

Журнал эволюционной биохимии и физиологии, 2020, T. 56, № 1, стр. 62-72
ОТНОСИТЕЛЬНАЯ РОЛЬ АМПЛИТУДЫ СИГНАЛА И СКОРОСТИ ЕЕ ИЗМЕНЕНИЯ ДЛЯ ГЕНЕРАЦИИ ИМПУЛЬСНОЙ АКТИВНОСТИ В НЕЙРОНАХ ПРОДОЛГОВАТОГО МОЗГА АМФИБИЙ
Н. Г. Бибиков *
АО Акустический институт имени акад. Н.Н. Андреева
Москва, Россия
* E-mail: nbibikov1@yandex.ru
Поступила в редакцию 10.03.2017
После доработки 07.12.2018
Принята к публикации 20.02.2019
Аннотация
В полностью адаптированном режиме исследовали ответы одиночных нейронов дорсального ядра продолговатого мозга травяной лягушки (Rana t. temporaria L.) на длительные тональные сигналы, модулированные по амплитуде повторяющимися отрезками низкочастотного шума. Ответ нейрона на модулированный шумом сигнал сопоставляли с текущими мгновенными значениями амплитуды сигнала и скорости ее изменения. Выявлено наличие нейронов, ответ которых определяется текущей амплитудой сигнала, скоростью нарастания амплитуды, а также сочетанием этих двух параметров. Для значительного большинства исследованных клеток основным параметром, вызывающим генерацию спайка, являлась скорость нарастания амплитуды. Проведено сопоставление данных с особенностями реакции нейронов кохлеарного ядра млекопитающих.
ВВЕДЕНИЕ
При анализе сигналов в слуховом пути позвоночных нейроны стволовых отделов мозга осуществляют выделение особенностей временно́го течения сигнала в определенных частотных каналах. Для изучения механизмов, обусловливающих этот анализ, используются различные приемы, большинство из которых основано на методике обратной триггерной корреляции [1]. В рамках этой методики осуществляется синхронная суммация ответов нейрона, соответствующих участкам несущей звукового сигнала или его огибающей, непосредственно предшествующим моменту возникновению спайков. При исследовании тех звеньев слухового пути, в которых воспроизведение несущей частоты ограничено, метод применяется почти исключительно в варианте изучения механизмов кодирования амплитуды огибающей. Это относится к высшим отделам слуховой системы млекопитающих и практически ко всем нейронам холоднокровных животных [2, 3]. В этой методике косвенно предполагается прямая и непосредственная связь ответа нейрона именно с текущей амплитудой сигнала. Так, при исследовании нейронов кохлеарного ядра крысы подчеркивали сходство циклической гистограммы реакции на повторяющиеся отрезки шумовой модуляции с формой огибающей сигнала, прошедшего операцию однополупериодного выпрямления [4–6].
В то же время имеется множество данных, свидетельствующих, что стимулом для возбуждения слуховых нейронов может являться не амплитуда сигнала сама по себе, а некоторые характеристики, описывающие динамику ее изменения. Многочисленные эксперименты, в том числе использующие внутриклеточное отведение [7, 8], показывают, что у многих нейронов слуховой системы спайк возникает только при резком изменении синаптического входа или внутриклеточного импульса тока. Эта особенность наблюдалась даже у волокон слухового нерва и сферических нейронов антеровентрального кохлеарного ядра. Длительный тонический разряд этих элементов при действии немодулированных тональных отрезков достигался вследствие того, что на их немногочисленные синапсы поступали короткие импульсные воздействия, каждый из которых резко изменял значение мембранного потенциала [9]. Поскольку частота следования этих импульсов была монотонно связана именно с амплитудой сигнала, метод триггерной корреляции демонстрировал прямую связь ответа именно с текущим значением амплитуды. Однако в других отделах слуховой системы нейроны могут реагировать и на другие особенности сигнала. Так, клетки-октопусы постеровентрального кохлеарного ядра явно реагируют на крутизну фронта нарастания амплитуды сигнала [8]. В вышележащих отделах слуховой системы вопрос о том, какой временной признак сигнала является наиболее важным для генерации спайковых разрядов нейрона, часто остается открытым.
В настоящей работе была поставлена задача определить, на какие особенности огибающей реагируют клетки ствола мозга в полностью адаптированном режиме. Для этого регистрировали реакцию нейронов дорсального ядра продолговатого мозга амфибий на длительные тональные сигналы, модулированные по амплитуде повторяющимися отрезками сравнительно низкочастотного шума. Такой сигнал уже применялся для получения функции обратной триггерной корреляции, связывающей ответ с мгновенной амплитудой сигнала [10–12]. Однако, исходя из предположения о том, что мгновенная амплитуда сигнала может являться не единственным и даже не основным стимулом, приводящим к генерации спайка, была поставлена задача оценить то, на какую особенность огибающей реагирует та или иная клетка. При этом в качестве таких особенностей временного течения огибающей мы принимали только саму текущую амплитуду и текущую скорость ее изменения.
Хорошо известно, что вследствие распространения возбуждения вдоль слухового пути ответ нейронов запаздывает относительно моментов появления в звуковом сигнале тех его особенностей, которые вызывают спайк. Рассмотрим, как можно оценить эту групповую задержку (латентный период ответа) при действии длительного, амплитудно-модулированного сигнала в адаптированном режиме. Ясно, что латентный период реакции исследуемого нейрона на тональный отрезок может не соответствовать интересующей нас задержке, поскольку уже давно было показано, что ответ на амплитудно-модулированный сигнал в адаптированном режиме весьма далек от ответа того же нейрона на короткий отрезок чистого тона [10, 13–15]. Можно использовать для оценки этой задержки значение интервала, отделяющего момент генерации спайка от максимума функции обратной триггерной корреляции. Однако такой подход также трудно считать адекватным, поскольку само использование функции обратной триггерной корреляции предполагает, что стимулом для генерации спайка является именно мгновенное значение амплитуды. Заметим, что при низкочастотной модуляции нередко отмечалась нереально малая, практически нулевая задержка максимума этой функции относительно максимума амплитуды стимула [11].
Имея это в виду, ранее была разработана методика, позволяющая качественно показать, какие участки огибающей вызывают ответ клетки [12]. Огибающая сигнала была разбита на четыре участка: нарастание амплитуды от среднего значения до максимума, спад от максимума до среднего значения, спад от среднего значения до минимума и нарастание от минимума до среднего значения. Получали значения обратной триггерной корреляции ответа нейрона с каждой из четырех функций, имеющих единичные значения на одном из этих участков и нулевые на всех остальных. Для двух участков, характеризуемых наибольшими максимумами полученных функций, определяли задержки реакции. При этом обнаружилось, что для большинства нейронов как продолговатого, так и среднего мозга участки, оптимальные для генерации спайка, соседствовали друг с другом. Это были либо два участка, при которых амплитуда превышала среднее значение (ответ определяется мгновенным значением амплитуды), либо два участка, где амплитуда нарастала (спайки возникают в моменты роста амплитуды). Обычно интервал между задержками реакции на два избранных участка был близок к одной четверти периода, соответствующего верхней границе частоты модуляции. Во всех случаях задержка реакции на последующий участок была меньше, чем на предшествующий, что естественно объяснялось предположением, что оптимум ответа соответствует границе этих двух участков, то есть либо максимальному значению амплитуды, либо максимуму скорости ее нарастания.
Таким методом качественно можно было выделить нейроны, преимущественно реагирующие на амплитуду сигнала или на скорость ее изменения. Однако представляло интерес оценить вклад этих двух признаков количественно. В данной работе для этой цели мы использовали связь ответа с двумерной функцией, описывающей динамику изменения текущей амплитуды сигнала и ее производной. При этом предполагаемую задержку реакции на выделяемые нейроном особенности сигнала в адаптированном режиме принимали равной среднему значению задержек ответа на пару соседних выделенных участков.
МАТЕРИАЛ И МЕТОДИКА
Поскольку описание используемых сигналов, методы обращения с животным и регистрации импульсной активности уже были приведены ранее [11, 12, 15, 16], мы остановимся на этих аспектах по возможности кратко. Работа была выполнена на 32 самцах травяной лягушки в осенне-зимний период. Перед операцией, во время которой осуществляли дорсальный подход к зоне моста и продолговатого мозга, животные подвергались холодовому или эфирному наркозу в соответствии с общепринятыми правилами обращения с холоднокровными животными [17, 18]. Во время операции края раны дополнительно обрабатывались локальным анестетиком (лидокаин). Во время регистрации нейрональной активности животное, находящееся в звукоизолированной камере, было обездвижено аллоферином.
Акустический сигнал передавали через пластиковый звукопровод от динамического телефона на ухо, ипсилатеральное исследуемому ядру продолговатого мозга. Уровень сигнала в районе барабанной перепонки измеряли конденсаторным микрофоном, калиброванным вместе с насадкой.
Импульсную активность одиночных нейронов регистрировали стеклянными микроэлектродами, заполненными 2.5 М раствором NaCl с кончиком диаметром 1–3 мкм. Электрод перемещали в дорсовентральном направлении с помощью шагового двигателя непосредственно через сосудистое сплетение. Как правило, этот проход осуществлялся без нарушения целостности кончика электрода. Момент входа электрода в дорсальное слуховое ядро продолговатого мозга выявлялся по резкому появлению мультиклеточной нейронной активности в ответ на поисковые звуковые стимулы, которыми служили отрезки широкополосного шума длительностью 100 мс и уровнем 80 дБ УЗД.
После обнаружения одиночного нейрона при достаточном превышении амплитуды его спайков над фоновой активностью (более чем в 5 раз) животным предъявляли короткие (100 мс) тональные отрезки переменного уровня и переменной частоты с целью определения характеристической частоты (ХЧ) нейрона и порога его реакции.
Затем переходили к основному исследованию, при котором сигналом служил тон ХЧ, модулированный по амплитуде повторяющимися отрезками низкочастотного шумового сигнала. Использовали два частотных диапазона модулирующего шума: 0–15 Гц, и 0–50 Гц при длительности каждого из этих повторяющихся отрезков от 512 до 2018 мс. Фиксированные шумовые отрезки модулирующего сигнала следовали один за другим таким образом, что при переходе от одного отрезка к другому переходных процессов не возникало. Обычно каждая сессия включала от 50 до 150 таких повторяющихся отрезков. Несущая частота, которая всегда более чем на порядок превышала максимальную частоту модуляции, не была синхронизована с началом отрезка, и не воспроизводилась в импульсации исследованных нейронов.
Сигнал на телефоны поступал с выхода генератора синусоидальных колебаний Г3-110, имеющего вход для осуществления амплитудной модуляции. Формирование модулирующей функции производили генератором шумов фиксированной формы Г-5-54. Глубина модуляции выражалась в процентном отношении среднеквадратичного значения амплитуды модулирующего шума к среднему значению амплитуды несущей и калибровалась вольтметрами и осциллоскопом.
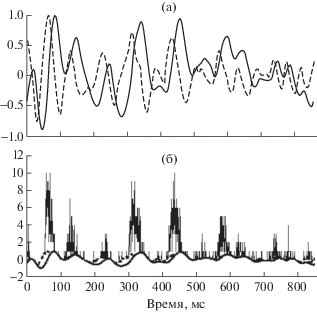
Огибающая каждого из модулирующих шумовых отрезков описывалась с помощью двух нормированных функций: временно́й динамики амплитуды и временно́й динамики ее производной (скорости изменения амплитуды). В качестве примера на рис. 1а показана форма этих функций в случае модуляции шумом в полосе 0–15 Гц на повторяющемся участке длительностью 856 мс. Обе функции нормированы по отношению к значению наибольшего отклонения от среднего. Для сигнала, иллюстрированного рис. 1, наибольшее отклонение амплитуды от среднего наблюдалось на 85 мс и соответствовало локальному максимуму (рис. 1а). Наибольшее изменение скорости в этих условиях также соответствовало максимуму и наблюдалось на 64 мс. Во всех случаях текущие значения амплитуды сигнала и его производной нормировали по отношению к экстремумам, которые принимались за единицу.
Рис. 1.
а – нормированная форма амплитуды огибающей (сплошная линия) и скорости ее изменения (штриховая линия) для одного из повторяющихся отрезков модуляции в полосе 0–15 Гц длительностью 856 мс; б – циклическая гистограмма реакции типичного нейрона дорсального ядра при ее сдвиге на 24 мс (см. текст) на фоне динамики изменения амплитуды (сплошная линия) и скорости изменения (штриховая линия) огибающей. На гистограмме по оси абсцисс – время в мс, по оси ординат – число спайков в интервале длительностью 0.5 мс при общей длительности сигнала 195.5 с (233 повторения отрезка).
Измерения начинали только тогда, когда средняя частота импульсации нейрона стабилизировалась, что обычно наступало через 10–20 с после начала воздействия. Таким образом, регистрация реакции нейрона на тон, модулированный отрезками шума, происходила в режиме практически полной адаптации. В процессе регистрации осуществлялся контроль тренда частоты импульсации, который, как правило, не превышал 20%. Время регистрации ответов на один стимул составляло от 100 до 300 с. При изменении параметров сигнала звук обычно прерывали и после паузы длительностью 10–20 с вновь дожидались устойчивого ответа без выраженного тренда частоты импульсации. В некоторых экспериментах переход от одного типа модулирующей функции к другому происходил без прерывания звучания сигнала. И в этих случаях регистрацию начинали вновь только после достижения устойчивого ответа без выраженного тренда частоты импульсации.
Потенциалы действия регистрируемого нейрона с помощью триггера Шмидта преобразовывались в последовательность стандартных электрических импульсов длительностью 0.5 мс, которые через аналогово-цифровой преобразователь передавались на компьютер для последующей обработки. Непосредственно во время опыта получали функцию, описывающую зависимость плотности импульсации от времени, которую использовали для контроля за отсутствием выраженного тренда частоты импульсации.
Основную обработку данных осуществляли после завершения эксперимента. В процессе этой обработки, проводимой на основе разработанных оригинальных программ, производили построение циклических гистограмм реакции нейрона путем синхронного суммирования спайков, генерируемых им в ответ на последовательно предъявляемые идентичные отрезки модулированного шумом тонального сигнала. Затем в соответствии с методикой, изложенной в работах [12, 16], осуществляли корреляцию ответа с четырьмя участками огибающей (нарастание амплитуды от среднего значения до максимума, спад амплитуды от максимума до среднего значения, спад амплитуды от среднего значения до минимума, рост амплитуды от минимума до среднего значения). После выявления двух участков огибающей, которые характеризовались наибольшей корреляцией с ответом нейрона, для них определяли значение задержки максимума корреляционной функции. Так, для нейрона, иллюстрированного рис. 1, оптимальными являлись те два участка, где амплитуда сигнала превышала среднее значение. Наивысшее значение корреляции огибающей сигнала с ответом нейрона было для участка нарастания амплитуды при постулируемой задержке момента генерации спайка относительно этого участка сигнала на 36 мс. Второй по эффективности оптимальный участок соответствовал спаду амплитуды от максимума к среднему значению при задержке ответа на 12 мс. Далее полученную циклическую гистограмму ответа нейрона смещали назад во времени на величину, равную среднему значению задержек (латентных периодов) ответов на двух оптимальных участках огибающей. При этом максимумы циклической гистограммы должны были совпадать с теми участками огибающей, которые предположительно вызывали ответ клетки. Для нейрона, иллюстрированного рис. 1, значение этого сдвига составило 24 мс. На рис. 1б циклическая гистограмма реакции нейрона приведена уже с учетом указанного сдвига. Из рисунка ясно следует, что генерация спайка приходится, главным образом, на участки сигнала, характеризующиеся превышением амплитуды огибающей над ее средним значением при положительном значении скорости нарастания амплитуды.
Для сравнительной оценки эффективности мгновенной амплитуды и скорости ее нарастания для генерации спайка использовали две процедуры. В соответствии с первой из них получали два численных отношения: 1 – число спайков, генерированных нейроном в те два интервала, когда амплитуда была выше средней, к числу спайков, генерированных на тех двух интервалах, где амплитуда была ниже средней, 2 – число спайков, генерированных на двух участках, когда амплитуда нарастала, к числу спайков, генерированных на двух участках, когда амплитуда спадала. Ясно, что первое отношение указывало на значимость текущей амплитуды для генерации нейроном импульсного разряда, а второе – на значимость текущей скорости ее изменения. Затем эти два значения сравнивали.
Однако при подобной обработке описание огибающей звукового сигнала могло оказаться недостаточно подробным, поскольку при этом не учитывались все текущие значения амплитуды и скорости ее нарастания. Поэтому в соответствии со второй процедурой обработки число спайков циклической гистограммы в каждый момент времени дополнительно умножалось на абсолютные значения отклонения амплитуды от среднего (при оценке роли амплитуды) или соответственно на абсолютные значения скорости нарастания (при оценке роли скорости). Частные, полученные соответственно при положительных и отрицательных значениях амплитуды и скорости нарастания, сравнивали между собой. Также как и при использовании первой процедуры, результатом являлись два числа, характеризующих соответственно эффективность амплитуды и скорости ее изменения для генерации нейроном импульсного разряда.
РЕЗУЛЬТАТЫ
Вначале приведем результаты, полученные с применением указанной методики, для типичного нейрона дорсального ядра, иллюстрированного рис. 1. Для этой клетки после сдвига циклической гистограммы ответа на величину, равную среднему значению задержек ответа на двух оптимальных участках огибающей (24 мс), число спайков, зарегистрированных при амплитуде выше средней, составило 1027, а ниже средней – 334. Число спайков, зарегистрированных при положительном значении производной, составило 1056, а при отрицательном – 305. Очевидно, что эта клетка проявляет предпочтительную реакцию при значениях амплитуды выше средней и при положительных значениях скорости ее нарастания. Сравнительную эффективность скорости нарастания амплитуды и самой амплитуды оценивали путем сопоставления численной величины отношения числа спайков, генерированных при нарастании амплитуды, к числу спайков, возникших на участках уменьшения амплитуды, и отношения числа спайков, генерированных при значениях амплитуды выше средней, к числу спайков, генерированных при амплитуде сигнала ниже средней. Для клетки, иллюстрированной на рис. 1, отношение этих величин составило 3.46 : 3.07 = 1.13, то есть роль скорости изменения была несколько выше, чем роль непосредственно самой текущей амплитуды сигнала.
Если для той же цели использовать не только качественные оценки динамики огибающей, но и текущие абсолютные значения, то отношение числа спайков, зарегистрированных при превышении амплитуды сигнала над средним значением к числу спайков, генерированных при амплитудах ниже среднего, составило 3.39, а для положительных значений скорости нарастания относительно отрицательных значений этого параметра – 5.2. Отношение этих двух значений при таком методе подсчета равно 1.56, то есть значение скорости изменения амплитуды при этом (более точном) методе подчеркивается гораздо сильнее. Таким образом, нейрон, который при качественном рассмотрении, казалось бы, реагирует на участок повышенной амплитуды сигнала, при учете всех текущих значений параметров оказывается скорее детектором скорости изменения амплитуды, чем самой амплитуды.
Количественные данные, касающиеся роли амплитуды сигнала и ее производной, будут приведены, основываясь на данных, полученных вторым из указанных методов. Всего c разной степенью подробности было исследовано 42 нейрона дорсального ядра продолговатого мозга. Все они характеризовались устойчивым ответом в адаптированном режиме. У 24 нейронов наблюдали фоновую импульсации в отсутствие стимулов с частотой более 0.5 имп/с.
У 6 из исследованных клеток не отмечалось выраженной зависимости ответа от формы огибающей. У остальных 36 клеток, эффективно воспроизводящих огибающую стимула, были получены двумерные функции, демонстрирующие зависимость ответа от двух параметров огибающей (амплитуды сигнала и скорости ее изменения). У 12 клеток ответ был исследован при двух значениях частотного диапазона модулирующей функции, у двенадцати – только при частотном диапазоне 0–15 Гц и у двенадцати – только при частотном диапазоне 0–50 Гц. Всего было проведено 48 регистраций. Во всех случаях уровень сигнала составлял 30 дБ над порогом реакции, а глубина модуляции – 20%.
У 22 из 24 клеток, исследованных при частотном диапазоне шума 0–15 Гц, количество спайков, возникающее при превышении амплитуды сигнала над средним значением, было больше, чем при амплитуде сигнала, меньшей среднего значения. Тот же результат был получен и при использовании второго метода, учитывающего абсолютные значения параметров огибающей. Что касается скорости изменения амплитуды, то при использовании первого метода расчета количество спайков, возникающих при нарастании амплитуды, было больше, чем при ее уменьшении также у 22 нейронов, а используя второй метод, мы получили предпочтение к участкам нарастания амплитуды у всех исследованных клеток.
У 21 из 24 клеток, исследованных при частотном диапазоне модулирующего шума 0–50 Гц, число спайков, возникающих при превышении амплитуды сигнала над ее средним значением, было больше, чем при амплитуде сигнала, меньшей среднего значения. Тот же результат был получен и при использовании второго метода, учитывающего абсолютные значения параметров огибающей. Положительные значения скорости изменения амплитуды при использовании первого метода расчета также предпочитал 21 нейрон, но при использовании второго метода предпочтение к участкам нарастания амплитуды наблюдалось у всех исследованных клеток.
Результаты, касающиеся роли амплитуды и ее первой производной в реакции нейронов дорсального ядра, приведены на рис. 2а в логарифмическом масштабе. На точечной гистограмме показана связь отношения числа спайков, возникших при амплитуде сигнала выше средней, к числу спайков, возникших при амплитуде ниже средней (ось абсцисс) и числа спайков, возникших на участках роста амплитуды к числу спайков возникших при ее уменьшении (ось ординат). Видно, что большинство точек расположены выше диагонали, соответствующей равенству этих отношений. Рис. 2б количественно иллюстрирует отношение степени предпочтения нарастания амплитуды к степени предпочтения положительных значений амплитуды. На оси абсцисс данные для 48 регистраций упорядочены в зависимости от степени предпочтения скорости нарастания над значением амплитуды. На оси ординат приведен показатель эффективности скорости изменения амплитуды по сравнению с самой амплитудой сигнала. Черными столбиками отмечены клетки, у которых при использовании методики, использованной в работе [12], оптимальными являлись два участка нарастания амплитуды, а столбиками со штриховкой отмечены случаи, когда при использовании того же метода наблюдалось предпочтение тех двух участков, на которых амплитуда была выше средней.
Рис. 2.
а – взаимозависимость степени предпочтения для возникновения спайка положительных значений текущего значения амплитуды над отрицательными и участков нарастания амплитуды над участками спада. На оси абсцисс указано отношение числа спайков, возникших при амплитуде выше среднего значения к числу спайков, возникших при амплитуде ниже среднего, на оси ординат – отношение числа спайков, возникших при положительной скорости нарастания амплитуды к числу спайков, возникших при отрицательной скорости нарастания амплитуды; б – отношение степени предпочтения скорости нарастания амплитуды к ее текущему значению как показатель эффективности скорости изменения амплитуды для генерации спайка, приведенные для 48 регистраций. Черными столбиками отмечены нейроны, для которых при использовании корреляционного метода оптимальными являлись участки нарастания амплитуды, а столбиками со штриховкой отмечены случаи предпочтения участков, на которых амплитуда была выше средней. По оси абсцисс: номер эксперимента. По оси ординат: показатель эффективности скорости изменения амплитуды для генерации спайка.
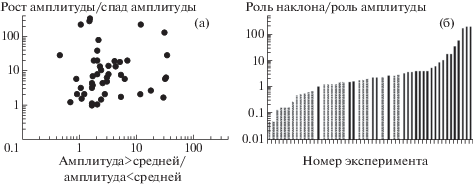
Нетрудно отметить, что хотя качественно данные, полученные этими двумя методами, довольно близки, некоторые нейроны, которые согласно классификации, использованной в работе [12], предпочитали участки с амплитудой выше средней, при учете численных значений этих параметров проявили предпочтение именно к скорости нарастания амплитуды. Всего предпочтение к росту амплитуды по сравнению с значениями самой амплитуды наблюдали в 76% случаев. У некоторых нейронов этот эффект был настолько силен, что при практически полном отсутствии предпочтения к амплитуде сигнала ответ мог возникать только при высоких значениях скорости нарастания.
Рассмотрим несколько конкретных примеров, иллюстрирующих поведение типичных нейронов дорсального ядра, зарегистрированных в данной работе. На рис. 3 полученные гистограммы приведены в трехмерной проекции. Рис. 3а демонстрирует случай, при котором в процессе генерации спайков сохраняется явное предпочтение к амплитудам сигнала, превышающим средние значение огибающей. Но даже и у этого нейрона можно отметить, что импульсы чаще возникают при положительном значении производной.
Рис. 3.
Трехмерные гистограммы зависимости числа спайков в ответе нейронов дорсального ядра (ось z) от текущих нормированных значений амплитуды сигнала (ось x) и ее первой производной (ось y). а – реакция нейрона, имеющего характеристическую частоту 0.54 кГц и порог 40 дБ уровня звукового давления (УЗД), на тон, модулированный шумом в полосе 0–15 Гц, при постулируемой задержке реакции на 18 мс; б – реакция нейрона, имеющего характеристическую частоту 0.64 кГц и порог 32 дБ УЗД, на сигнал, модулированный шумом в полосе 0–15 Гц, при постулируемой задержке реакции на 12 мс; в – реакция нейрона, имеющего характеристическую частоту 0.45 кГц и порог 46 дБ УЗД, на сигнал, модулированный шумом в полосе 0–50 Гц при постулируемой задержке реакции на 12.5 мс; г – реакция нейрона, имеющего характеристическую частоту 0.71 кГц и порог 47 дБ УЗД, на сигнал, модулированный шумом в полосе 0–15 Гц при постулируемой задержке реакции на 16.5 мс. Во всех случаях средний уровень сигнала составлял 30 дБ над порогом, а среднеквадратичное уклонение глубины модуляции шума – 20%. Остальные объяснения в тексте.
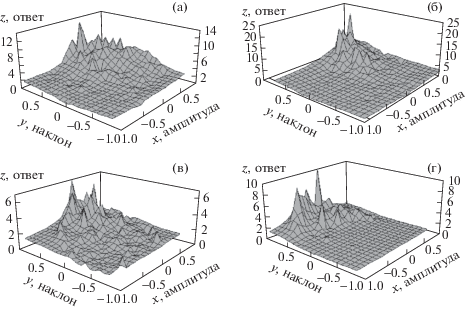
На рис. 3б приведен пример реакции, в которой клетка отвечает исключительно при положительных значениях обоих параметров. Та же тенденция видна и на рис. 3в, хотя в этом случае можно видеть спорадические ответы при отрицательном значении одного из параметров. Наконец, рис. 3г иллюстрирует случай практически полного предпочтения именно скорости нарастания амплитуды.
ОБСУЖДЕНИЕ
Хотя в данной работе мы не осуществляли морфологического контроля расположения электрода, долголетний опыт позволяет определить эту локализацию с большой долей вероятности. В продолговатом мозге лягушки находится еще одно ядро слухового пути – верхняя олива, но оно расположено существенного глубже, чем дорсальное ядро и при переходе от одного ядра к другому всегда имеется зона, в которой регистрируются спонтанно-активные клетки, не реагирующие, однако, на звук. Все исследованные в этой работе нейроны были зарегистрированы непосредственно после вхождения электрода в продолговатый мозг.
Важный вопрос касается справедливости предпосылок, использованных в данной работе для количественного определения относительного вклада собственно текущей амплитуды сигнала и производной этой величины в генерацию импульсов исследуемой клетки. Ясно, что результаты работы зависели, прежде всего, от значения предполагаемой групповой задержки реакции относительно той особенности сигнала, которая вызывает ответ.
В одной из работ, ставивших близкие задачи [19], авторы, работавшие на нейронах коры обезьяны, использовали фиксированную величину предполагаемой задержки реакции, одинаковую для всех исследованных клеток. Такой подход может в определенной мере являться приемлемым для сигнала, где стимулом служил сравнительно короткий отрезок тона с фиксированной по форме модуляцией и резкими нарастанием и спадом амплитуды в начале и в конце. Однако в условиях длительного, непрерывно действующего сигнала предположение об одинаковой задержке реакции для всех нейронов представляется мало обоснованной. Я выбрал для оценки этой задержки данные, полученные на основе исследования реакции именно в адаптированном режиме, когда задержки реакции оценивались путем анализа ответов на характерные участки огибающей. Такой подход представляется более рациональным.
Следует указать на некоторое расхождение полученных в данной работе данных с результатами исследования того же отдела слухового пути амфибий, выполненного путем оценки эффективности разных участков огибающей [12]. В указанной работе было получено, что для большинства нейронов продолговатого мозга оптимальными являлись те два участка огибающей, где амплитуда сигнала превышала среднее значение. Однако при этом отмечалось, что у всех нейронов этой группы наиболее эффективным из двух выделенных участков являлся тот, на котором амплитуда нарастала. Подход, использованный в работе [12], являлся скорее качественным, чем количественным, поскольку конкретные значения текущих параметров сигнала (амплитуда и скорость ее изменения) не учитывались. В данной работе я принимал во внимание нормированные по отношению к максимальной величине численные значения этих двух параметров. При этом наблюдались случаи, когда при качественной оценке оптимальными участками сигнала оставались участки с большой амплитудой, но при этом вклад скорости изменения оказывался большим, чем самой амплитуды (см. рис. 1 и 2б).
Результаты, полученные в настоящей работе, позволяют считать, что для большинства слуховых нейронов второго порядка при воздействии сравнительно низкочастотной амплитудной модуляции в режиме полной адаптации решающим фактором для генерации нейронного импульса является скорость изменения огибающей.
Этот результат следует сопоставить с другими данными, полученными в режиме, близком к полной адаптации, как на нейронах амфибий, так и в кохлеарном ядре млекопитающих. В значительном большинстве этих работ оценивали корреляционные функции между импульсной активностью нейрона и формой сигнала, непосредственно предшествующей появлению импульса, т.е. очевидно вызвавшего генерацию спайка.
В первых работах, выполненных на нейронах кохлеарного ядра мыши [4–6], при анализе активности, вызванной действием тональных сигналов, модулированных низкочастотным шумом, обычно выявлялось две группы клеток. В части нейронов полученная корреляционная функция имела один максимум, что соответствовало возникновению импульса с определенной задержкой в ответ на локальный максимум амплитуды сигнала. Однако в других клетках этому локальному максимуму предшествовал (то есть имел большую задержку на функции обратной триггерной корреляции) локальный минимум. Эти данные можно было интерпретировать двояким образом. Наиболее распространенная интерпретация сводилась к тому, что у нейрона последовательно активировались возбуждающие и тормозные входы. Однако, как уже отмечалось в предыдущей работе [12], вполне возможно интерпретировать эти особенности функции как проявление аккомодационных свойств, когда нейрон отвечает при резком переходе от значений амплитуды ниже среднего уровня к положительным значениям, то есть реагирует на скорость изменения амплитуды.
В большинстве последующих работ, выполненных с использованием метода обратной триггерной корреляции, также выявлялось чередование максимума и минимума, которые, на наш взгляд, следует интерпретировать как проявление дифференцирующих свойств нейронов слуховой системы. Особенно четко это свойство проявлялась у нейронов ангулярного ядра продолговатого мозга птиц [20], которое, являясь гомологом вентрального ядра млекопитающих, имеет много общего и с дорсальным ядром продолговатого мозга амфибий.
Важные данные, касающиеся относительной роли амплитуды сигнала и скорости ее изменения, были получены при изучении бинаурального взаимодействия в другой слуховой структуре ствола мозга млекопитающих – медиальном ядре верхней оливы. Хорошо известно, что нейроны этого ядра являются детекторами совпадения, осуществляя сравнение задержки сигнала на двух ушах. Это положение легло в основу общепринятой модели бинаурального взаимодействия. Однако в работе [21] было показано, что наилучшие результаты в предсказании поведения нейронов медиального ядра могут быть получены, если предположить, что участками огибающей, временно́е совпадение которых приводит к генерации импульса, являются не максимумы амплитуды, а максимумы скорости ее нарастания.
Следует также указать на публикацию, в которой большинство феноменов, связанных с восприятием высоты звука, хорошо объясняется на основании модели, основой которой являются нейроны – детекторы наклона амплитуды [22]. В указанной модели высота определяется как обратная величина от значения максимума функции одномерного распределения межимпульсных интервалов в совокупности нейронов – детекторов наклона огибающей входного сигнала.
В работе [19], о которой уже упоминалось выше, исследовались ответы нейронов слуховой коры бодрствующей обезьяны при действии последовательности изолированных отрезков тона с фиксированной формой амплитудной модуляции длительностью 0.5 с. Авторы разбивали этот отрезок на пять участков разной длины (начальное нарастание, конечный спад, нарастание, спад и постоянство уровня). Оценивалась эффективность каждого из этих участков для возникновения импульса нейрона с предполагаемым фиксированным латентным периодом, равным 15 мс. При таких ограничениях в большинстве клеток оптимальным являлся короткий участок нарастания сигнала, а не участок с максимальной амплитудой. Этот результат вновь свидетельствует о том, что производная огибающей является важнейшим текущим параметром сигнала, вызывающим ответы слуховых нейронов.
Данные, касающиеся роли, которую играют различные участки сигнала в генерации ответа нейрона, были получены и при исследовании иных сенсорных систем. Так, в работе [23] изучалась реакция нейронов церкальной системы сверчка на сравнительно низкочастотные ветровые стимулы. Использовали сигнал, в котором на некотором участке времени амплитуда стимула была постоянна, при том что на других участках постоянным поддерживали скорость ее изменения или даже ускорение. Ответ нейрона наблюдали почти исключительно на участках линейного возрастания скорости. Заметим, что в этой работе исследовали кодирование несущей сигнала, а не его огибающей.
Можно провести качественное сопоставление характеристик исследованных нами клеток с особенностями нейронов, расположенных в основном приемнике волокон слухового нерва млекопитающих – вентральном кохлеарном ядре. В этой структуре описаны клетки, имеющие малое число входов, и эффективно анализирующие тонкую структуру сигнала (сферические и глобулярные клетки), нейроны, анализирующие изменения огибающей (мультиполярные или звездчатые клетки), и фазические нейроны, реагирующие на начало или резкое изменение амплитуды сигнала в широкой полосе частот (клетки-октопусы). Главным образом благодаря работам сотрудников лаборатории биоакустики Висконсинского университета ионные механизмы, определяющие специфику реакции этих групп клеток, были подробно изучены методом внутриклеточной регистрации [7, 8, 24–27]. Сферические клетки при действии внутриклеточного толчка постоянного тока реагируют одним импульсом только на его начало, вследствие того, что входящий натриевый ток резко сменяется выходящим калиевым [8, 25]. Однако поскольку оба этих тока являются крайне быстрыми, то при воздействии импульсного входа от небольшого числа иннервирующих клетку волокон слухового нерва ее ответ носит тонический характер.
Мультиполярные клетки, ионные механизмы которых позволяют воспроизводить динамику синаптического входа, также реагируют тонически, получая множество входов от волокон слухового нерва [7, 25]. Наконец, у клеток октопусов, также получающих входы от большого числа волокон, калиевый ток, присутствующий даже в состоянии покоя, не позволяет возникнуть спайку при медленном нарастании синаптического воздействия [24, 26]. Ответ возникает только в моменты быстрого изменения амплитуды, когда этот выходящий ток не успевает нейтрализовать поступающее возбуждение. В результате для их возбуждения решающим фактором оказывается скорость изменения мембранного потенциала. При действии тональных отрезков они реагируют только на начало сигнала. Качественно подобная классификация применима и для нейронов продолговатого мозга птиц [28].
Важно отметить, что методом внутриклеточной регистрации при фиксации тока было показано, что и в дорсальном ядре продолговатого мозга амфибий могут присутствовать все три типа клеток, описанных для вентрального кохлеарного ядра млекопитающих [29], хотя конкретные ионные механизмы, определяющие эти реакции, охарактеризованы не были.
Данные, полученные в настоящей работе, свидетельствуют о значительном предпочтении нейронами продолговатого мозга участков сигнала с нарастающей амплитудой. Есть все основания полагать, что как у амфибий, так и у млекопитающих этот факт объясняется, главным образом, аккомодационными характеристиками нейронов, определяемыми особенностями ионных токов, прежде всего включением низкопорогового калиевого тока и инактивацией натриевых каналов аксонного холмика.
Еще одним механизмом, также приводящим к подчеркиванию ответа в моменты нарастания сигнала, является рефрактерность исследуемой клетки. Если нейрон генерировал спайк на участке нарастания амплитуды, его ответ в момент максимума и спада амплитуды будет подавлен. Весьма характерный пример такого поведения иллюстрирован на рис. 10 Zhao & Liang [30], когда при больших глубинах модуляции на фазовой гистограмме реакции возникала пауза в момент максимальной амплитуды синусоидально модулированного сигнала.
В настоящей работе особенности сигнала, приводящие к появлению спайка, оценивались на основе оценки задержки его появления относительно временно́го течения огибающей. Другой подход к оценке того, на какой участок сигнала отвечает нейрон, можно получить при исследовании ответа на сигнал, модулированный синусоидой [31, 32], если проанализировать зависимость положения максимума фазовой гистограммы от частоты модуляции. Поскольку эта зависимость обычно представляется линейной, можно оценить точку, в которой полученная прямая пересекает ось ординат. Если это пересечение происходит вблизи нуля, можно сделать вывод, что нейрон воспроизводит именно амплитуду сигнала. При положительных значениях ординаты точки пересечения можно предполагать интегрирующие свойства нейрона, а при отрицательных – дифференцирующие. К сожалению, таких данных в литературе немного, однако некоторые наши результаты [32] также свидетельствовали о важной роли фронта нарастания амплитуды при генерации импульса.
Подчеркнем, что существование двух отдельных наборов элементов, реагирующих соответственно на амплитуду сигнала и на скорость ее нарастания, позволяет расширить диапазон частот, кодируемых временны́м паттерном совокупности нейронов слуховой системы. Данные, приведенные на рис. 2б, позволяют предположить существование подобных специализированных групп среди нейронов второго порядка слуховой системы амфибий.
В данной работе исследовались реакции нейронов на сигналы, модулированные по амплитуде шумом в полосах 0–15 Гц и 0–50 Гц. Такая низкочастотная модуляция во многом определяет восприятие многих коммуникационных звуков, в том числе речи [33]. Для исследуемого объекта полоса 0–15 Гц примерно соответствует временно́му изменению суммарной интенсивности брачного хора, а полоса 0–50 Гц включает в себя частоту амплитудной модуляции изолированного брачного крика самца травяной лягушки [34].
При этом предпочтение нейронов дорсального ядра к высоким скоростям нарастания амплитуды находит соответствие с особенностями временно́го течения основного брачного крика исследуемого объекта. Известно, что этот сигнал состоит из последовательности коротких пульсов с частотой заполнения около 0.6 кГц, следующих с периодом от 20 до 40 мс. При этом передний фронт каждого пульса примерно в два раза короче заднего фронта [34]. Учитывая эти особенности, можно связать выявленный физиологический эффект с известным поведенческим результатом. Животные этого вида при двухальтернативном выборе уверенно предпочитали оригинальный брачный крик сигналу, предъявляемому в обратной временно́й последовательности [3].
Список литературы
De Boer E., Kuyper P. Triggered correlation. IEEE Trans. Biotmed. Eng. 15: 169–179. 1968.
Бибиков Н.Г. Корреляционный анализ кодирования амплитудной модуляции в кохлеарном ядре озерной лягушки. Сенсорные системы. 1: 353–361. 1987. [Bibikov N.G. Correlation analysis of amplitude modulation coding in the cochlear nucleus of the lake frog. Sensory systems. 1: 353–361. 1987. (In Russ).].
van Stokkum I.H.M. Sensitivity of neurons in the dorsal medullary nucleus of the grassfrog to spectral and temporal characteristics of sound. Hear. Res. 29: 223–235. 1987.
Moller A.R. Dynamic properties of the responses of single neurons in the cochlear nucleus. J. Physiol. (L.) 259: 63–82. 1976.
Moller A.R. Neural delay in the ascending auditory pathway. Exp. Brain Res. 43: 93–100. 1981.
Moller A.R. Use of pseudorandom noise in studies of frequency selectivity: The periphery of the auditory system. Biol. Cybern. 47: 95–102. 1983.
Ferragamo M.J., Golding N.L., Oertel D. Synaptic inputs to stellate cells in the ventral cochlear nucleus. J. Neurophysiol. 79: 51–63. 1998.
Golding N.L. Oertel D. Synaptic integration in dendrites: exceptional need for speed. J. Physiol. 590: 5563–5569. 2012.
Rhode W.S. Response patterns to sound associated with labeled globular/bushy cells in cat. Neuroscience 154: 87–98. 2008.
Бибиков Н.Г. Реакции нейронов слухового центра среднего мозга лягушки на тоны, амплитудно-модулированные псевдослучайным шумом. Нейрофизиология. 22: 227–235. 1990. [Bibikov N.G. Reaction of neurons in midbrain auditory center of the frog to tones amplitude-modulated by pseudo-random noise. Neurophysiology. 22: 227–235.1990. (In Russ).].
Бибиков Н.Г. Корреляция нейронных реакций кохлеарного ядра с низкочастотной амплитудной модуляцией тонального сигнала. Акустич. журн. 60(5): 555–566. 2014. [Bibikov N.G. Correlation of neural responses in the cochlear nucleus with low-frequency noise amplitude modulation of a tonal signal. Acoustical Physics. 60(5): 555–566. 2014. (In Russ).].
Бибиков Н.Г. Признаки огибающей звукового сигнала, выделяемые нейронами слухового центра продолговатого мозга лягушки. Биофизика. 60(3): 409–419. 2015. [Bibikov N.G. Some features of sound signal envelope extracted by the frog’s cochlear nucleus neurons. Biofizika. 60(3): 506–518. 2015. (In Russ).].
Бибиков Н.Г. Выделение амплитудно-модулированных отрезков в непрерывном тоне нейронами слуховой системы лягушки. Акустич. журн. 34: 400–401. 1988. [Bibikov N.G. Extraction of amplitude-modulated segments in a continuous tone by auditory neurons of the frog. Soviet Physics. Acoustics. 34: 400–401. 1988. (In Russ).].
Zheng Y., Escabí M.A. Distinct roles for onset and sustained activity in the neuronal code for temporal periodicity and acoustic envelope shape. J. Neurosci. 28(52): 14230–14244. 2008.
Бибиков Н.Г. Количественная оценка изменения синхронизации реакции нейронов кохлеарного ядра лягушки с огибающей звукового сигнала в процессе долговременной адаптации. Акустич. журн. 54: 669–681. 2008. [Bibikov N.G. The quantitative estimation of changes in the synchronization of the neuronal responses in the cochlear nucleus of the frog to the envelope of the signal in the process of long-term adaptation. Acoustical Physics. 54: 597–607. 2008. (In Russ).].
Бибиков Н.Г. Выделение некоторых особенностей низкочастотной огибающей тонального сигнала нейронами слухового центра среднего мозга лягушки. Сенсорные системы. 30: 201–214. 2016. [Bibikov N.G. Extraction of the low-frequency envelope features of the tone by neurons located in midbrain auditory center of the frog. Sensory systems. 30: 201–214. 2016. (In Russ).].
Suckow M.A., Terril L.A., Grigdesby C.F., March P.A. Evaluation of hypothermia-induced analgesia and influence of opioid antagonists in leopard frogs (Rana pipiens). Pharmacol. Biochem. Beha. 63: 39–43. 1999.
Zimmermann M. Ethical principles for maintenance and use of animals in neuroscience research. Neurosci. Lett. 73: 1. 1987.
Zhou Y., Wang X. Cortical processing of dynamic sound envelope transitions. J. Neurosci. 30: 16741–16754. 2010.
Steinberg L.J., Peña J.L. Difference in response reliability predicted by spectrotemporal tuning in the cochlear nuclei of barn owls. J. Neurosci. 31: 3234–3242. 2011.
Gai Y., Kotak.C., Sanes D.H., Rinzel J. On the localization of complex sounds: temporal encoding based on input-slope coincidence detection of envelopes. J. Neurophysiol. 112: 802–813. 2014.
Huang C., Rinzel J. A neuronal network model for pitch selectivity and representation. Front. Comput. Neurosci. 16 June 2016 http: .dx.doi.org/.https://doi.org/10.3389/fncom.2016.00057
Лапшин Д.Н., Рожкова Г.И., Веденина В.Ю. О параметрах стимула, определяющих реакции церкальных интернейронов. Сенсорные системы. 11: 176–180. 1997. [Lapshin D.N., Rozhkova G.I., Vedenina.Yu. On the stimulus parameters determining responses of cercal interneurons. Sensory systems. 11: 176–180. 1997. (In Russ).].
Ferragamo M.J., Oertel D. Octopus cells of the mammalian ventral cochlear nucleus sense the gain of depolarization. J Neurophysiol. 87: 2262–2270. 2002.
Wu S.H., Oertel D. Intracellular injection with horseradish peroxidase of physiologically characterized stellate and bushy cells in slices of mouse anteroventral cochlear nucleus. J Neurosci. 4: 1577–1588. 1984.
Oertel D., Bal R, Gardner S.M, Smith P.H., Joris P.X. Detection of synchrony in the activity of auditory nerve fibers by octopus cells of the mammalian cochlear nucleus. Proc. Nat. Acad. Sci. U S A 97: 11773–11779. 2000.
McGinley M.J., Oertel D. Rate thresholds determine the precision of temporal integration in principal cells of the ventral cochlear nucleus. Hear. Res. 216–217: 52–63. 2006.
Oline S.N., Ashida G. Burger R.M. Tonotopic optimization for temporal processing in the cochlear nucleus. J. Neurosci. 36: 8500–8515. 2016.
Yang S., Feng A.S. Heterogeneous biophysical properties of frog dorsal medullary nucleus (cochlear nucleus) neuron. J. Neurophysiol. 98: 1953–64. 2007.
Zhao H-B., Liang Z.A. Processing of modulation frequency in the dorsal cochlear nucleus of the guinea pig: amplitude modulated tones. Hear. Res. 82: 244–256. 1995.
Вартанян И.А., Гершуни Г.В. Ответы нейронов заднего холма на амплитудно модулированные сигналы при изменении частоты несущей и частоты модуляции. Нейрофизиология. 4: 12–22. 1971. [Vartanian I.A., Gershuni G. Responses of inferior colliculus neurons to the amplitude modulated signals when changing the carrier frequency and the modulation frequency. Neurophysiology. 4: 12–22. 1971. (In Russ).].
Городецкая О.Н., Бибиков Н.Г. Реакции слуховых нейронов продолговатого мозга лягушки на предъявление тонов с синусоидальной модуляцией амплитуды. Нейрофизиология. 17: 390–396. 1985. [Gorodetskaia O.N., Bibikov N.G. The reactions of the auditory neurons in the medulla of the frog brain to bring a tone with a sinusoidal amplitude modulation. Neurophysiology. 17: 390–398. 1985. (In Russ).].
Drullman R., Festen J.M., Plomp R. Effect of reducing slow temporal modulations on speech reception. J. Acoust. Soc. Am. 95: 2670–2680. 1994.
Eggermont J.J., Epping W.J.M. Sensitivity of neurons in the auditory midbrain of the grassfrog to temporal characteristics of sound. III. Stimulation with natural and synthetic mating. Hear. Res. 24: 255–268. 1986.
Дополнительные материалы отсутствуют.
Инструменты
Журнал эволюционной биохимии и физиологии