia, вероятно, действительно обладают аллелопатической активностью, конкретные механизмы которой было бы целесообразно исследовать. Согласно литературным данным [33–35], в листьях S. aucuparia подтвержден больший спектр вторичных метаболитов с потенциальной аллелопатической активностью: разнообразные органические кислоты, проантоцианиды, фенолы, флаваноиды, кверцетин. Интересны данные о высокой внутрисезонной изменчивости содержания фенольных соединений в листьях S. aucuparia с максимумом летом и снижением осенью [36]. Этим можно объяснить, почему вытяжки из летних листьев S. aucuparia имели более высокую ингибирующую активность по сравнению с вытяжками из осенних листьев. Заметим, что для двух растений-доноров из сем. Rosaceae, использованных в настоящей работе, по сравнению с другими местными видами древесных растений показано [37] наименьшее видовое богатство трав под их кронами (число видов трав/1 м2): P. padus – 5.9 ± 0.2; S. aucuparia – 6.2 ± 0.2; Pinus sylvestris – 7.0 ± 0.2; Salix spp. – 7.1 ± 0.4; Betula spp. – 7.1 ± 0.8; Populus tremula – 9.4 ± 0.5.
В среднем для всех тест-растений вытяжки из листьев A. negundo не показали ингибирующей активности как по сравнению с водой, так и по сравнению с вытяжками из листьев аборигенных деревьев. Влияние A. negundo на тест-растения было примерно таким же, как влияние S. caprea и P. padus, и слабее, чем ингибирующее влияние S. aucuparia. Таким образом, наша первая гипотеза не подтвердилась, и предположение об аллелопатическом воздействии A. negundo не является убедительным объяснением его способности влиять на местные сообщества.
Однако такая интерпретация справедлива, только если не учитывать особенности конспецифичных и гетероспецифичных эффектов. Такую специфику предполагала наша вторая рабочая гипотеза. Согласно нашим данным, есть вероятность, что гетероспецифичные эффекты вытяжек из листьев A. negundo могут быть более отрицательными, чем конспецифичные. Хотя это положение и не получило абсолютной поддержки, но и не отвергнуто. Установленные различия в реакции сеянцев A. negundo и сеянцев трав на вытяжки A. negundo небольшие, проявились они только в отношении длины корней и только с определенной долей условности их можно расценить как свидетельство того, что гетероспецифичные эффекты вытяжек из листьев A. negundo более негативны по сравнению с конспецифичными. Тем не менее стимулирование роста корней собственных сеянцев и угнетение роста корней трав вытяжками A. negundo свидетельствуют о том, что для понимания значения аллелопатических взаимодействий учет специфики тест-растений не менее важен, чем учет специфики растений-доноров. Предположение о разных направленности и силе гетероспецифичных и конспецифичных эффектов A. negundo нуждается в дальнейшей более строгой проверке в экспериментах, которые позволили бы одновременно оценить вероятность кон- и гетероспецифичных эффектов у нескольких видов растений.
Поскольку мы не можем полностью отвергнуть вторую рабочую гипотезу, неправильно утверждать, что несправедлива первая гипотеза. По-видимому, при исследовании аллелопатии A. negundo необходимо ставить вопросы с обязательным разграничением гетеро- и конспецифичных эффектов и в целом с учетом видовой специфики растений-реципиентов. Другими словами, вместо вопроса “Проявляет ли A. negundo аллелопатические способности?” правильнее ставить вопрос “В отношении каких видов-реципиентов A. negundo проявляет аллелопатические способности?”.
Вывод о слабой или неоднозначной способности к аллелопатическим воздействиям расходится с более ранними заключениями о том, что A. negundo обладает аллелопатической активностью [14–18]. Эти работы основаны на сравнениях эффектов от вытяжек из листьев A. negundo с водой, а не с другими растениями. Мы наблюдали отсутствие выраженных аллелопатических эффектов вытяжек из A. negundo как по сравнению с водой, так и по сравнению с вытяжками из листьев аборигенных деревьев.
Возможно, что механизмы успеха инвазивных растений, рассматриваемые в рамках гипотез нового оружия (NWH) и отсутствия естественных врагов (ERH), могут проявляться согласованно, поскольку обе гипотезы основаны на предположении синтеза специфически действующих метаболитов [38]. Если рассматривать эффекты, связанные с химическим составом листьев, такая согласованность не исключена. Возможно, что вещества, участвующие в защите от фитофагов, находясь в листьях, могут проявлять аллелопатическую активность после высвобождения из них. Ранее в отношении A. negundo были получены подтверждения как гипотезы ERH [39, 40], так и гипотезы NWH. Подтверждения гипотезы NWH распадаются на две группы: это или свидетельства возможности прямых аллелопатических эффектов [14–18], или свидетельства изменения под влиянием A. negundo почвенной биоты [19, 24, 41]. В пользу обоснованности гипотез, базирующихся на учете специфики химических веществ, свидетельствует широкий спектр у A. negundo вторичных метаболитов [42–45], включая алкалоиды, альдегиды, ароматические и гетероциклические соединения, карбоновые и жирные кислоты, кетоны, сложные эфиры, спирты, танины, терпеноиды, фитостероиды, флаваноиды и др. Для более надежного суждения о степени приложимости гипотезы NWH для объяснения инвазивного статуса A. negundo необходимо проведение не только лабораторных, но и вегетационных и полевых экспериментов разного дизайна.
ЗАКЛЮЧЕНИЕ
Эксперимент по оценке влияния водных вытяжек из листьев Acer negundo на раннее развитие нескольких видов травянистых растений и самого A. negundo дал неоднозначные результаты. В нашем эксперименте не удалось уверенно доказать, что водные вытяжки из листьев A. negundo влияют на раннее развитие тест-растений. Вместе с тем наши данные не позволяют исключить вероятность, что A. negundo может демонстрировать отрицательные эффекты при анализе гетероспецифичных взаимодействий, т.е. при влиянии на другие виды растений. В любом случае эффекты вытяжек из листьев A. negundo не отличались по силе от эффектов вытяжек из листьев местных растений Prunus padus и Salix caprea. Местный кустарник Sorbus aucuparia выраженно ингибировал раннее развитие тест-растений.
Таким образом, не получено свидетельств, что воздействия аллелопатического характера путем вымывания веществ из листьев могут быть реальным механизмом, обеспечивающим экологический успех A. negundo во вторичном ареале. Это заключение относится только к вытяжкам из листьев, хотя другие органы растений, например семена или корни, также могут обладать аллелопатической активностью. Нельзя также исключить, что аллелопатическая активность связана с климатическими условиями и может различаться в разных частях ареала. Поэтому для более определенных заключений об аллелопатии как механизме экологического успеха A. negundo необходимы дальнейшие исследования в широком географическом охвате и с использованием разных схем экспериментов.
Исследование выполнено при поддержке Российского фонда фундаментальных исследований (проект № 20-34-90084; О.С. Рафикова), а также в рамках темы государственного задания Института экологии растений и животных УрО РАН № AAAA-A19-119031890084-6 (Д.В. Веселкин).
Авторы декларируют отсутствие конфликта интересов.
Список литературы
Klironomos J.N. Feedback with soil biota contributes to plant rarity and invasiveness in communities // Nature. 2002. V. 417. № 6884. P. 67–70.
Callaway R.M., Ridenour W.M. Novel weapons: invasive success and the evolution of increased competitive ability // Front. Ecol. Environ. 2004. V. 2. № 8. P. 436–443.
Weidenhamer J.D., Callaway R.M. Direct and indirect effects of invasive plants on soil chemistry and ecosystem function // J. Chem. Ecol. 2010. V. 36. № 1. P. 59–69.
Anacker B.L., Klironomos J.N., Maherali H. et al. Phylogenetic conservatism in plant-soil feedback and its implications for plant abundance // Ecol. Lett. 2014. V. 17. № 12. P. 1613–1621.
Brouwer N.L., Hale A.N., Kalisz S. Mutualism-disrupting allelopathic invader drives carbon stress and vital rate decline in a forest perennial herb // AoB Plants. 2015. V. 7. plv014.
Kumar A.S., Bais H.P. Allelopathy and exotic plant invasion // Plant Communication from an Ecological Perspective. Springer, Berlin, Heidelberg, 2010. P. 61–74.
Zhang Z., Liu Y., Yuan L. et al. Effect of allelopathy on plant performance: a meta-analysis // Ecol. Lett. 2021. V. 24. № 2. P. 348–362. https://doi.org/10.1111/ele.13627
Olson B.E., Wallander R.T. Effects of invasive forb litter on seed germination, seedling growth and survival // Basic Appl. Ecol. 2002. V. 3. № 4. P. 309–317.
Blair A.C., Nissen S.J., Brunk G.R., Hufbauer R.A. A lack of evidence for an ecological role of the putative allelochemical (±)-catechin in spotted knapweed invasion success // J. Chem. Ecol. 2006. V. 32. № 10. P. 2327–2331.
Del Fabbro C., Güsewell S., Prati D. Allelopathic effects of three plant invaders on germination of native species: a field study // Biol. Invasions. 2014. V. 16. № 5. P. 1035–1042.
Gruntman M., Segev U., Glauser G., Tielbörger K. Evolution of plant defences along an invasion chronosequence: defence is lost due to enemy release–but not forever // J. Ecol. 2017. V. 105. № 1. P. 255–264.
Nielsen J.A., Frew R.D., Whigam P.A. et al. Germination and growth responses of co-occurring grass species to soil from under invasive Thymus vulgaris // Allelopathy J. 2015. V. 35. № 1. P. 139–152.
Виноградова Ю.К., Майоров С.Р., Хорун Л.В. Черная книга флоры Средней России: чужеродные виды растений в экосистемах Средней России. М.: ГЕОС, 2010. Т. 2. 512 с.
Csiszár Á. Allelopathic effects of invasive woody plant species in Hungary // Acta Silv. et Lignaria Hungarica. 2009. V. 5. P. 9–17.
Csiszár Á., Korda M., Schmidt D. et al. Allelopathic potential of some invasive plant species occurring in Hungary // Allelopathy J. 2013. V. 31. № 2. P. 309–318.
Еременко Ю.А. Аллелопатическая активность инвазионных древесных видов // Российский журн. биол. инвазий. 2014. Т. 7. № 2. С. 33–39.
Александров Д.С., Калашников Д.В. Влияние экстрактов листового опада клена остролистного и клена ясенелистного на прорастание семян и начальные этапы роста газонных трав и декоративных однолетников // Вестник ландшафтной архитектуры. 2019. № 19. С. 3–6.
Панасенко Н.Н., Володин В.В., Володченко Ю.С., Холенко М.С. Аллелопатические свойства Acer negundo // Ежегодник НИИ фундаментальных и прикладных исследований. 2018. № 1 (10). С. 34–36.
Веселкин Д.В., Рафикова О.С., Екшибаров Е.Д. Почва из зарослей инвазивного Acer negundo неблагоприятна для образования микоризы у аборигенных трав // Журн. общ. биол. 2019. Т. 80. № 3. С. 214–225. https://doi.org/10.1134/S0044459619030084
Цандекова О.Л., Уфимцев В.И. Формирование подстилки пойменных лесных биогеоценозов с участием Acer negundo L. // Изв. высших учебн. завед. Лесной журнал. 2019. № 3 (369). С. 73–81.
Putten W.H., Bardgett R.D., Bever J.D. et al. Plant-soil feedbacks: the past, the present and future challenges // J. Ecol. 2013. V. 101. № 2. P. 265–276.
Reinhart K.O., Callaway R.M. Soil biota and invasive plants // New Phytol. 2006. V. 170. P. 445–457. https://doi.org/10.1111/j.1469-8137.2006.01715.x
Dorning M., Cipollini D. Leaf and root extracts of the invasive shrub, Lonicera maackii, inhibit seed germination of three herbs with no autotoxic effects // Plant Ecol. 2006. V. 184. № 2. P. 287–296.
Saccone P., Pagès J.-P., Girel J. et al. Acer negundo invasion along a successional gradient: early direct facilitation by native pioneers and late indirect facilitation by conspecifics // New Phytol. 2010. V. 187. P. 831–842. https://doi.org/10.1111/j.1469-8137.2010.03289.x
Inderjit, Dakshini K.M.M. On laboratory bioassays in allelopathy // Bot. Rev. 1995. V. 61. № 1. P. 28–44.
Inderjit, Weston L.A. Are laboratory bioassays for allelopathy suitable for prediction of field responses? // J. Chem. Ecol. 2000. V. 26. № 9. P. 2111–2118.
John J., Patil R.H., Joy M., Nair A.M. Methodology of allelopathy research: 1. Agroforestry systems // Allelopathy J. 2006. V. 18. № 2. P. 173–214.
Adamowski W., Dvorak L., Ramanjuk I. Atlas of alien woody species of the Bialowieza Primaeval Forest // Phytocoenosis. 2002. V. 14. Suppl. Cartogr. Geobot. P. 1–303.
Drescher A., Magnes M. Anthropochoren im Nationalpark Donau-Auen-Ziel von Bekämpfungsmaßnahmen oder Bereicherung der Biodiversität? // 10 Österreichisches Botanikertreffen 30 Mai–1 Juni 2002. Bundesanstalt für alpenländliche Landwirtschaft Gumpenstein, 2002. S. 141–144.
Emelyanov A.V., Frolova S.V. Ash-leaf maple (Acer negundo L.) in coastal phytocenoses of the Vorona River // Rus. J. Biol. Invasions. 2011. V. 2. № 2–3. P. 161–163.
Kostina M.V., Yasinskaya O.I., Barabanshchikova N.S., Orlyuk F.A. Toward a issue of box elder invasion into the forests around Moscow // Russ. J. Biol. Invasions. 2016. V. 7. № 1. P. 47–51.
Veselkin D.V., Dubrovin D.I. Diversity of the grass layer of urbanized communities dominated by invasive Acer negundo // Rus. J. Ecol. 2019. V. 50. № 5. P. 413–421. https://doi.org/10.1134/S1067413619050114
Растительные ресурсы России: Дикорастущие цветковые растения, их компонентный состав и биологическая активность. Т. 2. Семейства Actinidiaceae – Malvaceae, Euphorbiaceae – Haloragaceae. / Отв. ред. А.Л. Буданцев. СПб; М.: Тов-во науч. изд. КМК, 2009. 513 с.
Olszewska M.A., Michel P. Antioxidant activity of inflorescences, leaves and fruits of three Sorbus species in relation to their polyphenolic composition // Nat. Prod. Res. 2009. V. 23. № 16. P. 1507–1521.
Gaivelyte K., Jakstas V., Razukas A., Janulis V. Variation of quantitative composition of phenolic compounds in rowan (Sorbus aucuparia L.) leaves during the growth season // Nat. Prod. Res. 2014. V. 28. № 13. P. 1018–1020. https://doi.org/10.1080/14786419.2014.902819
Olszewska M.A. Variation in the phenolic content and in vitro antioxidant activity of Sorbus aucuparia leaf extracts during vegetation // Acta Pol. Pharm. 2011. V. 68. № 6. P. 937–944.
Веселкин Д.В., Дубровин Д.И., Пустовалова Л.А., Коржиневская А.А. Влияют ли чужеродные деревья и кустарники на богатство напочвенного покрова: анализ в масштабе внутриценотической мозаики // Информационные технологии в исследовании биоразнообразия: Мат-лы III национ. науч. конф. с междунар. участием, посвященной 100-летию со дня рожд. акад. РАН П.Л. Горчаковского. Екатеринбург, 2020. С. 123–126.
Lau J.A., Schultheis E.H. When two invasion hypotheses are better than one // New Phytol. 2015. V. 205. P. 958–960. https://doi.org/10.1111/nph.13260
Пономарёва Е.А., Бессонова В.П. Структура повреждений листьев вредителями древесных растений в примагистральных лесополосах // Вестник Алтайского гос. аграрн. ун-та. 2016. № 1 (135). С. 77–82.
Veselkin D.V., Kuyantseva N.B., Chashchina O.E. et al. Levels of leaf damage by phyllophages in invasive Acer negundo and native Betula pendula and Salix caprea // Russ. J. Ecol. 2019. V. 50. № 6. P. 511–516. https://doi.org/10.1134/S1067413619060134
Reinhart K.O., Callaway R.M. Soil biota facilitate exotic Acer invasions in Europe and North America // Ecol. Appl. 2004. V. 14. № 6. P. 1737–1745.
Ping L., Shen Y., Jin Y. Volatiles released in succession from artificially damaged ash leaf maple leaves // Funct. Plant Biol. 2001. V. 28. № 6. P. 513–517.
Li J.G., Jin Y.J., Luo Y.Q. et al. Leaf volatiles from host tree Acer negundo: diurnal rhythm and behavior responses of Anoplophora glabripennis to volatiles in field // J. Integr. Plant Biol. 2003. V. 45. № 2. P. 177–182.
Bi W., Gao Y., Shen J. et al. Traditional uses, phytochemistry and pharmacology of the genus Acer (maple): a review // J. Ethnopharmacol. 2016. V. 189. P. 31–60.
Barrales-Cureño H.J., Salgado-Garciglia R., López-Valdez L.G. et al. Metabolomic data of phenolic compounds from Acer negundo extracts // Data in brief. 2020. V. 30. 105569. https://doi.org/10.1016/j.dib.2020.105569
Дополнительные материалы отсутствуют.