

Экология, 2023, № 3, стр. 212-219
Новый метод определения календарного возраста растений Heracleum sosnowskyi и оценка на его основе возрастного состава в ценопопуляциях вида на Севере
И. В. Далькэ a, *, С. П. Маслова a, С. Н. Плюснина a, Е. С. Зрайченко b, Ю. А. Бобров b
a Институт биологии Коми научного центра УрО РАН
167982 Сыктывкар, ул. Коммунистическая, 28, Россия
b ФГБОУ ВО “СГУ им. Питирима Сорокина”
167001 Сыктывкар, Октябрьский просп., 55, Россия
* E-mail: dalke@ib.komisc.ru
Поступила в редакцию 03.11.2022
После доработки 09.12.2022
Принята к публикации 31.01.2023
- EDN: EARBLW
- DOI: 10.31857/S0367059723030022
Аннотация
Проведены исследования по определению календарного возраста растений инвазионного вида борщевика Сосновского (Heracleum sosnowskyi Manden.). Изучение структуры корня показало наличие годичных колец, образованных приростами вторичной ксилемы, что позволяет использовать ксилохронологический подход для оценки календарного возраста H. sosnowskyi. Количество ксилемных колец на поперечном срезе корня соответствует числу остатков годичных приростов на продольном срезе подземного каудекса. Метод учета годичных приростов на продольном срезе каудекса является простым и эффективным и позволяет определить возраст растений H. sosnowskyi в полевых условиях, не прибегая к контрастированию тканей и микроскопии. Растения H. sosnowskyi в природных ценопопуляциях среднетаежной зоны Республики Коми приступали к цветению на третий год жизни, а их календарный возраст достигал 7 полных лет.
Определение возраста многолетних травянистых растений является сложной задачей и слабо изученным вопросом биологии травянистых многолетников. Анализ показал [1], что крупнейшая база данных по учету признаков растений в мире TRY (более 2 млрд. записей) содержит крайне малое количество записей (не более 6 тыс.) о возрасте растений, приступающих к цветению, или о потенциальном максимальном возрасте видов. Сведения о возрасте многолетних трав могут быть получены на основе анализа морфологических признаков растений, учета ежегодно отмирающих особей, оценки онтогенетического состояния растений [2, 3]. В последние десятилетия для оценки календарного возраста многолетних трав используют метод ксилохронологии, основанный на изучении годичных колец ─ слоев ежегодного прироста вторичной ксилемы корневой системы. Выраженность колец зависит от таксономической, морфологической и экологической специфики исследуемых объектов [4–6].
Оценку календарного возраста травянистых многолетников используют для изучения онтогенеза, определения скорости возобновления, расселения и распространения растений и их экологических стратегий [5, 7, 8]. Такие характеристики являются ключевыми при изучении биологии инвазионных видов, которые негативно влияют на биоразнообразие природных экосистем и культурных фитоценозов [9].
Борщевик Сосновского (Heracleum sosnowskyi Manden.) – один из широко распространенных чужеродных видов. В условиях подзоны средней тайги Республики Коми его моновидовые заросли встречаются повсеместно. Растение входит в списки агрессивных инвазионных видов, существенно влияет на структуру и разнообразие растительных и почвенных сообществ [10]. Исследования по определению возраста растений H. sosnowskyi единичны. В условиях севера календарный возраст растений в коллекции достигал 14–16 лет [11, 12]. Изучение возрастного состава ценопопуляций близкородственного вида Heracleum mantegazzianum показало, что в природных условиях растения достигают возраста 12 лет [13].
Сведения о возрасте инвазионных борщевиков в ходе жизненного цикла тесно связаны с разработкой их индивидуальных демографических моделей [14, 15]. Оценка времени перехода особей H. sosnowskyi в репродуктивное состояние в совокупности с расчетом дальности распространения семян [16] позволяет предсказывать направление и скорость их биологического вторжения, эффективно выполнять мероприятия по ликвидации нежелательных растений.
Целью работы был поиск эффективного метода изучения календарного возраста растений H. sosnowskyi на основе известных и новых методов определения возраста многолетних трав. Задачей исследований был анализ возрастного состава ценопопуляций растений H. sosnowskyi в условиях севера на основе предложенного метода.
МАТЕРИАЛ И МЕТОДЫ
Объектом исследования был борщевик Сосновского (Heracleum sosnowskyi Manden.) – многолетний, летнезеленый, травянистый, стержнекорневой, моноподиально нарастающий монокарпик с полурозеточным прямостоячим побегом [17]. Изучали растения, произрастающие в подзоне средней тайги Республики Коми на залежных участках с автоморфными подзолистыми почвами в окрестностях г. Сыктывкара. Исследование выполняли на 4 участках с географическими координатами: участок 1 – 61.662834° с.ш., 50.822097° в.д.; участок 2 – 61.650820° с.ш., 50.841117° в.д.; участок 3 – 61.645695° с.ш., 50.731975° в.д.; участок 4 – 61.607331° с.ш., 50.735631° в.д. Растительный покров участков представлен сообществами с доминированием растений H. sosnowskyi, которые длительное время не подвергались скашиванию или химической обработке. В сообществах H. sosnowskyi встречалось до 28 видов других растений (Urtica dioica, Poa pratensis, Symphytum asperum, Chamaenerion angustifolium, Elytrigia repens, Calamagrostis epigejos, Artemisia vulgaris и др.) с обилием 5–10%.
Для ретроспективного анализа и подтверждения присутствия растений H. sosnowskyi на выбранных участках с помощью программного обеспечения Google Earth Pro 7.3.4.8642 (http://google.com/) были просмотрены архивные спутниковые изображения местности. Скопления растений H. sosnowskyi хорошо заметны на спутниковых снимках благодаря раннему отрастанию весной и соцветиям белого (в период цветения) или коричневого (в период плодоношения) оттенков [18, 19].
Исследования проводили в течение вегетации – с апреля по сентябрь 2022 г. Для определения плотности растений H. sosnowskyi в ценопопуляциях на выбранных участках заложили 25 площадок размером 1 × 1 м. На площадках выкапывали все особи H. sosnowskyi, за исключением небольших по габитусу молодых растений с площадью листьев до 10 см2. Для сбора молодых растений использовали почвенный пробоотборник с внутренним диаметром 10.5 см, высотой 3.5 см и площадью 86.5 см2. Почвенные пробы просеивали через сито и подсчитывали в них число проростков и ювенильных растений. Всего проанализировано 40 почвенных проб.
Подземную часть собранных растений переносили в лабораторию, отмывали от почвы и проводили морфоструктурный анализ: измеряли наибольшую ширину каудекса и длину корня, глубину залегания почки возобновления, взвешивали сырую массу. Выполняли продольный срез через середину каудекса и подсчитывали количество годичных приростов.
Календарный возраст растений H. sosnowskyi определяли с помощью методов, описанных в литературе, и предложенного нами нового подхода. После внешнего осмотра на поверхности каудекса подсчитывали число кольцеобразных годичных групп листовых рубцов, которые остаются после регулярного отмирания листьев [12, 20].
На поперечных срезах базальной части главного корня возраст растений определяли методом ксилохронологии, подсчитывая число колец ежегодного прироста ксилемы [4, 6]. Для этого у каждого растения делали 2–3 поперечных среза главного корня вблизи каудекса. Срезы обрабатывали 0.5%-ным раствором флороглюцина в смеси спирта и дистиллированной воды (1 : 1) в течение 1–2 мин, а затем воздействовали концентрированной соляной кислотой. Одревесневшие оболочки ксилемы окрашивались в отчетливый красно-малиновый цвет [21]. Для верификации метода были изучены зоны прироста ксилемы на поперечных срезах корней растений первого и второго года жизни. Временные препараты с окрашенными поперечными срезами просматривали под микроскопом Axiovert 200 M (Carl Zeiss, Германия) в режимах светлого поля и фазового контраста, неокрашенные образцы – в режиме флуоресценции. Фотосъемку выполняли цифровой камерой AxioCam ERc 5s (Carl Zeiss, Германия). Диаметр клеток ксилемы и толщину годичных колец (n > 50 для каждого параметра) измеряли при помощи программы ZEN 2011 (Carl Zeiss, Германия).
Предложенный нами метод определения календарного возраста H. sosnowskyi основан на подсчете количества остатков годичных приростов каудекса, для чего выполняли продольный разрез через его центр. Число годичных приростов на срезе соответствовало календарному возрасту образца. В тексте, таблице и на рисунках приведены средние значения и стандартные отклонения.
РЕЗУЛЬТАТЫ
Ретроспективный анализ спутниковых изображений показал, что растения H. sosnowskyi произрастали на модельных участках не менее 10 лет. В 2022 г. средняя плотность растений без учета проростков текущего года составляла 17 ± 7 шт./м2 участка.
Подземная часть растений H. sosnowskyi представлена погруженным каудексом – базальным многолетним участком монокарпического побега из резидов однолетних вегетативных побегов с верхней частью главного корня (рис. 1). На верхушке каудекса закладывается терминальная зимующая почка возобновления. Главный корень ветвится до 3–4-го порядков, при этом все корни имеют контрактильные свойства, способствующие заглублению верхушки вегетативного растения.
Рис. 1.
Подземная часть растений Heracleum sosnowskyi: 1 – подземный каудекс с группами листовых рубцов (ЛР); 2 – главный корень (ГК) и боковые корни (БК).
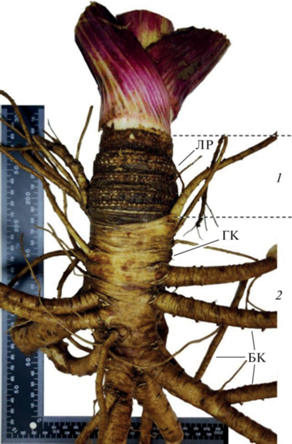
Изучение глубины залегания почек возобновления H. sosnowskyi показало, что она увеличивалась с возрастом и составляла в среднем 9 см у 3─ 6-летних растений (табл. 1). Ширина каудекса, а также длина и биомасса подземной части вместе с корнями достигали наибольших значений у растений в возрасте от 3 до 5 лет.
Таблица 1.
Морфоструктура подземных органов растений Heracleum sosnowskyi разного возраста
| Показатель | Возраст растений, полных лет жизни | |||||||
|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | |
| Объем выборки | 20 | 11 | 25 | 31 | 22 | 18 | 8 | 3 |
| Длина каудекса и корней, см | $\frac{{8 \pm 3}}{{4 - 13}}$ | $\frac{{17 \pm 5}}{{10 - 30}}$ | $\frac{{28 \pm 18}}{{12 - 90}}$ | $\frac{{38 \pm 18}}{{15 - 90}}$ | $\frac{{41 \pm 21}}{{10 - 90}}$ | $\frac{{38 \pm 12}}{{25 - 65}}$ | $\frac{{33 \pm 22}}{{17 - 80}}$ | $\frac{{23 \pm 2}}{{20 - 25}}$ |
| Ширина каудекса, см | $\frac{{0.4 \pm 0.1}}{{0.2 - 0.6}}$ | $\frac{{1 \pm 1}}{{1 - 2}}$ | $\frac{{2 \pm 2}}{{1 - 7}}$ | $\frac{{4 \pm 2}}{{1 - 10}}$ | $\frac{{4 \pm 3}}{{1 - 9}}$ | $\frac{{4 \pm 1}}{{2 - 6}}$ | $\frac{{4 \pm 3}}{{2 - 8}}$ | $\frac{{3 \pm 1}}{{2 - 4}}$ |
| Глубина залегания почки возобновления, см | $\frac{{4 \pm 1}}{{1 - 6}}$ | $\frac{{6 \pm 4}}{{2 - 16}}$ | $\frac{{7 \pm 4}}{{3 - 17}}$ | $\frac{{9 \pm 4}}{{4 - 20}}$ | $\frac{{9 \pm 6}}{{3 - 25}}$ | $\frac{{9 \pm 4}}{{5 - 18}}$ | $\frac{{9 \pm 5}}{{3 - 20}}$ | $\frac{{7 \pm 1}}{{6 - 8}}$ |
| Сырая масса каудекса и корней, г | $\frac{{0.5 \pm 0.3}}{{0.1 - 1.1}}$ | $\frac{{6 \pm 6}}{{2 - 20}}$ | $\frac{{146 \pm 232}}{{3 - 800}}$ | $\frac{{270 \pm 373}}{{3 - 1340}}$ | $\frac{{339 \pm 396}}{{7 - 1360}}$ | $\frac{{194 \pm 209}}{{7 - 600}}$ | $\frac{{330 \pm 513}}{{7 - 1400}}$ | $\frac{{31 \pm 1}}{{30 - 31}}$ |
| Доля генеративных особей, % | 0 | 0 | 12 | 23 | 18 | 11 | 25 | 0 |
Показатели морфоструктуры каудекса и корня существенно варьировали у особей одного возраста и между разновозрастными группами. Коэффициент вариации размеров подземных органов, глубины залегания почек возобновления был на уровне 50%, сырая масса характеризовалась изменчивостью 100–170%.
Растения H. sosnowskyi переходили к цветению в возрасте двух полных лет или позже, т.е. не ранее, чем на третий год от прорастания семени. Доля генеративных особей варьировала от 11 до 25% и была наибольшей у 3- и 6-летних растений (см. табл. 1).
Анализ структуры каудекса H. sosnowskyi показал, что на складчатой поверхности после ежегодного отмирания листьев остаются кольцеобразные группы листовых рубцов, которые располагаются в виде валиков (см. рис. 1). По числу групп листовых рубцов можно определить возраст видов рода Heracleum. Выявлено, что листовые рубцы лучше всего различимы у растений с шириной каудекса от 4 до 10 см в возрасте от 2 до 7 лет. Следует отметить, что доля таких растений не превышала 30% от общего количества изученных. Складчатая поверхность побеговой части каудекса растений H. sosnowskyi значительно затрудняла подсчет колец с листовыми рубцами.
Анализ окрашенных поперечных срезов корня H. sosnowskyi показал формирование хорошо выраженных зон ежегодного прироста ксилемы в центральном цилиндре – колец ксилемы (рис. 2). На поперечном срезе корня растений первого года жизни ряды водопроводящих элементов ксилемы расходятся радиально от центра к периферии стелы и разделены лучами паренхимы (см. рис. 2а). У изученных растений средний диаметр сосудов варьировал от 13 до 51 мкм. К концу периода вегетации по периферии центрального цилиндра в большей части лучей водопроводящие клетки располагаются группами, что создает выраженную визуальную границу прироста ксилемы в первый год жизни. Диаметр стелы на поперечном срезе корня первого года жизни составлял 1–2 мм. В последующие годы диаметр сосудов ксилемы варьировал от 28 до 109 мкм (см. рис. 2б). Лучи водопроводящих клеток ксилемы расширялись за счет формирования групп широкопросветных сосудов в конце периода вегетации. Толщина прироста ксилемы на второй и в последующие годы функционирования корня варьировала от 0.2 до 2.2 мм. Между зонами годичного прироста ксилемы формировалось в разной степени выраженное кольцо паренхимы.
Рис. 2.
Фрагмент поперечного среза главного корня растений Heracleum sosnowskyi первого (а) и шестого (б) года жизни: флороглюцин + HCl; фазовый контраст; КС – сосуды ксилемы, ПЦЦ – паренхима центрального цилиндра. Пунктирной линией обозначены границы годичных колец.
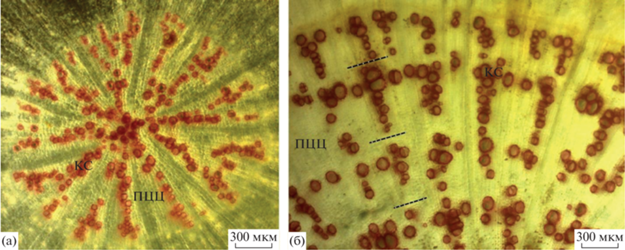
Описанные выше ежегодно повторяющиеся особенности формирования водопроводящей системы у H. sosnowskyi позволяют визуально разделять годичные слои прироста ксилемы – годичные кольца. Годичные кольца на поперечных срезах корней хорошо просматривались у 85% изученных особей (104 образца) в возрасте от 0 до 7 полных лет жизни. В остальных случаях четкие границы между годичными кольцами отсутствовали, и определить возраст растений методом ксилохронологии было невозможно.
Исследования показали, что на радиальных срезах каудекса H. sosnowskyi хорошо просматриваются границы годичных побегов, сформировавшихся в предыдущие годы жизни (годичные приросты). По нашим данным, в подзоне средней тайги в течение вегетационного периода у вегетативных растений образуются в среднем 2–3 листа. Поскольку несущие их метамеры укорочены, ежегодно остающиеся листовые рубцы, сформированные в течение одного вегетационного сезона, сильно сближены. При внешнем осмотре они расположены непосредственно друг под другом вокруг каудекса, а их группа – на едином валике в виде кольца, междоузлия не выражены. Однако междоузлия между группами узлов листьев разных лет жизни хорошо определяются при внешнем осмотре. Такое хорошо выраженное междоузлие и группа сближенных узлов составляют годичный прирост каудекса. На продольном разрезе в паренхимной сердцевине каудекса в конце каждого вегетационного сезона формируется разделительная зона, состоящая из сильно сжатых в горизонтальной плоскости тонкостенных клеток, на месте которых часто образуются разрывы. Участки паренхимной сердцевины, ограниченные этими разрывами, по сути являются визуальным отображением годичного прироста на продольном разрезе каудекса. Количество годичных приростов соответствовало числу ксилемных колец в основании главного корня (рис. 3), т.е. числу вегетационных периодов.
Рис. 3.
Продольный срез каудекса (а) и поперечный срез главного корня (б) генеративной особи Heracleum sosnowskyi на шестой год жизни (г.ж.): флороглюцин + HCl; 1 – группы листовых рубцов, 2 – зоны разделения годичных побегов, 3 – годичные кольца.
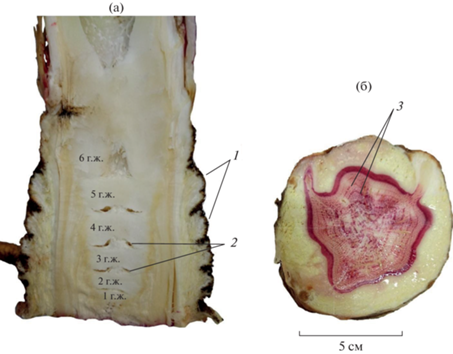
Таким образом, для определения возраста H. sosnowskyi на продольном (радиальном) срезе каудекса следует подсчитывать число остатков годичных приростов, отделенных друг от друга разделительной зоной, хорошо распознаваемой без окрашивания тканей.
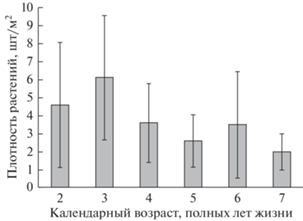
Исследование возрастной структуры ценопопуляций H. sosnowskyi показало, что количество молодых растений, выросших из семян в текущем году, и особей в возрасте одного полного года жизни составляло 370 ± 309 и 199 ± 288 шт/м2 соответственно. Средняя плотность остальных растений в возрасте двух полных лет жизни и старше была на порядок ниже – 17 ± 7 шт/м2. Наибольший календарный возраст растений составил 7 полных лет. С увеличением возраста количество особей в составе возрастных групп снижалось (рис. 4).
ОБСУЖДЕНИЕ
На основе полученных нами данных о расположении почек возобновления в почве растения H. sosnowskyi следует отнести к геофитам, а не к гемикриптофитам, как было указано ранее [12]. Погружение каудекса на глубину до 10 см способствует защите почек возобновления от вымерзания в условиях сезонного климата [22]. Заглубление каудекса осуществляется благодаря формированию контрактильной корневой системы, что также обеспечивает поддержание массивных черешков и листьев H. sosnowskyi, формирующих характерную для вида архитектонику ценоза [23, 24].
Анализ морфоструктуры каудекса растений видов рода Heracleum показал, что ежегодно вегетативный побег удлиняется и формирует кольцо из листовых рубцов, подсчитывая которые можно определить возраст растений [12, 20]. По нашим наблюдениям, на складчатой поверхности каудекса H. sosnowskyi редко сохраняются выраженные кольцеобразные годичные группы листовых рубцов. Это ограничивает использование данного метода для оценки реального календарного возраста растений в природных условиях.
Определение возраста трав методом ксилохронологии корня связано с выраженной слоистостью вторичной ксилемы и ее соответствием годичной ритмике развития растений [6]. Необходимым условием применения ксилохронологии является окрашивание водопроводящей ткани корня [4]. Данный метод успешно был апробирован для определения возраста растений H. mantegazzianum [13].
Изучение анатомической структуры главного корня растений H. sosnowskyi выявило наличие годичных колец, образованных приростами вторичной ксилемы корня (см. рис. 2, 3). Это позволяет использовать ксилохронологический подход для оценки календарного возраста растений H. sosnowskyi в лабораторных условиях.
Нашей задачей было найти простой и эффективный способ определения календарного возраста растений H. sosnowskyi, чтобы использовать его в полевых условиях. Для этого мы делали продольные срезы каудекса и подсчитывали число годичных побегов предыдущих лет жизни (годичных приростов), границы которых хорошо выражены в сердцевинной паренхиме (см. рис. 3). Результаты исследований показали, что количество годичных приростов соответствовало числу ксилемных колец в основании главного корня, т.е. числу вегетационных периодов. Этот метод позволяет не прибегать к контрастированию тканей, микроскопии и пригоден для быстрого определения возраста растений в полевых условиях.
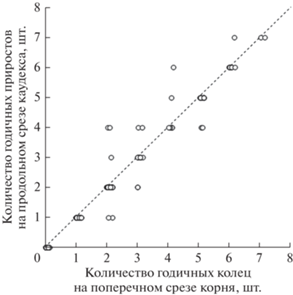
Сопоставление данных о календарном возрасте растений H. sosnowskyi, полученных разными методами, показало, что в 86% случаев (у 90 из 104 разновозрастных особей) наблюдали точное совпадение количества годичных колец на поперечных срезах корня с числом годичных приростов на продольном разрезе каудекса. Обнаружена высокая и статистически значимая коррелированность полученных разными методами данных: r-Пирсона = 0.97, p-value < 0.0001 (рис. 5).
Рис. 5.
Календарный возраст растений Heracleum sosnowskyi, установленный методом ксилохронологии корня (ось абсцисс) и после подсчета годичных приростов на продольном срезе каудекса (ось ординат). Пунктирная линия соответствует полному совпадению результатов определения возраста растений разными методами (n = 90); точки вне линии показывают отклонения в определении возраста растений (n = 14).
Изучение возрастной структуры ценопопуляций H. sosnowskyi показало, что его заросли имеют значительный демографический фонд, состоящий из всходов и ювенильных особей в первые два года жизни (570 ± 333 шт/м2). Этот фонд поддерживается за счет регулярного плодоношения и пополнения почвенного банка семян H. sosnowskyi [24]. В возрасте двух полных лет жизни и старше плотность растений была на порядок ниже. Растения H. sosnowsky поддерживают сравнительно высокий уровень генеративности в возрасте от 2 до 7 лет жизни, что способствует возобновлению ценопопуляций в случае повреждения или гибели части растений. В условиях севера ценопопуляции H. sosnowskyi способны к самоподдержанию в течение длительного времени и продолжают расширять свои площади на пригодных территориях [10, 19].
Таким образом, максимальный возраст особей H. sosnowskyi, подсчитанный разными методами, составляет в среднем 7 лет. Это согласуется с результатами, полученными нами на основе анализа спутниковых изображений модельных участков. Растения приступали к цветению в возрасте 2–6 лет. Доля генеративных особей в разных возрастных группах была высокой и составляла 25%. В искусственных посадках в условиях севера максимальная продолжительность жизни растений H. sosnowskyi была выше и достигала 13 лет [11]. Следовательно, в природных условиях происходило сокращение продолжительности жизни, но при этом сохранялся высокий уровень репродуктивности растений разных возрастов. В оптимальных экологических условиях произрастания растений на севере ценопопуляции H. sosnowskyi возобновляются за счет ежегодного пополнения почвенного банка семян [24]. Это способствует регулярному омоложению ценопопуляций, увеличению скорости их развития, обеспечивает самоподдержание и расселение вида на пригодных территориях в среднетаежной зоне Республики Коми [10, 16, 19].
Анализ возрастной структуры ценопопуляций H. sosnowskyi является необходимым инструментом оценки динамики инвазии – размножения, распространения и захвата новых территорий, и будет содействовать разработке стратегий управления видом.
ЗАКЛЮЧЕНИЕ
Изучение разных методов и подходов к определению календарного возраста растений H. sosnowskyi показало, что метод подсчета листовых рубцов на каудексе ненадежен. Морфоанатомическое исследование выявило наличие годичных колец, образованных приростами вторичной ксилемы корня. Это позволяет использовать ксилохронологический подход для оценки календарного возраста растений H. sosnowskyi. Установлено, что количество ксилемных колец на поперечном срезе корня соответствует числу остатков годичных приростов на продольном срезе каудекса. Предложенный нами новый метод учета годичных приростов на продольном срезе каудекса является простым и эффективным и позволяет определить календарный возраст растений H. sosnowskyi, не прибегая к контрастированию тканей и микроскопии растений. Растения H. sosnowskyi в природных ценопопуляциях приступали к цветению в возрасте 2–6 лет, а их календарный возраст достигал в среднем 7 лет. Это свидетельствует о динамично развивающихся, стабильно омолаживающихся ценопопуляциях H. sosnowskyi в благоприятных экологических условиях на севере.
Исследование выполнено в рамках НИОКТР “Фотосинтез, дыхание и биоэнергетика растений и фототрофных организмов (физиолого-биохимические, молекулярно-генетические и экологические аспекты)” (№ 122040600021-4), поддержано проектом РФФИ “Моделирование потенциальных ареалов инвазионных видов Heracleum sosnowskyi и Heracleum mantegazzianum на территории Восточной Европы” (№ 20-54-18002 Болг_а).
Авторы выражают благодарность и.о. директора Института биологии ФИЦ Коми НЦ УрО РАН, к.б.н. И.Ф. Чадину за обсуждение результатов работы, глубокоуважаемым рецензентам за конструктивные предложения при подготовке рукописи.
Авторы заявляют об отсутствии конфликта интересов.
Настоящая статья не содержит исследований с участием людей и животных в качестве объектов изучения.
Список литературы
Kattge J., Boenisch G., Diaz S. et al. TRY plant trait database – enhanced coverage and open access // Global Change Biology. 2020. V. 26. № 1. P. 119–188. https://doi.org/10.1111/gcb.14904
Работнов А.Т. Методы определения возраста и длительности жизни у травянистых растений // Полевая геоботаника. М.;Л.: Изд-во АН СССР, 1960. Т. 2. С. 249–262.
Жукова Л.А. Многообразие путей онтогенеза в популяциях растений // Экология. 2001. №. 3. С. 169–176.
Dietz H., Ullmann I. Age-determination of dicotyledonous herbaceous perennials by means of annual rings: exception or rule? // Annals of Botany. 1997. V. 80. P. 377–379. https://doi.org/10.1006/anbo.1997.0423
Schweingruber F.H., Poschlod P. Growth rings in herbs and shrubs: life span, age determination and stem anatomy // Forest Snow and Landscape Research. 2005. V. 79. № 3. P. 195–415.
Степанова А.В. Ксилохронология травянистых и полудревесных двудольных растений // Ботан. журн. 2011. Т. 96. № 6. С. 673–680.
Harper J.L., White J.E. The Demography of Plants // Ann. Rev. of Ecology, Evolution, and Systematics. 1974. V. 5. P. 419–463. https://www.jstor.org/stable/2096894
Ценопопуляции растений (основные понятия и структура) / Смирнова О.В., Заугольнова Л.Б., Ермакова И.М. и др. М.: Наука, 1976. 216 с.
Rejmanek M., Richardson D.M. What attributes make some plant species more invasive? // Ecology. 1996. V. 77. P. 1655–1661. https://doi.org/10.2307/2265768
Chadin I., Dalke I., Zakhozhiy I. et al. Distribution of the invasive plant species Heracleum sosnowskyi Manden. in the Komi Republic (Russia) // PhytoKeys. 2017. V. 77. P. 71–80. https://doi.org/10.3897/phytokeys.77.11186
Болотова Е.С. Продолжительность жизни борщевика Сосновского в условиях культуры в центральной зоне Коми АССР // Биологические исследования на северо-востоке европейской части СССР (Ежегодник). Сыктывкар, 1974. С. 54–59.
Сацыперова И.Ф. Борщевики флоры СССР – новые кормовые растения. Л.: Наука, 1984. 223 с.
Pergl J., Perglová I., Pyšek P., Dietz H. Population age structure and reproductive behaviour of the monocarpic perennial Heracleum mantegazzianum (Apiaceae) in its native and invaded distribution ranges // American Journal of Botany. 2006. V. 93. № 7. P. 1018–1028. https://bsapubs.onlinelibrary.wiley.com/doi/full/10.3732/-ajb.93.7.1018
Hüls J., Otte A., Eckstein R.L. Population life-cycle and stand structure in dense and open stands of the introduced tall herb Heracleum mantegazzianum // Biological Invasions. 2007. V. 9. № 7. P. 799-811. https://doi.org/10.1007/s10530-006-9082-9
Nehrbass N., Winkler E., Müllerová J. et al. A simulation model of plant invasion: long-distance dispersal determines the pattern of spread // Biological Invasions. 2007. V. 9. P. 383–395. https://doi.org/10.1007/s10530-006-9040-6
Chadin I., Dalke I., Tishin D. et al. A simple mechanistic model of the invasive species Heracleum sosnowskyi propagule dispersal by wind // PeerJ. 2021. 9:e11821. https://doi.org/10.7717/peerj.11821
Безделев А.Б., Безделева Т.А. Жизненные формы семенных растений российского Дальнего Востока. Владивосток: Дальнаука, 2006. 296 с.
Товстик Е.В., Адамович Т.А., Ашихмина Т.Я. Идентификация участков массового роста борщевика Сосновского с помощью спектральных индексов по данным Sentinel-2 // Теоретическая и прикладная экология. 2019. № 3. С. 34–40. https://doi.org/10.25750/1995-4301-2019-3-034-040
Далькэ И.В., Захожий И.Г., Чадин И.Ф. Распространение борщевика Сосновского и мероприятия по его ликвидации на территории МО ГО “Сыктывкар” (Республика Коми) // Вестник Института биологии Коми НЦ УрО РАН. 2018. № 3(205). С. 2–13. https://doi.org/10.31140/j.vestnikib.2018.3(205).1
Ламан Н.А., Прохоров В.Н., Масловский О.М. Гигантские борщевики – опасные инвазивные виды для природных комплексов и населения Беларуси. Минск: Ин-т экспериментальной ботаники им. В.Ф. Купревича НАН Беларуси, 2009. 40 с.
Барыкина И.П., Веселова Т.Д., Девятов А.Г. Справочник по ботанической микротехнике. Основы и методы: Учеб. пособие. М., 2004. 312 с.
Чадин И.Ф., Далькэ И.В., Малышев Р.В. Оценка морозостойкости борщевика Сосновского (Heracleum sosnowskyi Manden.) после удаления снежного покрова в ранневесенний период // Российский журн. биологич. инвазий. 2018. № 4. С. 105–116.
Tappeiner U., Cernusca A. Model simulation of spatial distribution of photosynthesis in structurally differing plant communities in the Central Caucasus // Ecological Modelling. 1998. V. 113. № 1. P. 201–223. https://doi.org/10.1016/S0304-3800(98)00144-6
Dalke I.V., Chadin I.F., Zakhozhiy I.G. et al. Traits of Heracleum sosnowskyi plants in monostand on invaded area // PLOS ONE. 2015. N. 10 (11). P. e0142833. https://doi.org/10.1371/journal.pone.0142833
Дополнительные материалы отсутствуют.