

Журнал эволюционной биохимии и физиологии, 2020, T. 56, № 1, стр. 73-76
ПОВЕДЕНЧЕСКИЕ И ЭЛЕКТРОКАРДИОГРАФИЧЕСКИЕ РЕАКЦИИ САМОРЕГУЛЯЦИИ ПРИ ДОСТИЖЕНИИ ЦЕЛИ РАЗНОЙ СТЕПЕНИ СЛОЖНОСТИ У ПРИМАТОВ
Т. Г. Кузнецова 1, *, И. Ю. Голубева 1, **, В. Т. Шуваев 1
1 Федеральное государственное бюджетное учреждение науки Институт физиологии им. И.П. Павлова
Российской академии наук
Санкт-Петербург, Россия
* E-mail: Dr.tamara.kuznetsova@gmail.com
** E-mail: GolubevaIU@infran.ru
Поступила в редакцию 03.10.2018
После доработки 08.05.2019
Принята к публикации 22.07.2019
Саморегуляция – один из эволюционных механизмов установления и поддержания физиологических и психологических функций организма на относительно постоянном уровне. В зарубежной литературе реакции саморегуляции рассматриваются как способность к формированию поведенческого торможения или как умение переносить задержку и усилия для получения более ценного результата [1]. В нашем исследовании мы изучаем поведенческие и вегетативные реакции, позволяющие организму осуществлять усилия и выносить задержку при достижении цели, т.е. рассматриваем саморегуляцию с позиций физиологии ВНД, что согласуется с мнением других авторов [2].
Цель работы: проведение сравнительного анализа поведенческих и вегетативных реакций саморегуляции у детей 2–3 и 6–7 лет, шимпанзе и макак при выполнении различных заданий в двух ситуациях: 1) положительный результат достигался быстро, без обучения – “легкое задание”, 2) когда для достижения результата требовались время и обучение – “трудное задание”.
Использовали два методических подхода: 1) достижение положительного результата – реальной цели-объекта с применением методики приближающейся цели, позволяющей градуально изменять напряжение модулирующих систем в зависимости от скоростей движения подкрепления; 2) достижение положительного результата при выполнении заданий с использованием различных когнитивных методик, таких как конструирование, выбор по образцу, выявление информативных признаков изображений, подробно описанных в предыдущих работах [3, 4]. В табл. 1 представлены примеры заданий для разных групп испытуемых.
Таблица 1.
Примеры легких и трудных заданий для разных групп испытуемых
| Испытуемые/ задания | Легкие задания | Трудные задания | ||||
|---|---|---|---|---|---|---|
| Дети 2–3 лет | Конструирование из 2–3 элементов; выбор по образцу реальных геометрических фигур | Конструирование из 5–6 элементов; выбор по образцу абстрактных изображений | ||||
| Дети 6–7 лет | Достижение быстро приближающейся цели (250 мм/с); выявление простых информативных признаков (например, черный/белый) | Достижение медленно приближающейся цели (25–10 мм/с); выявление сложных информативных признаков (например, наличие/отсутствие углов) | ||||
| Шимпанзе | Достижение быстро приближающейся цели (250 мм/с); конструирование из 2–3 элементов; выбор по образцу геометрических фигур и их изображений | Достижение медленно приближающейся цели (25–10 мм/с); конструирование из 5–6 элементов | ||||
| Макаки | достижение быстро приближающейся цели (250 мм/с); выбор по образцу геометрических фигур | Достижение медленно приближающейся цели (25–10 мм/с); выбор по образцу абстрактных изображений | ||||
Работа с разрешения родителей проведена на базе детского сада № 81 Санкт-Петербурга с участием 38 детей 2–3 лет и 39 детей 6–7 лет, и на 7 макак резусов Macaca mulatta (7 ± 2.9 лет) и 14 шимпанзе Pan troglodytes (14 ± 1.4 лет) из биоколлекции “Коллекция лабораторных млекопитающих разной таксономической принадлежности” Института физиологии им. И.П. Павлова, поддержанной программой биоресурсных коллекций ФАНО России.
Во время исследования велась видеозапись поведенческих реакций. Регистрировались: 1) речевые реакции у детей и вокализации у обезьян; 2) пассивное избегание – отведение взгляда, отворот головы и корпуса; 3) двигательная разрядка – монотонные движения, перемена позы и т.п.; 4) реакции на себя – почесывания, прикосновения к различным участкам тела, манипуляции с одеждой у детей; 5) переключение (ориентировочно-исследовательская, игровая деятельность и др.); 6) уходы с рабочего места. Сравнительный анализ количества указанных поведенческих реакций при выполнении легкого и трудного задания проводили с применением критерия Вилкоксона для зависимых выборок.
У детей и шимпанзе дополнительно записывалась ЭКГ во втором стандартном отведении (aVL) с применением усилителя фирмы “Мицар ЭЭГ” (СПб) с портом для кардиоэлектродов (у детей) и диагностического комплекса “Валента” (“Нео”, СПб, 1992) и разработанного нами кардиографического пояса (для шимпанзе) [3]. Анализировали индекс напряжения (ИН = АМо/(2ВР × Мо), где АМо – амплитуда моды, Мо – мода – диапазон наиболее часто встречающихся значений R–R интервалов и ВР – вариационный размах кардиоинтервалов). Для статистической обработки использовался однофакторный критерий ANOVA.
В процессе достижения цели или обучения у испытуемых наблюдались следующие поведенческие реакции саморегуляции: у макак – уходы, двигательная разрядка и пассивное избегание; у шимпанзе доминировали пассивное избегание, двигательная разрядка и реакции на себя, кроме этого присутствовали вокализации и переключения на другие виды деятельности, а у детей в возрасте 2–3 лет в равной степени наблюдались переключения, пассивное избегание, двигательная разрядка, реакции на себя и речевые реакции. Речевую активность детей можно рассматривать как внешнее проявление мыслительного процесса в трудной ситуации [5] или как защитную «оральную» агрессию [6]. У детей 6–7 лет проявление внешних поведенческих реакций сокращалось, что, вероятно, связано с созреванием тормозных процессов и созданием мысленного образа действий. Так, с нашей точки зрения, усложнение и увеличение разнообразия поведенческих реакций саморегуляции от макаки к шимпанзе и ребенку является показателем повышения адаптивных возможностей к психоэмоциональным нагрузкам в филогенезе приматов. Общим для всех испытуемых при усложнении экспериментальной задачи оказалось доминирование двигательной разрядки и пассивного избегания, свидетельствующих о единых филогенетических корнях адаптивного поведения приматов.
Учитывая, что ИН отражает степень напряжения приспособительных возможностей человека, а шимпанзе по интеллектуальному развитию сопоставимы с детьми дошкольного возраста, была проанализирована связь между изменением поведения и ИН у детей 2–7 лет и шимпанзе.
По данным [7] у детей ИН в пределах 90 у.е. характеризует “нормальное” функциональное состояние (норму), при значениях от 90 до 300 у.е. – дистресс, а выше 300 у.е. – срыв механизмов адаптации. Сопоставление поведенческих реакций саморегуляции с уровнем ИН у детей обнаружило взаимосвязь между этими показателями независимо от возраста и используемой методики, и при этом выявились две группы испытуемых. Так, у 70% детей 2–3 лет и 60% детей 6–7 лет при выполнении “легких” заданий он достоверно увеличился относительно фона, достигнув значений, отражающих дистресс (p < 0.001). Выполнение трудных заданий не изменило уровень ИН (рис. 1а, гистограмма, левая панель), но при этом достоверно (p < 0.05) увеличилось общее количество поведенческих реакций (рис. 1а, диаграмма, левая панель). У второй группы детей (30% детей 2–3 и 40% 6–7-летних) ИН уже в исходном состоянии отражал напряжение – дистресс (рис. 1а, гистограмма, правая панель). При выполнении легких заданий, независимо от используемой методики, он удваивался, а при затруднениях достигал 500 у.е. (p < 0.0001), что свидетельствовало о перенапряжения адаптивных резервов этих детей, но при этом у них практически не изменилось поведение (рис. 1а, диаграмма, правая панель). Сопоставление поведения и ИН у шимпанзе показало сходство этих показателей с таковыми у детей (рис. 1б). Следует отметить, что для антропоидов нормативных данных по ИН нет, а частота сердечных сокращений, мода и ее амплитуда сопоставимы с этими показателями у ребенка дошкольного возраста [3]. Этот факт позволил нам считать диапазон значений ИН в пределах до 100 у.е. как “норму”, которую мы наблюдали у 80% шимпанзе. У остальных особей в исходном состоянии ИН достигал 350 у.е., расцениваемое как состояние “дистресса”. Дальнейшее нарастание ИН (p < 0.0001), как и у детей, рассматривалось как перенапряжение адаптивных резервов организма.
Рис. 1.
Динамика ИН (индекс напряжения механизмов сердечного ритма) и поведенческих реакций саморегуляции у детей (1) и у шимпанзе (2) при выполнении легких и трудных заданий. Обозначения: гистограммы – изменение ИН (у.е.) в исходном состоянии (baseline); при выполнении легких (simple) и трудных (difficult) заданий, данные представлены как среднее арифметическое ± стандартная ошибка среднего; лепестковые диаграммы – количество поведенческих реакций (n) при легких (сплошная линия) и трудных (пунктирная линия) заданиях: 1 – речевые реакции у детей и вокализации у шимпанзе; 2 – пассивное избегание; 3 – двигательная разрядка; 4 – реакции на себя; 5 – переключение, отвлечение; 6 – уходы (у шимпанзе).
Однофакторный критерий ANOVA: 1а) p < 0.0001, F2,39 = 87.035; 1б) p < 0.0001, F2,39 = 117.08; 2а) p = 0.0305, F2,18 = 4.263; 2б) p < 0.0001, F2,18 = 43.710.
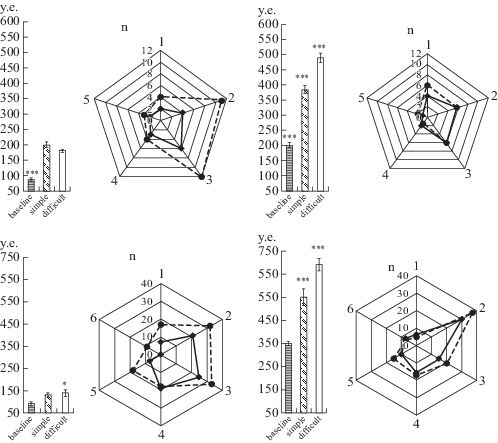
Таким образом, адаптивные реакции саморегуляции при психоэмоциональной нагрузке у высших обезьян и детей независимо от вида, возраста испытуемых и используемых методик могут происходить либо преимущественно за счет видоизменения и увеличения количества поведенческих реакций при мало изменяющемся ИН, либо преимущественно за счет изменения напряжения механизмов регуляции сердечного ритма при мало изменяющихся видимых поведенческих реакций. Полученные результаты демонстрируют общие защитные механизмы приматов к психоэмоциональным нагрузкам.
Список литературы
Beran M.J. The comparative science of “self-control”: what are we talking about? Frontiers in Psychology. 6: 51. 2015.
Yamanashi Y., Matsuzawa T. Emotional consequences when chimpanzees (Pan troglodytes) face challenges: individual differences in self-directed behaviours during cognitive tasks. Animal Welfare. 19(1): 25–30. 2010.
Кузнецова Т.Г., Сыренский В.И., Гусакова Н.С. Шимпанзе: онтогенетическое и интеллектуальное развитие в условиях лабораторного содержания. СПб. Изд. Политехника. 2006. [Kuznetsova T.G., Syrenskij V.I., Gusakova N.S. Shimpanze: ontogeneticheskoe i intellektual’noe razvitie v uslovijah laboratornogo soderzhanija. [Chimpanzees: ontogenetic and intellectual development in laboratory conditions.] SPb. Izd. Politehnika. 2006 (in Russ)].
Кузнецова Т.Г., Голубева И.Ю. Сравнительный анализ выбора по образцу у приматов. Вестник СПбГУ. Серия 12, “Психология, социология, педагогика”. 2: 108–118. 2014. [Kuznetsova T.G., Golubeva I.Ju. Sravnitel’nyj analiz vybora po obrazcu u primatov [Comparative analysis of matching-to-sample results among primates]. Vestnik SPbGU. Serija 12. “Psihologija, sociologija, pedagogika”. 2: 108–118. 2014].
Выготскй Л.С. Мышление и речь. Изд. Хранитель. Москва. 2008. [Vygotskj L.S. Myshlenie i rech’. [Thinking and speech]. Moscow. Izd. Hranitel’. 2008].
Перлз Ф. Гештальт-семинары. Пер. с англ. М. Институт Общегуманитарных исследований. 2007. [Perls F.S. Gestalt Therapy Verbatim. Gestalt psychology. Michigan University. 1969. (Russ Ed. Perlz F. Geshtalt-seminary. M.: Institut Obshchegumanitarnyh issledovanij. 2007)].
Оценка уровня стресса с помощью анализа вариабельности ритма сердца. 2011. Available at: http://www.vedapuls.ru/stress [Ocenka urovnja stressa s pomoshh’ju analiza variabel’nosti ritma serdca. [Stress assessment using heart rate variability analysis]. 2011. Available at: http://www.vedapuls.ru/stress].
Дополнительные материалы отсутствуют.
Инструменты
Журнал эволюционной биохимии и физиологии