

Экология, 2022, № 4, стр. 312-318
Скорость деструкции и структура населения членистоногих опада инвазивного Solidago gigantea не подтверждают гипотезу “преимущества домашнего поля”
Е. Н. Устинова a, *, М. Н. Маслов a, С. Н. Лысенков a, А. В. Тиунов b
a Московский государственный университет им. М.В. Ломоносова
119991 Москва, Ленинские горы, 1, Россия
b Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
* E-mail: Ustinolena@ya.ru
Поступила в редакцию 28.12.2021
После доработки 03.02.2022
Принята к публикации 07.02.2022
- EDN: EGTXDL
- DOI: 10.31857/S0367059722040060
Аннотация
Скорость деструкции опада инвазивного растения в сообществах аборигенных видов может быть замедлена, поскольку организмы-деструкторы не адаптированы к опаду инвазивного вида. Мы сравнивали скорость деструкции растительных остатков и структуру сообщества членистоногих при инкубации опада инвазивного гигантского золотарника Solidago gigantea (Asteraceae) и трех видов аборигенных растений (Urtica dioica, Cirsium arvense и Chamaenerion angustifolium) при инкубации опада в биотопах с доминированием “своих” и “чужих” видов растений. Наши результаты позволяют предположить, что сообщество членистоногих, участвующих в разложении опада S. gigantea и других видов, не специализировано и не обеспечивает более высокую или более низкую скорость деструкции инвазивного вида. Ни скорость деструкции опада, ни структура и разнообразие сообществ членистоногих не подтверждают гипотезу “преимущества домашнего поля”.
Согласно гипотезе “преимущества домашнего поля” (home field advantage), деструкция растительного опада происходит быстрее под “материнским” растением, чем при перемещении его под растения других видов, так как организмы-деструкторы могут быть адаптированы к определенным видам растений [1, 2]. По данным обширного метаанализа (125 экспериментов из 35 исследований), в среднем разложение опада в “своем” сообществе происходит быстрее, но эффект проявляется преимущественно при сильных различиях в качестве “своего” и “чужого” опада [3]. Вопрос о преимуществе домашнего поля приобретает особый интерес в связи с влиянием на аборигенные сообщества инвазивных растений. Опад чужеродных растений может значительно отличаться от опада аборигенных [4–6], и организмы-деструкторы могут быть не адаптированы к нему.
Целью работы была проверка гипотезы домашнего поля на примере североамериканского гигантского золотарника Solidago gigantea Aiton (Asteraceae), одного из агрессивных инвазионных растений Палеарктики [7]. Мы сравнивали скорость деструкции опада гигантского золотарника и структуру сообщества населяющих опад членистоногих при инкубации опада в биотопах с разными доминирующими видами растений. В качестве контроля были использованы три вида аборигенных растений.
МАТЕРИАЛ И МЕТОДЫ
Сбор растительного материала и последующую инкубацию опада проводили на залежи в с. Луцино Одинцовского района Московской области (55°42.092′ с.ш., 36°45.653′ в.д.). Выбранная площадка представляет собой постагрогенную залежь возрастом 20 лет с дерново-подзолистой типичной почвой (Albic Retisols). Травяной фитоценоз представлен разнотравно-злаковым лугом. В сентябре 2018 г. были собраны увядшие побеги гигантского золотарника (S. gigantea) и аборигенных видов растений. Скорость деструкции разных видов опада во многом определяется соотношением C/N [8], поэтому для сравнения с золотарником (C/N в свежем опаде золотарника составило 33.3 ± 4.3 [здесь и далее среднее ± SD, n = 5]) были выбраны растения, имеющие более низкое (крапива двудомная Urtica dioica L. (Urticaceae) – 18.3 ± 1.9) и сходное (бодяк полевой Cirsium arvense (L.) Scop. (Asteraceae) – 34.6 ± 2.2 и иван-чай узколистный Chamaenerion angustifolium (L.) Scop. (Onagraceae) – 37.4 ± 2.5) соотношение C/N. Данные растения произрастают на выбранной залежи на расстоянии не более 150 м друг от друга. Каждое из четырех растений формирует в районе исследования плотные монодоминантные заросли площадью 2–6 м2, которые далее названы биотопами бодяка, золотарника, иван-чая и крапивы по соответствующему доминантному виду.
По 7 г (воздушно-сухой вес) листьев и стеблей поместили в нейлоновые мешки (размер 15 × 15 см, диаметр ячеи 1.5 мм). По 5 мешков с опадом каждого вида поместили на поверхность почвы в исследуемые биотопы 9 октября 2018 г. Для предотвращения схлопывания в мешок клали “каркас” (отрезанные от пластиковых бутылок кольца шириной 1–2 см). В пределах каждого биотопа расстояние между мешками было 15–20 см. Мешки закрепляли на поверхности почвы с помощью металлических колышков. Мешки с частично разложившимся опадом собрали через 8 мес., 5 июня 2019 г., определили его влажность, потерю сухой массы за время инкубации, общее содержание С и N (элементный анализатор Elementar Vario EL III).
Экстракцию членистоногих производили на воронках Тулльгрена перед высушиванием опада. Животных определяли по морфологическим признакам по возможности до вида (Oribatida, Mesostigmata, Collembola) или семейства (все остальные членистоногие, а также ювенильные стадии клещей). Всего обнаружено 2870 особей из 53 таксонов различного ранга, которые были включены в дальнейший анализ, несмотря на разный таксономический уровень идентификации.
Статистическую обработку данных проводили в программной среде R [9]. Для сравнения молярного соотношения С/N, скорости деструкции (потеря массы за 8 мес.) и разнообразия (индекс Шеннона, H) обитающих в опаде членистоногих использовали двухфакторный дисперсионный анализ с последующим применением критерия Тьюки. Для проверки гипотезы преимущества домашнего поля определяли наличие взаимодействия факторов “вид опада” и “биотоп”.
Кроме того, для оценки гипотезы преимущества домашнего поля для каждого вида опада рассчитывали индексы HFAi (Home Field Advantage index) [10, 11], основанные на сравнении относительной потери массы определенного вида опада, наблюдаемой в том биотопе, из которого он был получен (“своем”), с потерей массы, наблюдаемой в других биотопах (“чужих”). Индекс отражает величину “дополнительной” деструкции в своем биотопе: положительное значение индекса свидетельствует о наличии эффекта преимущества домашнего поля, нулевое – об отсутствии эффекта домашнего поля, а отрицательное – об отрицательном влиянии своего биотопа на скорость деструкции [10, 11]. Значимость отличия от нуля рассчитанных индексов HFAi оценивали по включению нуля в доверительный интервал (для нормального распределения), полученный путем процедуры бутстреппинга (999 реплик) в пакете “boot” [12] в R.
Для выявления связи между количественными переменными использовали ранговую корреляцию Спирмена (RS). Для оценки связи структуры сообщества членистоногих в опаде с параметрами среды (влажность, отношение С/N и скорость разложения опада) и влияния на сообщество членистоногих вида опада и биотопа применяли анализ избыточности (RDA) с фракционированием вариации в пакете vegan [13]. Значимость полученных моделей проверяли с помощью дисперсионного анализа, сравнивая различия в отклонении остатков вложенных моделей в перестановочном тесте (999 перестановок).
РЕЗУЛЬТАТЫ
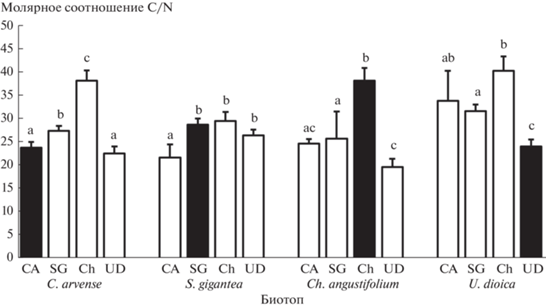
Соотношение C/N в опаде (рис. 1) значимо различалось в конце инкубации в зависимости от вида опада (p < 0.001; все четыре вида значимо различались, минимальное значение было у крапивы, максимальное – у иван-чая), биотопа (p < 0.001; соотношение C/N было в среднем выше в биотопе крапивы, чем в трех других) и взаимодействия этих факторов (p < 0.001; только у золотарника значение C/N не различалось значимо между биотопами).
Рис. 1.
Соотношение C/N в опаде 4 видов растений (CA – C. arvense, SG – S. gigantea, Ch – Ch. angustifolium, UD – U. dioica) после инкубации в разных биотопах (черные столбики обозначают вид опада, который инкубировался в “домашнем” биотопе). На рис. 1, 2 и 4 показаны среднее значение и стандартное отклонение (n = 5); разными буквами обозначены значимые различия в пределах одного биотопа (критерий Тьюки, p < 0.05).
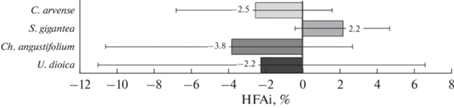
Скорость деструкции опада зависела от его вида (p < 0.001) и биотопа, в котором инкубировался образец (p < 0.001): опад крапивы разлагался быстрее по сравнению с другими видами, и разложение всех видов опада шло быстрее в биотопе крапивы (рис. 2). Однако взаимодействие факторов “вид опада” и “биотоп” было статистически незначимо (p = 0.707), что свидетельствует об отсутствии эффекта домашнего поля. Скорость деструкции коррелировала с разнообразием организмов, измеренным с помощью индекса Шеннона (RS = 0.240; p = 0.032), и влажностью опада (RS = 0.442; p < 0.001), но не с соотношением С/N (RS = –0.207; p = 0.066). Об отсутствии эффекта домашнего поля свидетельствует также невысокая величина индексов HFAi, которые ни для одного из видов опада значимо не отличались от нуля (рис. 3).
Рис. 2.
Потеря массы 4 видов растений (CA – C. arvense, SG – S. gigantea, Ch – Ch. angustifolium, UD – U. dioica) за 8 мес. инкубации в разных биотопах (черные столбики обозначают вид опада, который инкубировался в “домашнем” биотопе).

Рис. 3.
Индексы “преимущества домашнего поля” (HFAi), отражающие увеличение скорости деструкции опада 4 видов растений в “своем” биотопе. Приведены средние значения и 95%-ные доверительные интервалы (для нормального распределения), полученные методом бутстреппинга. Включение нуля в доверительный интервал свидетельствует об отсутствии значимых отличий от нуля.
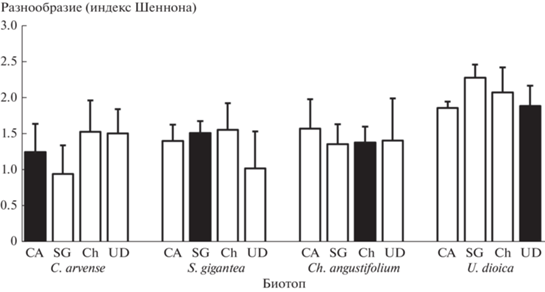
На разнообразие организмов, измеренное при помощи индекса Шеннона (рис. 4), значимо влиял биотоп (p < 0.001). В образцах опада в биотопе крапивы разнообразие было максимальным (H = = 2.0 ± 0.3 по сравнению с 1.4 ± 0.4 для остальных биотопов). Вид опада не влиял на разнообразие членистоногих (р = 0.446).
Рис. 4.
Разнообразие организмов, измеренное с помощью индекса Шеннона, в опаде 4 видов растений (CA – C. arvense, SG – S. gigantea, Ch – Ch. angustifolium, UD – U. dioica) при инкубации в разных биотопах (черные столбики обозначают вид опада, который инкубировался в “домашнем” биотопе). Критерий Тьюки не выявил значимых различий между разными видами опада.
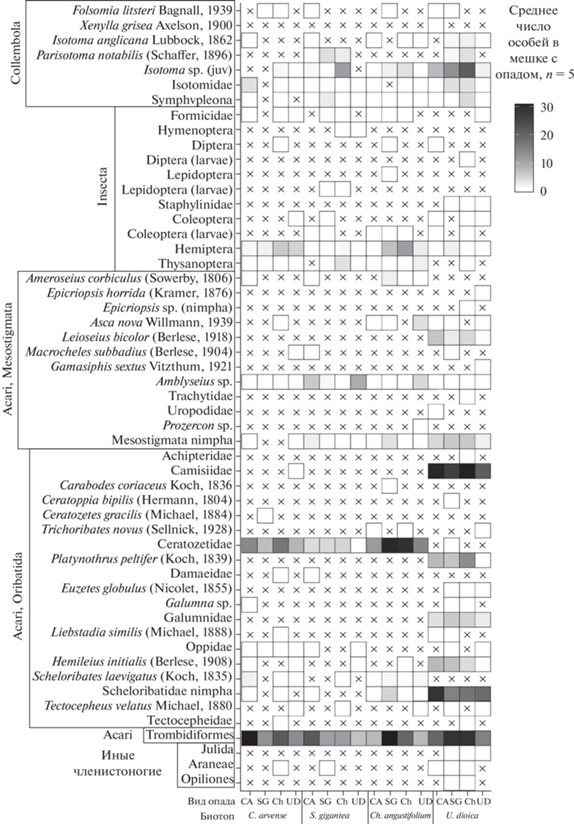
Общая численность животных (рис. 5) в биотопе крапивы также была выше (p < 0.001 при сравнении биотопа крапивы с каждым из трех остальных критерием Тьюки) в основном за счет более высокой численности панцирных клещей из семейств Scheloribatidae, Galumnidae, Camisiidae и ювенильных особей Isotoma sp. (Collembola).
Рис. 5.
Численность организмов в разных видах опада (CA – C. arvense, SG – S. gigantea, Ch – Ch. angustifolium, UD – U. dioica) и разных биотопах. Крестиком отмечено отсутствие данного таксона в соответствующих пробах.
Структура обитающих в опаде членистоногих была связана с соотношением С/N (4.8% объясненной дисперсии, p = 0.005), скоростью разложения опада (9.9%, p = 0.001) и влажностью (8.1%, p = 0.003). На структуру сообщества обитающих в опаде членистоногих оказывал значимое влияние биотоп (22.8%, p = 0.001), но не вид опада (2.1%, p = 0.099). Взаимодействия данных факторов не обнаружено. Это предполагает, что структура сообщества почвенных членистоногих не зависела от инкубации опада в “своем” или “чужом” биотопе (рис. 6).

ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Полученные результаты не подтвердили гипотезу “преимущества домашнего поля”: эффект более быстрого разложения опада в “своем” биотопе не проявился ни при сравнении аборигенных и инвазивного видов, ни при сравнении видов из разных семейств. Это согласуется с результатами исследования скорости разложения опада инвазивных растений (Bromus tectorum, Centaurea stoebe, Euphorbia esula) в сообществах инвазивных и аборигенных растений на горных пастбищах, где также не обнаружено повышенной скорости деструкции при инкубации опада в “своем” сообществе [14]. Однако эффект “домашнего поля”, как правило, влияет на скорость разложения менее чем на 10% [3]. Размер нашей выборки и в работе [14] мог оказаться недостаточным для выявления такого незначительного влияния. В пользу этого предположения свидетельствует и высокая вариация индексов HFAi, которые ни для одного из четырех видов опада не отличались значимо от нуля (см. рис. 3).
Интересно отметить, что для всех видов опада, кроме золотарника, индексы HFAi были меньше нуля, т.е. наблюдается тенденция к отрицательному влиянию своего биотопа на скорость деструкции опада. Более быстрое разложение опада в чужом биотопе было отмечено и в других работах [10, 15] и объясняется, как правило, тем, что опад низкого качества может разлагаться быстрее в чужом сообществе с опадом более высокого качества [10, 16]. C другой стороны, в биотопах с низким качеством опада ограниченные в питательных веществах редуценты могут привлекаться внесенным опадом более высокого качества, в результате чего он будет разлагаться быстрее, чем ожидалось [3, 17, 18]. В целом метаанализ 35 работ по изучению скорости деструкции опада разных видов деревьев в своих и чужих биотопах выявил большой разброс в эффекте домашнего поля: от уменьшения скорости разложения в своем биотопе на 9% до увеличения на 29% [2]. В экспериментах с опадом травянистых растений величина эффекта еще меньше [3], что согласуется с невысокими значениями индексов HFAi в нашей работе.
Сильно отличающийся по содержанию азота опад крапивы быстрее разлагался в биотопе крапивы, но это не является подтверждением гипотезы “преимущества домашнего поля”, так как опад крапивы имел высокую скорость деструкции во всех биотопах, и скорость деструкции любого вида опада повышалась в биотопе крапивы. Повышение скорости деструкции могло быть связано с переносом азота из богатой азотом почвы биотопа в опад растений [19], однако мы не зафиксировали снижения соотношения С/N опада при инкубации в биотопе крапивы. Не обнаружено и статистически значимой корреляции между скоростью деструкции и соотношением С/N.
Разнообразие и обилие членистоногих было выше в биотопе крапивы вне зависимости от вида опада, что могло способствовать более высоким темпам деструкции [18]. Однако структура сообщества членистоногих также не испытывала влияние эффекта “преимущества домашнего поля”. При исследовании сообществ деструкторов опада пастбищ и раннесукцессионных лесов в “своем” и “чужом” местообитаниях [16] было обнаружено, что вторичные деструкторы (клещи) более специализированы к местообитаниям в отличие от первичных деструкторов (бактерий и грибов). Наша работа также демонстрирует, что и разнообразие, и структура сообщества почвенных животных зависят от биотопа, но не от вида опада. Эксперименты по перемещению сообщества панцирных клещей из подстилки дуба и сосны в подстилку инвазивной робинии (Robinia pseudoacacia) в Венгрии [20] показали, что подстилка робинии благоприятна только для адаптированного к ее опаду сообщества клещей, но не для сообществ клещей из подстилки дуба и сосны. Авторы предположили, что это может быть связано с низким разнообразием сапротрофных грибов, ассоциированных с подстилкой робинии. В нашем случае инвазионный статус растения не повлиял на состав сообщества членистоногих. Несмотря на то, что инвазия золотарника (S. gigantea или близкого вида S. canadensis) оказывает влияние на почвенные грибы [21], азотофиксирующие бактерии [22], почвенную микробную биомассу и ее активность [23, 24], сообщество почвенных членистоногих, видимо, более устойчиво к инвазии этого вида и не претерпевает значительных изменений при его внедрении [25, 26].
Таким образом, наши результаты предполагают, что сообщество членистоногих, участвующих в разложении опада инвазивного вида S. gigantea, не специализировано к нему и не обеспечивает более высокую скорость деструкции. Биотоп крапивы поддерживает более высокое разнообразие подстилочных членистоногих, но они также не демонстрируют специализации, поскольку эффективно перерабатывают не только опад крапивы, но и другие виды опада. Ни скорость деструкции опада, ни структура и разнообразие сообществ членистоногих не подтверждают гипотезу “преимущества домашнего поля”.
Мы благодарим специалистов, помогавшим в определении почвенных членистоногих: Collembola – А.Ю. Короткевич (каф. зоологии и экологии МПГУ), Oribatida – В.Д. Леонов (ИПЭЭ РАН), Mesostigmata – М.С. Бизин (ИПЭЭ РАН).
Список литературы
Gholz H.L., Wedin D.A., Smitherman S.M. et al. Long-term dynamics of pine and hardwood litter in contrasting environments: toward a global model of decomposition // Glob. Chang. Biol. 2000. V. 6. № 7. P. 751–765.
Ayres E., Steltzer H., Simmons B.L. et al. Home-field advantage accelerates leaf litter decomposition in forests // Soil Biol. Biochem. 2009. V. 41. № 3. P. 606–610.
Veen G.F., Freschet G.T., Ordonez A., Wardle D.A. Litter quality and environmental controls of home-field advantage effects on litter decomposition // Oikos. 2015. V. 124. № 2. P. 187–195.
Ehrenfeld J.G. Effects of exotic plant invasions on soil nutrient cycling processes // Ecosystems. 2003. V. 6. № 6. P. 503–523.
Allison S.D., Vitousek P.M. Rapid nutrient cycling in leaf litter from invasive plants in Hawai’i // Oecologia. 2004. V. 141. № 4. P. 612–619.
Ashton I.W., Hyatt L.A., Howe K.M. et al. Invasive species accelerate decomposition and litter nitrogen loss in a mixed deciduous forest // Ecol. Appl. 2005. V. 15. № 4. P. 1263–1272.
Самые опасные инвазионные виды России (ТОП-100) / Ред. Дгебуадзе Ю.Ю., Петросян В.Г., Хляп Л.А. М.: КМК, 2018. 688 с.
Berg B., McClaugherty C. Plant litter: decomposition, humus formation, carbon sequestration. Berlin: Springer, 2008. 338 p.
R Core Team. R: A language and environment for statistical computing. //R Foundation for Statistical Computing. Vienna, Austria, 2020.
Perez G., Aubert M., Decaëns T. et al. Home-field advantage: a matter of interaction between litter biochemistry and decomposer biota //Soil Biol. Biochem. 2013. V. 67. P. 245–254.
Ayres E., Steltzer H., Berg S., Wall D.H. Soil biota accelerate decomposition in high-elevation forests by specializing in the breakdown of litter produced by the plant species above them //Journal of Ecology. 2009. V. 97. № 5. P. 901–912.
Canty A., Ripley B. Boot: Bootstrap R (S-Plus) functions (R package version 1.3-9). Vienna, Austria: R Foundation for Statistical Computing, 2013.
Oksanen J., Kindt R., Legendre P. et al. The vegan package // Community ecology package. 2007. V. 10. P. 631–637.
McTee M.R., Lekberg Y., Mummey D. et al. Do invasive plants structure microbial communities to accelerate decomposition in intermountain grasslands? // Ecol. Evol. 2017. V. 7. № 24. P. 11227–11235.
Gießelmann U.C., Martins K.G., Brändle M. et al. Lack of home-field advantage in the decomposition of leaf litter in the Atlantic Rainforest of Brazil // Appl. Soil Ecol. 2011. V. 49. P. 5–10.
John M.G.S., Orwin K.H., Dickie I.A. No ‘home’ versus ‘away’ effects of decomposition found in a grassland–forest reciprocal litter transplant study //Soil Biol. Biochem. 2011. V. 43. № 7. P. 1482–1489.
Gartner T. B., Cardon Z. G. Decomposition dynamics in mixed-species leaf litter //Oikos. 2004. V. 104. № 2. P. 230–246.
Hättenschwiler S., Tiunov A.V., Scheu S. Biodiversity and litter decomposition in terrestrial ecosystems // Annu. Rev. Ecol. Evol. Syst. 2005. V. 36. P. 191–218.
Shilenkova O.L., Tiunov A.V. Soil-litter nitrogen transfer and changes in δ13C and δ15N values in decomposing leaf litter during laboratory incubation // Pedobiologia. 2013. V. 56. № 3. P. 147–152.
Gergócs V., Rétháti G., Hufnagel L. Litter quality indirectly influences community composition, reproductive mode and trophic structure of oribatid mite communities: a microcosm experiment // Exp. Appl. Acarol. 2015. V. 67. № 3. P. 335–356.
Stefanowicz A.M., Stanek M., Nobis M., Zubek S. Species-specific effects of plant invasions on activity, biomass, and composition of soil microbial communities // Biol. Fert. Soils. 2016. V. 52. № 6. P. 841–852.
Wang C., Jiang K., Zhou J., Wu B. Solidago canadensis invasion affects soil N-fixing bacterial communities in heterogeneous landscapes in urban ecosystems in East China // Sci. Total Environ. 2018. V. 631. P. 702–713.
Zhang C.B., Wang J., Qian B.Y., Li W.H. Effects of the invader Solidago canadensis on soil properties // Appl. Soil Ecol. 2009. V. 43. № 2–3. P. 163–169.
Scharfy D., Güsewell S., Gessner M.O., Venterink H.O. Invasion of Solidago gigantea in contrasting experimental plant communities: effects on soil microbes, nutrients and plant-soil feedbacks // J. Ecol. 2010. V. 98. № 6. P. 1379–1388.
Sterzyńska M., Shrubovych J., Nicia P. Impact of plant invasion (Solidago gigantea L.) on soil mesofauna in a riparian wet meadows // Pedobiologia. 2017. V. 64. P. 1–7.
Ustinova E.N., Schepetov D.M., Lysenkov S.N., Tiunov A.V. Soil arthropod communities are not affected by invasive Solidago gigantea Aiton (Asteraceae), based on morphology and metabarcoding analyses // Soil Biol. Biochem. 2021. V. 159. P. 108 288.
Дополнительные материалы отсутствуют.