

Журнал эволюционной биохимии и физиологии, 2020, T. 56, № 5, стр. 391-398
НЕОБЫЧНАЯ КОРРЕЛЯЦИЯ МЕЖДУ РИТМАМИ АКТИВНОСТИ–ПОКОЯ И ТЕМПЕРАТУРЫ ТЕЛА У ГОЛОГО ЗЕМЛЕКОПА (Heterocephalus glaber) В СРАВНЕНИИ С ПЯТЬЮ ДРУГИМИ ВИДАМИ МЛЕКОПИТАЮЩИХ
В. М. Ковальзон 1, *, О. А. Аверина 2, В. А. Минков 3, А. А. Петрин 1, М. Ю. Высоких 2
1 Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
Москва, Россия
2 Институт физико-химической биологии им. А.Н. Белозерского
Московского государственного университета им. М.В. Ломоносова
Москва, Россия
3 Национальный исследовательский университет Высшая школа экономики
Москва, Россия
* E-mail: kovalzon@sevin.ru
Поступила в редакцию 15.04.2020
После доработки 22.05.2020
Принята к публикации 26.05.2020
Аннотация
Температуру тела (ТТ) и двигательную активность (ДА) регистрировали в течение месяца с помощью внутрибрюшинного датчика у следующих шести видов млекопитающих: голых землекопов (Heterocephalus glaber, 2♂, 2♀); лабораторных мышей линии C57Bl/6j (6♂); лабораторных беспородных белых крыс (4♂); монгольских хомячков (Allocricetulus curtatus, 3♂); соболей (Martes zibellina, 2♂, 2♀); лесных хорьков (Mustela putorius, 2♂, 2♀). При этом землекопы находились в своей колонии в особых условиях лаборатории, 2 недели – в постоянной темноте, 2 недели – в режиме чередования светлого и темного периодов (12L/12D). Мыши и крысы находились в индивидуальных камерах в обычных лабораторных условиях в режиме 12L/12D, а соболя и хорьки – в природных вольерах при естественном световом режиме в осеннее время. Все исследованные грызуны, кроме землекопов, являются ночными животными. Соответственно все они демонстрировали четкий циркадианный ритм активности–покоя – высокий уровень ДА в темный период суток и низкий – в светлый. Ритм ТТ коррелировал синфазно с ритмом активности–покоя: подъемы происходили в ночное время, а спады – в дневное (0.9 < r < 1). При этом нарастание ДА в ночное время у хомячков, возникавшее при понижении окружающей температуры, сочеталось с соответствующим подъемом ТТ. Мелкие хищники – соболя и хорьки – демонстрировали более “сглаженный” циркадианный ритм. В отличие от всех этих видов млекопитающих, ТТ землекопов резко падала на 3–5°С (до 28°С) при повышении ДА (обычно в утренние и вечерние часы) и подскакивала (до 36°С) в состоянии покоя (в ночные часы) как в условиях постоянной темноты, так и в режиме 12L/12D (–0.7 < r < –0.9). В целом эти животные демонстрировали оригинальную систему терморегуляции с “установленной точкой”, видимо, около 33.5°С.
ВВЕДЕНИЕ
Голые (точнее, бесшерстные) землекопы (Heterocephalus glaber) – одни из самых удивительных и загадочных наземных млекопитающих [1–3]. Они обитают в аридных зонах Северо-Восточной Африки (Эфиопия, Кения, Сомали), живут под землей большими колониями по нескольку десятков или даже сотен животных. При этом у них, как у пчел, размножается только одна самка (“королева”, или “матка”) огромного размера, которую оплодотворяет один или несколько самцов. Все остальные особи – “рабочие”, вторичные половые признаки у них редуцированы и пол определяется только путем генотипирования. Землекопы не болеют, не стареют, очень долго живут, устойчивы к гипоксии и гиперкапнии и т.д. и поэтому привлекают пристальный интерес представителей самых разных наук [2–6]. Экология, морфология, биохимия, генетика и молекулярная биология этого вида изучены неплохо [1, 3, 5–7], чего нельзя сказать о его физиологии. Последняя в значительной степени основана на единичных отрывочных наблюдениях [8] и даже “легендах”. Так, считается, что голые землекопы не способны поддерживать температуру своего тела, хотя никто этого толком не изучал [9, 10]. Циркадианные ритмы этого животного также изучены крайне слабо [11, 12]. Путем автоматического анализа двухлетних видеозаписей поведения двух колоний землекопов, находящихся в лаборатории в прозрачных лабиринтах, было показано, что в целом у колонии отсутствует циркадианный ритм активности–покоя как в условиях постоянной темноты, так и в режиме 12L/12D. Это означает, что либо он действительно отсутствует у отдельной особи, либо индивидуальные ритмы находятся не в фазе друг с другом. Однако внутри колонии существует синхронизация ритмов активности–покоя, поскольку, когда большинство членов колонии бодрствуют, они мешают спать и всем остальным особям. Рабочие особи проводят в состоянии поведенческого покоя и сна 45–70% времени суток, а “матки”, самые активные члены каждой колонии – 30–40% [11].
В другой работе, весьма обширной и сложно изложенной, изучали как индивидуальные ритмы активности при беге в колесе, так и ритм колонии в целом, с помощью регистрации количества пересечений каждым животным определенного места в лабиринте, где находился регистратор. Были обнаружены очень значительные межиндивидуальные расхождения; тем не менее, было подтверждено, что у подавляющего большинства членов колонии (25 из 29), действительно, циркадианный ритм активности–покоя отсутствует, но при изоляции животных он выявился у 13 из 23 особей, с повышением уровня активности в темный (“ночной”) период в камере [12].
В обоих исследованиях применены лишь косвенные, поведенческие методы определения циркадианной ритмики, которые не могут дать окончательного ответа на вопрос о наличии и характере такой ритмики у особей землекопов. В настоящей работе мы поставили своей целью напрямую ответить на вопрос: отличаются ли околосуточные ритмы активности–покоя и ТТ этих животных от аналогичных параметров других млекопитающих?
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
С этой целью мы вживляли внутрибрюшинно через небольшой разрез (под легким наркозом: голые землекопы – изофлуран, прочие – Золетил 35–40 мг/кг, в/м) экологгеры (миниатюрные автономные датчики температуры и ДА, ООО “ЭМБИ РЕСЕРЧ”, Новосибирск) следующим животным: голым землекопам (2 самца, 2 самки), лабораторным мышам линии C57Bl/6j (6 самцов), лабораторным белым крысам (4 самца), монгольским хомячкам (Allocricetulus curtatus, 3 самца), соболям (Martes zibellina, 2 самца, 2 самки) и хорькам (Mustela putorius, 2 самца, 2 самки). Разрез зашивали, животным вводили антибиотик. Исследование проведено в соответствии с принципами Базельской декларации и одобрено биоэтическим комитетом ИПЭЭ РАН.
Температуру тела (ТТ) измеряли в С°, а двигательную активность (ДА) – в долях G, отклонении ускорения датчика в момент измерения от ускорения свободного падения (т.е., если животное падает в момент измерения, то показатель ДА равен единице). Датчики производили измерения каждую минуту и усредняли их за период, установленный пользователем. В данном исследовании этот период составлял 10 минут. В таком режиме заряда аккумулятора экологгера хватает примерно на 6 нед.
Землекопы поступили из источника в Берлинском зоопарке и были уже не первым поколением, выросшим в неволе. Это были взрослые особи, возрастом около трех лет и весом: самцы – 34 и 47 г, самки – 42 и 52 г. После операции землекопы были помещены обратно в свою колонию, которая содержалась в отдельном лабораторном помещении в специальных лабиринтах из прозрачных плексигласовых цилиндров, диаметром 25 см и высотой 22–30 см, соединенных системой плексигласовых труб диаметром 5 × 4 см. Данные лабиринты имитируют тоннели и камеры обитания голых землекопов в природе. В помещении, в котором расположены лабиринты, поддерживается стабильный микроклимат: температура 27 ± 1.5°С, влажность 50 ± 15%. В качестве подстила используется деревянная щепа. Ежедневно в период с 18:00 до 20:00 животным предоставляют кормовую смесь, состоящую из овощей (батат, морковь, кольраби, корень сельдерея), фруктов (яблоки, груши) и гранулированного автоклавируемого комбикорма, из расчета 6 г на 1 особь в сутки. Воду землекопы не потребляют.
В течение первых двух недель все землекопы содержались в условиях “постоянной темноты”; освещение слабым красным светом (20 лк) включалось только во время раздачи корма, в 18–20 ч. Затем в течение двух недель животные пребывали в условиях “день-ночь”: 12 ч (с 10 до 22 ч) – яркий белый свет (200 лк), 12 ч (с 22 до 10 ч) – слабый красный (12L/12D). Через месяц подопытных животных вновь извлекали из колонии (все животные в колонии чипированы), и под легким наркозом датчики были извлечены через небольшой разрез на коже живота. Рана была зашита, животным введен антибиотик, и они помещены обратно в свою колонию. Данные из накопителя датчика считаны с помощью специального прибора.
Мышей и крыс после операции помещали в индивидуальные камеры, содержавшиеся в отдельном помещении, изолированном от внешних воздействий, при световом цикле 12L/12D, температуре 22–24°С и свободном доступе к воде и пище. Через месяц датчики извлекали, как описано выше. Аналогичную процедуру применяли и ко всем остальным животным. Хомячков в зимний период помещали в индивидуальные камеры, находившиеся в специальной холодильной установке, где животные проходили адаптацию в течение месяца при комнатной температуре (24 ± 1°С) и световом режиме 12L/12D, а затем температуру в камере постепенно понижали до 4 ± 1°С, на 1°С в день. Период освещенности также постепенно сокращали – с 12 (с 9 до 21 ч) до 2 ч (с 14 до 16 ч) в сутки, по полчаса в день. Монгольские хомячки относятся к факультативным гибернаторам, и лишь часть животных погружалась в торпор. Для целей данного исследования были отобраны те животные (3 самца), которые в спячку не погружались. Соболя и хорьки находились в обширных вольерах в полуестественных условиях в лесном массиве на территории биостанции “Черноголовка” в осеннее время.
Полученные кривые ТТ и ДА подвергались математическим процедурам – усреднению, стандартизации и (для соболей и хорьков) экспоненциальному сглаживанию (с коэффицентом 0.3) с определением коэффициента корреляции между ними.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Представленные иллюстрации демонстрируют не только фрагменты нативных кривых, но и результаты их обработки. Все исследованные грызуны, кроме землекопов, являются ночными животными. Соответственно как лабораторные животные, пребывающие в условиях изоляции и искусственной освещенности, они демонстрировали хорошо известные [13] четкие циркадианные ритмы: высокий уровень ДА в темный, “ночной” период суток и низкий – в светлый, “дневной”. Ритм ТТ был синхронен и синфазен с ритмом активности–покоя: подъемы в ночное время (до 39.5°С) и спады – в дневное (до 34.5°С) (рис. 1, а–f).
Рис. 1.
(a) Усредненная динамика температуры тела (верхняя кривая) и двигательной активности (нижняя кривая) у 6 мышей: 6-суточный фрагмент месячной записи, M ± S.E.M. Затемненные участки – ночные периоды в камере (21–09 ч). По оси абсциcc – время суток. По левой ординате – температура брюшной полости в °С. По правой ординате – показания акселерометра (1 = 1g).
(b) Усредненные стандартизированные кривые суточной динамики Т° тела и двигательной активности за весь период эксперимента. По оси абсцисс – время суток; по осям ординат – отклонения от среднего значения обоих параметров.
(c) Точечная диаграмма зависимости Т° тела (по оси абсцисс) от двигательной активности (по оси ординат). По обеим осям – отклонения параметров от их средних значений. Наклонная прямая показывает тренд (0.9< r < 1; p < 0.001).
(d–f) То же, что на a–c, для 4 крыс.
Видно, что увеличение двигательной активности у мышей и крыс (главным образом – в ночное время) сочетается с подъемами температуры тела до 38.5°С, а периоды покоя (преимущественно – в дневное время) сочетаются со снижением температуры тела до 36–36.5°С. Эти колебания явно синхронизируются включениями и выключениями света.
Fig. 1. (a) Averaged dynamics of body temperature (upper curve) and locomotor activity (lower curve) in 6 mice: 6-day fragment of a monthly record, M ± SEM. Shaded areas – night periods in a chamber (9 pm–9 am). Abscissa – the time of day. Left ordinate – temperature of the abdominal cavity (°C). Right ordinate – accelerometer readings (1 = 1 g).
(b) Averaged standardized curves of the daily dynamics of body temperature and locomotor activity over the entire period of the experiment. Abscissa – the time of day; both ordinates – deviations from the average values of both parameters.
(c) Scatter plot of body temperature dependency (abscissa) on locomotor activity (ordinate). Both axes – deviations of the parameters from their average values. The sloping line shows the trend (0.9 < r < 1; p < 0.001).
(d–f) Same as on a–c, for 4 rats.
It is evident that increases in locomotor activity in mice and rats (mainly at night) are accompanied by body temperature elevations up to 38.5°C, while rest or dormancy periods (mainly in the daytime) are accompanied by decreases in body temperature down to 36–36.5°C. These fluctuations are synchronized by turning the light on and off.
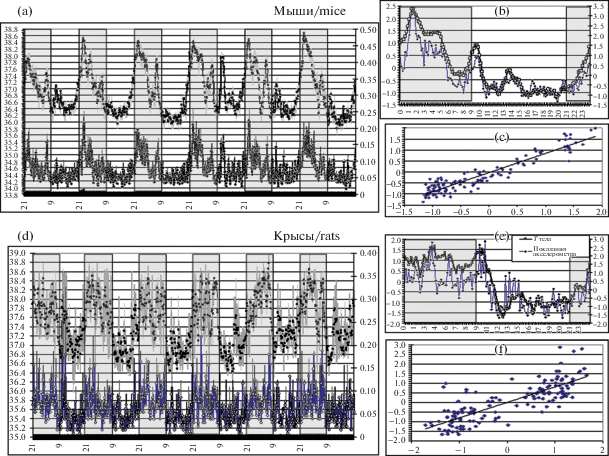
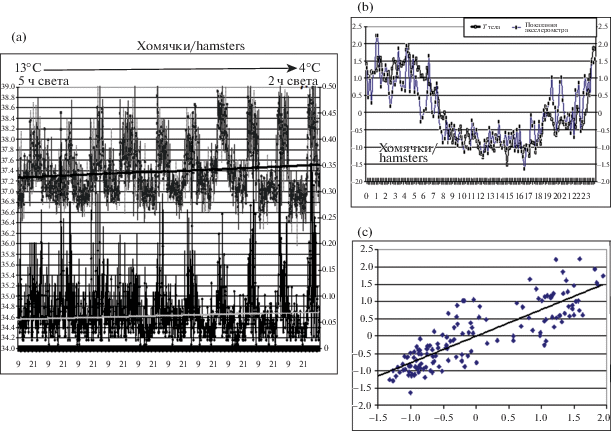
Динамика ТТ в значительной степени связана с интенсивностью мышечной теплопродукции – это особенно ярко проявилось в опытах с хомячками (рис. 2, а–c). Как видно из представленной записи, постепенное понижение внешней температуры привело к значительному повышению ДА в период, соответствовавший “ночному” во время адаптации. Это сочеталось с аналогичным ростом ТТ в ночной период (до 39.2°С). Иными словами, чем ниже была окружающая температура, тем больше бегал хомячок в привычное для него ночное время (несмотря на то, что светлый период все время сокращался) и сильнее разогревался, что позволяло ему сохранять тепло в дневной период покоя. Коэффициент корреляции между показателем ДА и ТТ у всех исследованных грызунов был близок к единице (0.9 < r < 1; p < 0.001).
Рис. 2.
(a) Усредненный 10-суточный фрагмент 40-дневой записи температуры тела (верхняя кривая, M ± S.E.M.) и двигательной активности (нижняя кривая) у трех монгольских хомячков (Allocricetulus curtatus) в ходе опыта по понижению окружающей температуры. Распределения по осям – как на рис. 1, a. Предварительно животные были адаптированы к окружающей температуре 24°С и световому циклу 12L/12D (свет с 9 до 21 ч). В ходе опыта температуру в камере понижали на 1°С в день в течение 20 дней, т.е. до 4°С. Одновременно период освещенности сокращали на 30 мин в сутки, т.е. до 2 ч (14–16 ч). На данном фрагменте графика температура в камере в левой его части была 13°С, а в правой – 4°С; период освещенности в левой части составлял 5 ч (12 : 30–17 : 30), а в правой – 2 ч. Это отмечено горизонтальной стрелкой в верхней части графика.
Видно, что понижение внешней температуры сопровождается повышением двигательной активности и соответственно нарастанием подъемов температуры тела (показаны тренды). Также видно, что все эти подъемы температуры и двигательной активности происходят в привычный для животных ночной период, несмотря на постепенное сокращение реального светового периода в камере.
(b, c) То же, что на рис. 1, b, 1, c (0.9 < r < 1; p < 0.001).
Fig. 2. (a) Averaged 10-day fragment of a 40-day record of body temperature (upper curve, M ± SEM) and locomotor activity (lower curve) in three Mongolian hamsters (Allocricetulus curtatus) during the experiment on lowering the ambient temperature. Axial distributions – as in Fig. 1a. Animals were pre-adapted to an ambient temperature of 24°C and a 12L/12D cycle (light from 9 am to 9 pm). During the experiment, the temperature in the chamber was lowered by 1°C a day for 20 days, i.e. down to 4°C. At the same time, the illumination period was shortened by 30 min a day, i.e. down to 2 h (2 pm–4 pm). On this graph fragment, the chamber temperature was 13°С in its left part and 4°С in the right; the illumination period was 5 h (0 : 30 pm–5 : 30 pm) in the left part and 2 h in the right. This is indicated by a horizontal arrow at the top of the graph.
It is evident that a decrease in the external temperature is accompanied by an increase in locomotor activity and, accordingly, an increase in body temperature elevations (trends are shown). It is also evident that all these elevations, both of temperature and locomotor activity, occur during the nocturnal period, to which all the animals are accustomed, despite a gradual shortening of the real light period in the chamber.
(b, c) Same as in Fig. 1b, 1c (0.9 < r < 1; p < 0.001).
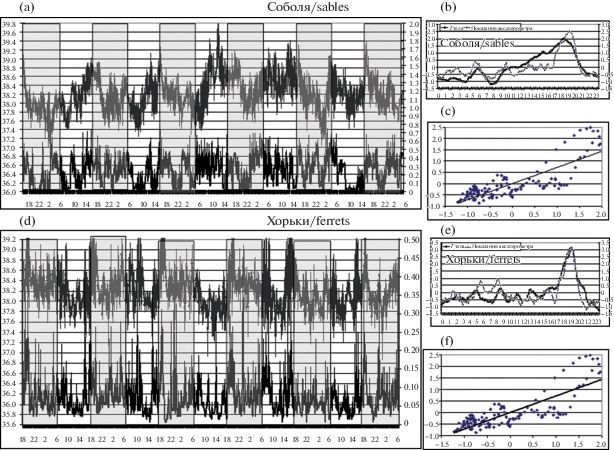
У мелких хищников – соболей и хорьков – циркадианная ритмика выглядела менее выраженной и менее зависимой от светового цикла, отчасти, вероятно, из-за того, что они находились под воздействием разнообразных обонятельных и звуковых стимулов, приходящих к ним из окружающего леса (рис. 3, а–f). Циркадианные кривые хорьков указывают на некоторое преобладание ночной активности, а соболей – скорее на вечерний. Корреляция между обоими регистрируемыми показателями на нативных кривых практически отсутствовала. Однако детальный математический анализ всего периода регистрации с использованием метода экспоненциального сглаживания позволил выявить внутреннюю периодичность циркадианных кривых с основной акрофазой около 20 ч, отражающей, видимо, период кормления животных. Также была выявлена довольно высокая корреляция обеих сглаженных циркадианных кривых (0.7 < r < 0.8). На слабо выраженный характер циркадианного ритма активности–покоя у европейского хорька (Mustela furo), в отличие от выраженного ритма у золотистого хомячка (Mesocricetus auratus), указывали ранее и другие авторы [14].
Рис. 3.
(a) То же, что на рис. 1a–1c, для 4 соболей. (b, с) – то же, что на рис. 1b, 1с, но кривые дополнительно подвергнуты математической процедуре сглаживания.
(d–f) То же для 4 хорьков (0.7 < r < 0.8; p < 0.001).
Fig. 3. (a) Same as in Fig. 1a–1c, for 4 sables. (b, с) – same as in Fig. 1b, 1c, but the curves are additionally subjected to a mathematical smoothing procedure.
(d–f) The same for 4 ferrets (0.7 < r < 0.8; p < 0.001).
Разительный контраст со всеми этими млекопитающими демонстрировали голые землекопы. Как видно из рис. 4, a–e, ТТ у них резко падала на 3–5°С при повышении ДА, практически до уровня окружающей среды (до 28°С) и подскакивала до 36°С в состоянии покоя. При этом все животные явно демонстрировали наличие терморегуляции, стремясь поддерживать ТТ на уровне 33.5°С (видимо, близком к “установленной точке”) – по сравнению с 37–37.5°С у мышей, крыс и хомячков, и 38–38.5°С у соболей и хорьков. Демонстрируемые термограммы явно свидетельствуют в пользу наличия оригинальной системы поддержания температурного гомеостаза у голых землекопов. Такая парадоксальная “отрицательная” связь между ДА и ТТ у голых землекопов была впервые отмечена в работе [8], но ранее не подвергалась систематическому изучению. Каким образом землекопам удается резко охлаждаться при усилении мышечной активности, как бы “вопреки” второму закону термодинамики – остается совершенно загадочным.
Рис. 4.
Голые землекопы (n = 4). (a) То же, что на рис. 1, a. Левая половина рисунка – 3-суточная регистрация в условиях постоянной темноты. Правая половина рисунка – 3-суточная регистрация после перехода в режим 12L/12D.
(b) То же, что на рис. 1, b, в режиме полной темноты.
(c) То же, что на рис. 1, c, в режиме полной темноты (–0.8< r < –0.9; p < 0.001).
(d, e) То же, что на рис. 1, b и рис. 1, c в режиме 12L/12D.
Видно, что температура тела резко падает (до 29–31°С) при усилении двигательной активности животных, главным образом в дневное время. При ослаблении двигательной активности в ночное время температура тела быстро возвращается к нормальным (для данных животных) значениям 33–34°С. Также видно, что включения и выключения света оказывают мало влияния на циркадианные ритмы двигательной активности и температуры тела, которые, по-видимому, синхронизируются другими внешними факторами – появлением людей и раздачей корма.
Fig. 4. Naked mole rats, n = 4. (a) Same as in Fig. 1, a. The left half of the Figure – 3-day registration under conditions of constant darkness. The right half of the figure – 3-day registration after switching to a 12L/12D regime.
(b) Same as in Fig. 1, b in total darkness.
(c) Same as in Fig. 1, c in total darkness (–0.8 < r < –0.9; p < 0.001).
(d, e) Same as on Fig. 1, b and Fig. 1, c under a 12L/12D regime.
It is seen that body temperature drops sharply (down to 29–31°C) when locomotor activity rises, mainly in the daytime. When locomotor activity decreases at night, body temperature quickly returns to normal (for these animals) values of 33–34°С. It is also evident that turning the light on and off has a little effect on circadian rhythms of locomotor activity and body temperature, which appear to be synchronized by the other external factors, e.g., such as the appearance of people coming to feed animals.
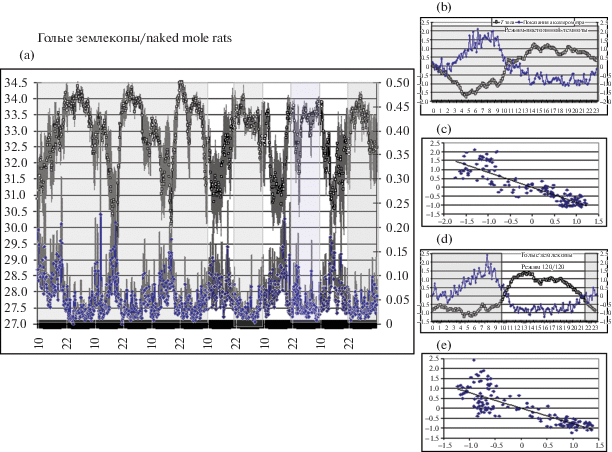
Отрицательная корреляция между показателем ДА и ТТ землекопов была очень высокой (–0.8 < < r < –0.9), преимущественно с двумя циркадианными пиками активности (и соответственно минимальными значениями ТТ): в утренние и вечерние часы. Причем эта ритмика не зависела от светового цикла: она сохранялась и в условиях “постоянной темноты”, и в режиме освещенности 12L/12D. По-видимому, землекопы каким-то образом ощущают присутствие людей – по проникающим к ним (несмотря на предпринятые меры по изоляции) вибрации, шумам, запахам и пр. И это присутствие их беспокоит. Тогда первый пик активности (10–12 ч), вероятно, связан с появлением людей в здании Института, а второй (18–20 ч) – с вхождением сотрудника в комнату, где размещены лабиринты, и раздачей корма. Ночью же, когда в здании никого нет, землекопы в основном находятся в состоянии поведенческого покоя.
ЗАКЛЮЧЕНИЕ
Таким образом, настоящее исследование на грызунах показало, что мыши, крысы и монгольские хомячки в лабораторных условиях являются ярко выраженными ночными животными, и их ночной пик ДА сочетается с подъемом ТТ, а в значительной степени и определяет его. Обитающие в природном окружении мелкие хищники – соболя и хорьки – демонстрируют слабо выраженную циркадианную ритмику, нечетко связанную с изменением освещенности.
Совершенно иной тип циркадианной ритмики демонстрируют голые землекопы – с высоким уровнем ДА в дневное время, независимым от освещенности и сочетающимся, несмотря на мышечную активность, с резким падением ТТ, вплоть до комнатной температуры. По прекращении ДА ТТ быстро поднимается обратно до среднего уровня, видимо, близкого к “установленной точке”. В целом полученные нами результаты являются еще одним свидетельством уникальных биологических характеристик, выработанных этим видом млекопитающих в ходе его эволюции. Несомненно, что изучение физиологических особенностей голых землекопов принесет еще немало сюрпризов.
Список литературы
Browe B.M., Vice E.N., Park T.J. Naked mole-rats: blind, naked, and feeling no pain. Anat. Rec. 303 (1): 77–88. 2020.
Kupferschmidt K. Forever young? Naked mole rats may know the secret. Science. 359 (6375):506-507. 2018.
Skulachev V.P., Holtze S., Vyssokikh M.Y., Bakeeva L.E., Skulachev M.V., Markov A.V., Hildebrandt T.B., Sadovnichii V.A. Neoteny, prolongation of youth: from naked mole rats to “naked apes” (humans). Physiol. Rev. 97: 699–720. 2017.
Buffenstein R. Negligible senescence in the longest living rodent, the naked mole-rat: insights from a successfully aging species. J. Comp. Physiol. B. 178: 439–445. 2008.
Clayson M.S., Devereaux M.E.M., Pamenter M.E. Neurokinin-1 receptor activation is sufficient to restore the hypercapnic ventilatory response in the Substance P-deficient naked mole-rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2020. https://doi.org/10.1152/ajpregu.00251.2019
Schuhmacher L.-N., Husson Z., Smith E. St. John. The naked mole-rat as an animal model in biomedical research: current perspectives. Open Access Animal Physiology. 7: 137–148. 2015.
Kim E.B., Fang X., Fushan A.A., Huang Z., Lobanov A.V., Han L., Marino S.M., Sun X., Turanov A.A., Yang P., Yim S.H., Zhao X., Kasaikina M.V., Stoletzki N., Peng C., Polak P., Xiong Z., Kiezun A., Zhu Y., Chen Y., Kryukov G.V., Zhang Q., Peshkin L., Yang L., Bronson R.T., Buffenstein R., Wang B., Han C., Li Q., Chen L., Zhao W., Sunyaev S.R., Park T.J., Zhang G., Wang J., Gladyshev V.N. Genome sequencing reveals insights into physiology and longevity of the naked mole rat. Nature. 479: 223–227. 2011.
Herold N., Spray S., Horn T., Henriksen S.J. Measurements of behavior in the naked mole-rat after intraperitoneal implantation of a radio-telemetry system. J. Neurosci. Meth. 81: 151–158. 1998.
Buffenstein R., Yahav S. Is the naked mole-rat Heterocephalus glaber an endothermic yet poikilothermic mammal? J. Therm. Biol. 16 (4): 227–232. 1991.
Yahav S., Buffenstein R. Huddling behavior facilitates homeothermy in the naked mole rat Heterocephalus glaber. Physiol. Zool. 64 (3): 871–884. 1991.
Davis-Walton J. Sleep arrhythmia in the eusocial naked mole-rat. Naturwissenschaften. 81: 272–275. 1994.
Riccio A.P., Goldman B.D. Circadian rhythms of locomotor activity in naked mole-rats (Heterocephalus glaber). Physiol. Behav. 71: 1–13. 2000.
Benstaali C., Mailloux A., Bogdan A., Auzéby A., Touitou Y. Circadian rhythms of body temperature and motor activity in rodents. Their relationships with the light-dark cycle. Life Sci. 68: 2645–2656. 2001.
Stockman R., Albers H.E., Baum M.J. Activity in the ferret: oestradiol effects and circadian rhythms. Anim. Behav. 33: 150–154. 1985.
Дополнительные материалы отсутствуют.
Инструменты
Журнал эволюционной биохимии и физиологии