

Журнал эволюционной биохимии и физиологии, 2020, T. 56, № 3, стр. 245-250
СПОНТАННАЯ ЭЛЕКТРИЧЕСКАЯ АКТИВНОСТЬ ФАЛЛОПИЕВЫХ ТРУБ КРЫСЫ
К. В. Казарян 1, Н. Г. Унанян 1, *, А. В. Мкртчян 1
1 Институт физиологии им. Л.А. Орбели, НАН
Ереван, Армения
* E-mail: naira_hunanyan@yahoo.com
Поступила в редакцию 07.12.2018
После доработки 05.01.2020
Принята к публикации 20.01.2020
Аннотация
Проведен сравнительный анализ характеристик спонтанной электрической активности пейсмекерных областей (овариальные и цервикальные зоны) фаллопиевых труб крысы. По сравнению с другими активными отделами миометрия овариальная зона левого рога характеризуется наиболее высокими значениями таких показателей активности, как амплитуда потенциала действия, средняя скорость нарастания пика, продолжительность генеза вспышек активности, частота ритмогенеза, свидетельствующими о высокой степени активности данной зоны. Таким образом, выявлена характерная особенность спонтанной электрической активности овариального отдела левой маточной трубы, что может свидетельствовать о наличии дополнительных ее резервных возможностей для проведения электрической волны возбуждения, обеспечивающей распространение соответствующей ей сократимости из овариальной зоны рога до цервикального локуса.
ВВЕДЕНИЕ
Основная функциональная роль репродуктивного органа (деторождение) обеспечивается контрактильной деятельностью гладкомышечной ткани, которая, в свою очередь, является следствием возникновения спонтанной электрической активности [1–5]. Последняя представлена в виде периодически формирующихся в группы высокочастотных спайк-подобных потенциалов, наблюдаемых как в фаллопиевых трубах, так и в самом теле матки. Вспышки активности у небеременных крыс систематически и асинхронно возникают и затухают в определенных зонах миометрия [1, 6, 7]. Единичные спайки способствуют возникновению контрактуры, однако для поддержания сократимости необходимы многократно возникающие и координированные спайки [8, 9]. Таким образом, каждая контрактура сопровождается предварительно возникающими вспышками активности, которые впоследствии затухают.
Проведенный ранее анализ электрической активности миометрия небеременных крыс показал, что она генерируется с присущими ей показателями и спонтанной контрактурой в конечных зонах (овариальная и цервикальная) маточных труб [10, 11]. В дистальной зоне рога волна электрической активности способна распространяться в двух противоположных направлениях, а в овариальной – преобладает однонаправленная передача импульса вдоль миометрия [10–13].
Фаллопиевы трубы, будучи парными полыми элементами в пределах единого репродуктивного органа, характеризуются идентичными физиологическими свойствами. Вместе с тем каждому из рогов может быть свойственна некая асимметрия, зависящая от таких факторов, как физиологическое состояние всей репродуктивной системы в целом, наличие определенного гормонального фона и т.д. [14]. Проведение сравнительного анализа характеристик пейсмекерной активности крайних отделов обоих маточных рогов поможет решению данного вопроса, чему и посвящена настоящая работа.
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЙ
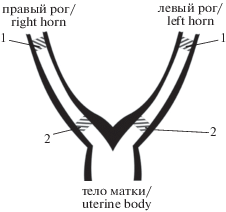
Работа выполнена в условиях in situ на небеременных самках крыс массой 200–250 г. Животные анестезировались нембуталом (40–45 мг/кг) внутрибрюшинно. Вскрывалась брюшная полость и обнажался корпус матки с расположенными с двух сторон маточными трубами. Матка денервировалась перерезкой корешков нервов plexus hypogastricus, uterinus, uterovaginalis [15]. Регистрация активности проводилась одновременно с поверхности разных отделов матки – овариальной и цервикальной областей обоих рогов (рис. 1). Спонтанная электрическая активность отводилась биполярными электродами (межэлектродное расстояние составляло 2 мм). После завершения регистрации животные усыплялись введением дополнительного количества нембутала.
Рис. 1.
Схематическое изображение маточных труб крысы. 1, 2 – соответственно области регистрации из овариальной и цервикальной областей маточных труб.
Fig. 1. Diagrammatic representation of rat fallopian tubes. 1, 2 – recording areas, respectively, in ovarian and cervical regions of rat fallopian tubes.
Анализировали следующие параметры спонтанных потенциалов действия: амплитуда (А), средняя скорость нарастания пика (V), продолжительность нарастания пика (продолжительность увеличения амплитуды потенциала действия до максимального значения) (T), половина ширины (продолжительность формирования верхушки потенциала действия, соответствующей верхней половине его амплитуды) (t), продолжительность вспышки (усредненное значение длительности вспышки активности, наблюдаемое в течение 5 мин регистрации) (L), (частота разрядов активности в 1 мин) (F). Все измерения усреднялись в пределах одного эксперимента, а затем для всех экспериментов данной серии. На рисунках представлены суперпозиции двух типичных форм потенциалов действия. Вычислялся коэффициент, характеризующий скорость формирования пика потенциала действия, соответствующий верхней половине амплитуды (К = А/2:t).
Спонтанная электрическая активность регистрировалась на 6-канальном приборе, разработанном в институте физиологии им. Л.А. Орбели НАН РА для оценки электрической активности гладкой мускулатуры. Отношение сигнал–шум прибора позволяет достоверно регистрировать отклонения сигналов с амплитудой до 10 мкВ. Проводилась полосовая фильтрация регистрируемых сигналов в диапазоне 3–30 Гц. Программа, обеспечивающая регистрацию сигналов, разработана с применением пакета программ Lab View. Статистический анализ проводился с использованием пакетов программ Origin-8.5 и Sigma Plot 11.0. Для оценки достоверности различий применяли t-критерий Стьюдента. Все измерения представлены в процентном отношении к контролю. В экспериментах использованы 18 животных. Рассчитывали также процентное соотношение параметров активности для правого рога по отношению к левому.
Все эксперименты были проведены в соответствии с “Правилами и нормами гуманного обращения с объектами исследования” (NIH publication № 85-23 revised 1985).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
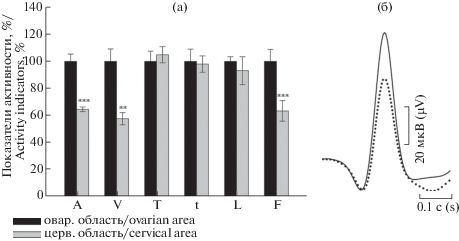
На рис. 2 показаны отдельные вспышки активности соответственно для овариальной и цервикальной областей одного из рогов и рядом показаны развернутые формы единичных потенциалов действия. Полученные нами значения параметров потенциалов действия для всех исследуемых зон свидетельствуют о наличии определенных различий между ними (табл. 1). Анализ изменений исследуемых параметров потенциалов действия овариальной зоны левой маточной трубы по отношению к цервикальному локусу этого же рога (рис. 3) выявил значительное уменьшение амплитуды (до 64.5%, p ≤ 0.001), средней скорости нарастания пика (до 57.5%, p ≤ 0.01) и частоты ритмогенеза вспышек (до 63.3%, p ≤ 0.001). Изменения времени нарастания пика, половины ширины и продолжительности вспышки) находятся в пределах разброса.
Рис. 2.
Вспышки электрической активности из овариальной и цервикальной областей рогов. Справа – соответственно усредненные развернутые формы потенциалов действия.
Fig. 2. Bursts of electrical activity registered from the ovarian and cervical horn areas. On the right – the average unfolded forms of action potentials taken from the respective horn areas.
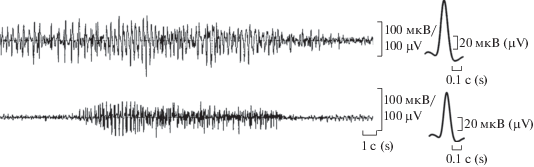
Таблица 1.
Показатели спонтанной активности различных отделов миометрия крысы в норме Table 1. Parameters of spontaneous activity of different myometrial areas in norm
| Области регистрации/ Registration areas | Aмплитуда потенциа-лов действия (A), мкВ/ Amplitude of action potentials (A), μV | Средняя скорость нарастания пика (V), мкВ/с/ Average peak rise rate (V), μV/s | Продолжительность нарастания пика (T), с/Duration of peak rise (T), s | Половина ширины (t), с/ Half width (t), s |
|---|---|---|---|---|
| Овариальная область левого рога/ Ovarian areas of the left horn | 85.1 ± 4.6 | 1306.5 ± 120.0 | 0.07 ± 0.01 | 0.07 ± 0.01 |
| Цервикальная область левого рога/Cervical areas of the left horn | 54.9 ± 1.5 | 751.6 ± 59.2 | 0.08 ± 0.00 | 0.07 ± 0.00 |
| Овариальная область правого рога/ Ovarian areas of the right horn | 58.6 ± 0.7 | 850.4 ± 18.3 | 0.06 ± 0.01 | 0.06 ± 0.01 |
| Цервикальная область правого рога/Cervical areas of the right horn | 55.7 ± 5.5 | 735.4 ± 52.3 | 0.07 ± 0.01 | 0.06 ± 0.01 |
Рис. 3.
(а) Показатели потенциалов действия цервикальной зоны левого рога по отношению к овариальной области левого рога (представлено в процентном соотношении). Контроль соответствует 100%. (б) Наложение друг на друга усредненных форм единичных потенциалов действия овариальной зоны (сплошной контур) и цервикальной области (штриховой контур). n = 18. По оси X – параметры спонтанных потенциалов действия: амплитуда (А), средняя скорость нарастания пика (V), продолжительность нарастания пика (T), продолжительность формирования пика потенциала действия, соответствующей верхней половине его амплитуды) (t), усредненное значение длительности вспышки активности, наблюдаемое в течение 5 мин регистрации (L), частота разрядов активности в 1 мин (F).
Fig. 3. (a) Percentage ratio of the action potential indicators in the left cervical horn area relative to the ovarian. Control corresponds to 100%. (b) Overlaying on each other averaged forms of single action potentials in the ovarian (continuous contour) and cervical (dashed contour) regions. n = 18.
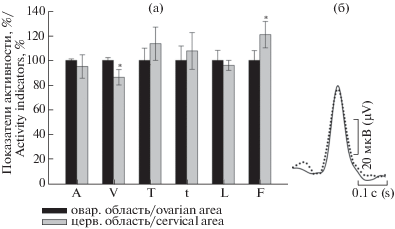
Результаты анализа, проведенного для тех же областей правого рога представлены на рис. 4а. В отличие от левого рога уменьшение величин основных параметров спайка (амплитуда, скорость нарастания спайка) цервикальной области по отношению к овариальной отмечается на небольшую величину. При этом для значений амплитуды потенциалов действия показана лишь тенденция к уменьшению (в пределах разбросов), средняя же скорость нарастания пика понижается лишь на 13.5%, p ≤ 0.05. В отношении половины периода нарастания пика и половины ширины для цервикальной области можно отметить лишь тенденцию к возрастанию (опять таки в пределах разброса). Вместе с тем, если частота вспышек для овариальной области левого рога больше таковой для цервикальной области, то в отношении правого рога отмечается противоположная картина (возрастает на 20%, p ≤ 0.05). Слева на рис. 4б показана картина наложения друг на друга контуров потенциалов действия овариальной и цервикальной зон правого рога.
Рис. 4.
(а) Показатели потенциалов действия цервикальной зоны правого рога по отношению к овариальной области правого рога (представлено в процентном соотношении). Контроль соответствует 100%. (б) Наложение друг на друга усредненных форм единичных потенциалов действия овариальной зоны (сплошной контур) и цервикальной области (штриховой контур). n = 18. Обозначения по оси X как на рис. 3.
Fig. 4. (а) Percentage ratio of the action potential indicators in the right cervical horn area relative to the ovarian. Control corresponds to 100%. (b). Overlaying on each other averaged forms of single action potentials in the ovarian (continuous contour) and cervical (dashed contour) regions. n = 18.
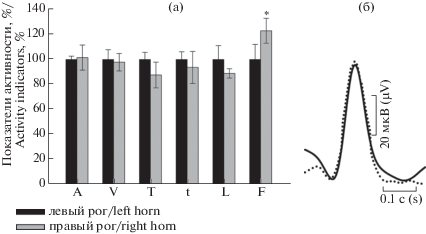
Сравнительный анализ электрофизиологических свойств обеих маточных труб выявил значительные различия между ними (рис. 5а). Суперпозиции потенциалов действия овариальной зоны левого рога по отношению к той же зоне правого рога также демонстрируют изменения их характеристик (рис. 5б). В отличие от овариального локуса, в спайке цервикальной зоны того и другого рогов не обнаружено определенных изменений, лишь несколько учащаются вспышки активности в правом роге (рис. 6а и 6б).
Рис. 5.
(а) Показатели потенциалов действия овариальной зоны правого рога по отношению к овариальной области левого рога (представлено в процентном соотношении). Контроль соответствует 100%. (б) Наложение друг на друга усредненных форм единичных потенциалов действия из овариальной зоны левого рога (сплошной контур) и овариальной области правого рога (штриховой контур). n = 18. Обозначения по оси X как на рис. 3.
Fig. 5. (а) Percentage ratio of action potential indicators in the right ovarian horn area relative to the left ovarian horn area. Control corresponds to 100%. (b) Overlaying on each other averaged forms of single action potentials in the left (solid outline) and right (dashed outline) ovarian horn areas. n = 18.
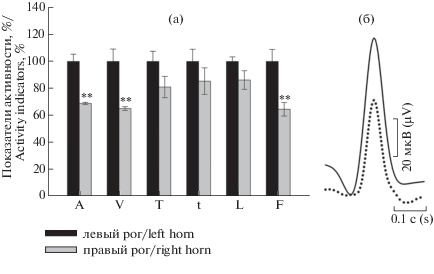
Рис. 6.
(a) Показатели потенциалов действия цервикальной зоны правого рога по отношению к цервикальной области левого рога (представлено в процентном соотношении). Контроль соответствует 100%. (б) Наложение друг на друга усредненных форм единичных потенциалов действия из цервикальной зоны левого рога (сплошной контур) и цервикальной области правого рога (штриховой контур). n = 18. Обозначения по оси X как на рис. 3.
Fig. 6. (a) Percentage ratio of action potential indicators in the right cervical horn area relative to the left cervical horn area. Control corresponds to 100%. (b) Overlaying on each other averaged forms of single action potentials in the left (solid outline) and right (dashed outline) cervical horn areas. n = 18.
Помимо вышеописанных показателей спайковой активности, в работе определены значения коэффициента К – для овариальных зон миометриальной ткани левого и правого рогов относятся соответственно друг к другу как 1.2:1, для цервикальных же областей – как 1:1.2. Таким образом, овариальная зона левого рога имеет наибольшее значение по сравнению со всеми остальными ритмогенными областями, потенциалы действия этой области характеризуются наиболее высокоамплитудными и быстрыми показателями. Подобная особенность показана для всего разряда активности в отношении продолжительности его генеза и частоты ритмики (см. табл. 1). Немаловажную роль в определении изученного типа активности овариальной зоны левого рога может играть локализация специфических пейсмекеров, подобных интерстициальным клеткам Кахаля в миометриальной ткани [3, 16].
Рассматриваемая нами более высокая электрофизиологическая активность левой фаллопиевой трубы может свидетельствовать о дополнительных резервных возможностях для реализации основной функции всего репродуктивного органа. Возможно, левый маточный рог в связи с более высокой электрофизиологической активностью способен при необходимости обеспечивать более усиленную перистальтику всего репродуктивного органа. Таким образом, сравнительный анализ характеристик спонтанной электрической активности крайних ритмогенных областей обеих фаллопиевых труб небеременной крысы показал, что овариальная зона левого рога характеризуется наиболее высокими значениями параметров потенциалов действия и характеристик вспышек активности (продолжительность генез отдельных вспышек, частота их возникновения), определяющих степень возбуждения миометрия [8, 17].
Список литературы
Garfield R.E., Maner W.L. Physiology and electrical activity of uterine contractions. Semin. Cell Dev. Biol. 18 (3): 289–295. 2007.
Kao C.Y. Electrical properties of uterine smooth muscle. In: Biology of the Uterus / Ed. R.M. Wynn, Plenum Press. New York, 423–496. 1977.
Parkington H.C., Coleman H.A. Excitability in uterine smooth muscle. Front Horm Res. 27: 179–200. 2001.
Kuriyama H. Excitation-contraction coupling in various visceral smooth muscles. In: Smooth muscle: an assessment of current knowledge. Ed. E. Bulbring, A.F. Brading, A.W. Jones, T. Tomita, Edward Arnold, London, 1981.
Aguilar H.N., Mitchell B.F. Physiological pathways and molecular mechanisms regulating uterine contractility. Hum. Reprod Update. 16 (6): 725–744. 2010.
Garfield R.E., Daniel E.E., Dukes M., Fitzgerald J.D. Changes of gap junctions in myometrium of guinea pig at parturition and abortion. Can. J. Physiol. Pharmacol. 60 (3): 335–341. 1982.
Казарян К.В., Унанян Н.Г., Саваян А.А., Пилипосян Т.А., Мкртчян А.В., Манукян А.М. Идентификация характеристик спонтанной электрической активности ритмогенных областей миометрия крысы. Ж. эвол. биохим. и физиол. 51 (5): 340–346. 2015. [Kazaryan K.V., Hunanyan N.G., Savayan A.A., Piliposyan T.A., Mkrtchyan A.V., Manukyan A.M. Journal of Evolutionary Biochemistry and Physiology. 51 (5): 388–395. 2015].
Maner W.L., Saade G.R., Garfield R.E. The physiology of uterine contractions. Clin Perinatol. 30 (4): 665–676. 2003.
Hutchings G., Williams O., Cretoiu D., Ciontea S.M. Myometrial interstitial cells and the coordination of myometrial contractility. J Cell Mol Med. 13 (10): 4268–4282. 2009.
Mancinelli R., Guariglia L., Racanicchi C., Bertuzzi A., Salinari S., Vitelli R. In vitro motility of non-pregnant rat uterine horns. Q J Exp Physiol. 73 (4): 459–469. 1988.
Crane L.H., Martin L. Pace-maker activity in the myometrium of the oestrous rat: in vivo studies using video-laparoscopy. Reprod. Fertil. Dev. 3 (5): 519–527. 1991.
Dodds K.N., Staikopoulos, Beckett E.A. Uterine contractility in the nonpregnant mouse: changes during the estrous cycle and effects of chloride channel blockade. Biol Reprod. 92 (6): 141. 2015.
Rabotti C., Mischi M. Propagation of electrical activity in uterine muscle during pregnancy. Acta Physiol (Oxf). 213 (2): 406–416. 2015.
Казарян К.В., Унанян Н.Г., Мкртчян А.В Сравнительный анализ электрофизиологических свойств маточных труб крысы. EESJ 45 (5): 11–15. 2019. [Kaza-ryan, K., Hunanyan N.G., Mkrtchyan A. Camparative analysis of the electrophysiological properties of the rat uterine horns. East European Scientific Journal. 45 (5): 11–15. 2019].
Чухриенко Д.П., Лютко А.В., Романенко Н.Т. Атлас урогинекологических операций (мочеполовая система женщины). Хирургия. Киев. Вища школа. 1981. [Chuhrienko D.P., Lyutko A.V., Romanenko N.T. Atlas uroginekologicheskih operacij (mochepolovaya sistema zhenshchiny) [Atlas urogynecological operations (women’s urogynecological system)]. Hirurgiya. Kiev. Vishcha shkola. 1981 (in Russ.)].
Hirst G.D.S., Ward S.M. Interstitial cells: involvement in rhythmicity and neural control of gut smooth muscle. J. Physiol. 550 (Pt 2): 337–346. 2003.
Sims S.M., Daniel E.E., Garfield R.E. Improved electrical coupling in uterine smooth muscle is associated with increased numbers of gap junctions at parturition. J Gen Physiol. 80 (3): 353–375. 1982.
Дополнительные материалы отсутствуют.
Инструменты
Журнал эволюционной биохимии и физиологии