

Журнал эволюционной биохимии и физиологии, 2019, T. 55, № 6, стр. 455-458
ПРИЧИНЫ УВЕЛИЧЕНИЯ КОЛИЧЕСТВА САМЦОВ МИДИИ МYTILUS GALLOPROVINCIALIS (LAMARK, 1819), КУЛЬТИВИРУЕМОЙ НА ЧЕРНОМОРСКОМ ПОБЕРЕЖЬЕ (КРЫМ, Г. СЕВАСТОПОЛЬ)
Н. С. Челядина 1, *, М. А. Попов 1, Н. В. Поспелова 1, Л. Л. Смирнова 2
1 Институт морских биологических исследований им. А.О. Ковалевского РАН
Севастополь, Россия
2 Институт природно-технических систем
Севастополь, Россия
* E-mail: chelydina2007@mail.ru
Поступила в редакцию 12.09.2018
После доработки 01.02.2019
Принята к публикации 25.04.2019
В последние десятилетия экосистема Черного моря претерпевает значительные изменения природного и антропогенного характера, что приводит к изменению видовой структуры таксоцена моллюсков, трансформируется популяционная структура их массовых видов – половая, размерная, пространственная [1]. Динамика половой структуры тесно связана с эволюционной пластичностью вида, экологической специализацией вида и физиологическим состоянием популяции [2]. Массовым видом двустворчатых моллюсков в Азово-черноморском бассейне является мидия Mytilus galloprovincialis. Ранее показано, что у мидий рода Mytilus в природных популяциях соотношение полов близко к 1:1 [3]. В последние годы в черноморской популяции M. galloprovincialis отмечается сдвиг половой структуры в сторону увеличения количества самцов [4]. Известно, что соотношение самцов и самок в популяции мидий зависит как от генетических механизмов формирования пола, так и от экологических условий среды [5]. У мидий, культивируемых у берегов Крыма, соотношение полов зависит от месторасположения мидийной фермы, при этом количество самцов увеличивается при неблагоприятных условиях среды, техногенной нагрузке [4]. Данные о соотношении полов в популяции моллюсков часто представлены в виде долей самцов и самок взрослых особей без учета их смертности, размера или других особенностей, которые могут потенциально влиять на их соотношение, а дифференциальная смертность в зависимости от пола может искажать соотношение полов у взрослых особей [5]. Информация о смене пола в онтогенезе у М. galloprovincialis практически отсутствует.
Исследование проводили на мидийно-устричной ферме, расположенной на внешнем рейде г. Севастополя (44°37'13.4'' N; 33°30'13.6'' E), и в полузакрытой гавани (44°36'56.4'' N; 33°30'10.6'' E) в период 2015–2018 гг. Ферма хорошо “вентилируется” даже при незначительных ветро-волновых перемешиваниях, относительное содержание кислорода составляет 93–125% от насыщения. Воды в районе фермы классифицированы как мезотрофные [8]. Кормовые условия в акватории морской фермы благоприятны для роста и развития моллюсков. Полузакрытая гавань расположена в 200 м от аварийного выпуска хозяйственно-бытовых сточных вод, сброс которых происходит два-три раза в месяц. В летний период, когда температура воды достигает максимальных значений (до 26.0–29.9°С), внутри гавани относительное содержание кислорода снижается до 60% от насыщения. Расположение фермы и гавани на одной географической широте позволяет пренебречь незначительным перепадом температур 0.1–0.3°С в периоды зимнего выхолаживания и весеннего прогрева.
Целью настоящей работы было выявить причины сдвига соотношения полов и увеличения доли самцов в популяции культивируемой мидии М. galloprovincialis в прибрежных водах юго-западного Крыма.
Мидию M. galloprovincialis с размером раковины 50–60 мм отбирали с экспериментальных веревочных коллекторов фермы (глубина 6 м). Возраст моллюсков (1–1.5 года) определяли по времени экспозиции экспериментального коллектора в акватории фермы. Нерест стимулировали в лабораторных условиях индивидуально для каждой мидии [3]. Пол мидий определяли после нереста с помощью микроскопа Jenaval; гермафродитных особей исключали. В конце апреля 2015 г. (при t воды = 11.9°С) отобранных самок и самцов мидий помещали в отдельные садки (по 100 экз. в каждый) и вывешивали в гавани. В середине апреля 2016 г. (t воды = 11.6°С) экспериментальную работу продолжили только с самками мидий, которых помещали в отдельные садки (по 100 экз. в каждый) и вывешивали на мидийно-устричной ферме и в гавани на глубине 3 м. Продолжительность экспозиции садков с мидиями в море составила 6 мес. (с апреля по октябрь). Соотношение полов в поселениях мидий на дне под коллекторами мидийно-устричной фермы (глубина 15.5 м) и на экспериментальных веревочных коллекторах (глубина 3 м) изучали в июне 2017 г. – после значительного волнения моря (4–5 баллов) и в сентябре 2017 г. и феврале 2018 г. – после длительного слабого волнения (1–2 балла). Стадии зрелости гонад и пол определяли методом визуального изучения мазков под микроскопом Jenaval [3]. Температуру воды измеряли in situ автономным CD-зондом SD204 “SAIV A/S Norway”.
Всего использовано 1600 мидий. Для статистической обработки полученных результатов использовали метод случайных псевдовыборок. Из каждого садка случайным образом отбирали трижды по 33 экз. моллюсков и анализировали каждую выборку отдельно. Для статистического анализа материала использовался пакет программ Microsoft Office Excel 2010. Определяли среднее арифметическое ($\bar {x}$), стандартное отклонение (S), доверительный интервал (Δx). Результаты на рисунках и тексте представлены как $\bar {x} \pm \Delta x$. Достоверность различий оценивали с помощью критерия Стьюдента.
После 6 мес. экспозиции в гавани, загрязненной бытовыми стоками, отмечена высокая смертность моллюсков в обоих садках: смертность самок была выше (48%), по сравнению с самцами (25%). У самцов М. galloprovincialis смену пола не наблюдали. Из оставшихся в живых самок 75% особей поменяло пол (p ≤ 0.01) (рис. 1).
Рис. 1.
Соотношение полов в садках с самками M. galloprovincialis после 6-месячной экспозиции (а – загрязненная акватория, 2015 г.; б – мидийно-устричная ферма, 2016 г.; в – загрязненная акватория, 2016 г.)
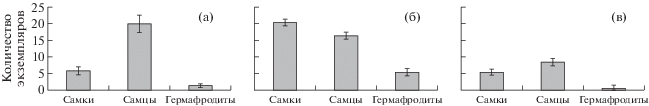
Помимо влияния антропогенного загрязнения, экспериментальные мидии, размещенные в гавани, по-видимому, испытывали недостаток корма в период формирования половых продуктов. Несмотря на то, что моллюски находились на стадии активного гаметогенеза и преднерестовой стадии, гонады мидий обоих полов были тонкие.
Смертность самок мидий на мидийно-устричной ферме в 2016 г. составила 19%, в загрязненной акватории возросла до 69%. Смена пола моллюсков отмечена в обеих акваториях, при этом часть самок стали гермафродитами (p ≤ 0.01). Доля гермафродитов на ферме была выше, по сравнению с гаванью (рис. 1).
Поскольку одной из причин изменения половой структуры мидийной популяции может быть их опадание на дно при штормах и несвоевременном прореживании мидийных коллекторов, было изучено соотношение полов мидий, отобранных одновременно с коллекторов мидийно-устричной фермы и на дне под фермой.
На дне под фермой в июне 2017 г. обнаружены единичные моллюски (длина раковины 60.2 ± 2.2 мм), в сентябре 2017 г. мидии на дне находились в друзах, а в феврале 2018 г. встречались единичные мелкие особи (размер раковины – 40.5 ± 1.2 мм). Среди опавших мидий в июне и феврале преобладали самки, в сентябре – самцы, доля гермафродитов составила 2–6%. На коллекторах фермы в течение всего периода исследований отмечен сдвиг соотношения полов в сторону самцов (табл. 1). Доля гермафродитов составила 5–6%.
Таблица 1.
Соотношения полов в популяции культивируемой мидии M. galloprovincialis в различных условиях обитания (2017–2018 гг.)
| Параметры | Мидийно-устричная ферма | Дно под фермой | ||||
|---|---|---|---|---|---|---|
| Июнь (n = 115) | Сентябрь (n = 110) | Февраль (n = 105) | Июнь (n = 105) | Сентябрь (n = 100) | Февраль (n = 110) | |
| ♀ : ♂ | 1 : 2.9 | 1 : 3 | 1 : 2 | 2.6 : 1 | 1 : 2.2 | 1.8 : 1 |
| Тводы, °С | 24.5 | 23.6 | 7.5 | 11 | 23.1 | 7.3 |
Наши данные показали, что инверсия пола у культивируемой M. galloprovincialis происходит только в одном направлении – от самки к самцу, что может быть одной из причин увеличения численности самцов в поселениях мидий в прибрежной зоне Крыма. Полученные ранее результаты [4] показали, что наибольший сдвиг половой структуры культивируемых мидий в сторону увеличения количества самцов (соотношение – 1 : 7) наблюдался на марихозяйствах, размещенных в акваториях с неблагоприятными гидрохимическими и гидрологическими условиями. Одной из причин сдвига половой структуры в сторону увеличения количества самцов в популяции черноморских мидий может быть низкая выживаемость самок. Предположительно изменение пола связано с наличием пищи: при хороших пищевых условиях моллюски развиваются как самки, при неудовлетворительных условиях питания или при стрессе – как самцы, что подтверждается и нашими результатами. Аналогичные результаты были получены на моллюсках Pinctada margaritifera, Mytella charruana [6]. Потребности в энергетических ресурсах у самцов и самок различны, выработка женских гамет более энергозатратный процесс, чем выработка мужских гамет [6]. Состояние гонад моллюсков перед нерестом косвенно указывает на кормовые условия в течение репродуктивного цикла [7].
Увеличение количества гермафродитов также вносит вклад в соотношение полов. Предполагают, что у раздельнополых видов гермафродиты являются переходной стадией при смене пола [7], что согласуется с нашими данными. Одной из причин изменения соотношения полов в мидийной популяций может быть опадание самок на дно, что, по-видимому, связано с менее прочным биссусным аппаратом особей женского пола. Согласно эволюционной теории пола [2] соотношение полов является регулятором численности популяции, при этом женские особи отвечают за количество, а мужские – за качество потомков. При стабильных оптимальных экологических условиях более важную роль играет количественный аспект (тогда в популяции больше самок), а в период изменяющихся, и даже экстремальных, условий увеличивается количество самцов. В этом случае неблагоприятные условия среды требуют больше мужских особей для быстрого приспособления.
Таким образом, увеличение количества самцов у мидии M. galloprovincialis, культивируемой на черноморском побережье Крыма, может происходить вследствие инверсии пола самок, их повышенной смертности и опадания на дно. При неблагоприятных экологических условиях, в том числе техногенной нагрузке, инверсия пола у самок М. galloprovincialis возрастает.
Список литературы
Черноморские моллюски: элементы сравнительной и экологической биохимии / Под ред. Г.Е. Шульмана, А.А. Солдатова. Севастополь. 2014. [Chernomorskie mollyuski: elementy sravnitel’noj i ekologicheskoj biohimii [Black Sea mollusks: elements of comparative and ecological biochemistry]. Eds. G.E. Shulman, A.A. Soldatov. Sevastopol. 2014 (in Russ).]
Геодакян В.А. Эволюционная теория пола. Природа. 8: 60–69. 1991. [Geodakyan V. A. Evolyucionnaya teoriya pola. [Evolutionary theory of a sex]. Priroda [Nature]. 8: 60–69. 1991 (in Russ).]
Челядина Н.С. Морфологические, биохимические и химические характеристики мидии Мytilus galloprovincialis lam., культивируемой в Чёрном море: Автореф. канд. дис. Севастополь. 2014. [Chelyadina N.S. Morfologicheskie, biohimicheskie i himicheskie harakteristiki midii Mytilus galloprovincialis lam., kul’tiviruemoj v Chyornom more: Avtoref. kand. Diss. [Morphological, biochemical and chemical characteristics of a mussel Mytilus galloprovincialis lam., cultivated in the Black Sea: PhD dissertation abstract]. Sevastopol. 2014 (in Russ).]
Пиркова А.В. Размножение мидии Mytilus galloprovincialis Lam. и элементы биотехнологии ее культивирования: Автореф. канд. дис. Севастополь. 1994. [Pirkova A.V. Razmnozhenie midii Mytilus galloprovincialis lam. i elementy biotekhnologii eyo kul’tivirovaniya: Avtoref. kand. Diss. [Reproduction of a mussel of Mytilus galloprovincialis lam. and elements of biotechnology of its cultivation: PhD dissertation abstract]. Sevastopol. 1994 (in Russ).]
Yusa Y., Breton S., Hoeh W.R. Population Genetics of Sex Determination in Mytilus Mussels: Reanalyses and a Model. Journal of Heredity. 1–7. 2013.https://doi.org/10.1093/jhered/est014
Teaniniuraitemoana V., Leprêtre M., Levy P., Vanaa V., Parrad S., Gaertner-Mazouni N., Gueguen Y., Huvet A., Le Moullac G. Effect of temperature, food availability, and estradiol injection on gametogenesis and gender in the pearl oyster Pinctada margaritifera. J. Exp. Zool. A. Ecol. Genet. Physiol. 325A: 13–24. 2015.https://doi.org/10.1002/jez.1992
Chávez-Villalba J., Soyez C., Aurentz H., Le Moullac G. Physiological responses of female and male black-lip pearl oysters (Pinctada margaritifera) to different temperatures and concentrations of food. Aquat. Living Resour. 26 (3): 263–271. 2013.
Рябушко Л.И., Поспелова Н.В., Балычева Д.С., Ковригина Н.П., Трощенко О.А., Капранов С.В. Исследования микрофитобентоса эпизоона Мytilus galloprovincialis Lam., фитопланктона и гидролого-гидрохимических характеристик акватории мидийной фермы (Севастополь, Черное море). Морской биологический журнал. 2 (4): 67–83. 2017. [Ryabu-shko L.I., Pospelova N.V., Balycheva D.S., Kovrigina N.P., Troshchenko O.A., Kapranov S.V. Epizoon microalgae of the cultivated mollusk Mytilus galloprovincialis lam. 1819, phytoplankton, hydrological and hydrochemical characteristics in the mussel-and-oyster farm area (Sevastopol, Black sea). Marine Biological Journal. 2 (4): 67–83. 2017 (in Russ).]
Дополнительные материалы отсутствуют.
Инструменты
Журнал эволюционной биохимии и физиологии