

Журнал эволюционной биохимии и физиологии, 2021, T. 57, № 5, стр. 392-401
СЕЗОННАЯ ДИНАМИКА СИСТЕМЫ КРАСНОЙ КРОВИ МАЛОЙ ЛЕСНОЙ МЫШИ (APODEMUS URALENSIS) В ГОРАХ ЦЕНТРАЛЬНОГО КАВКАЗА
М. М. Емкужева 1, *, Ф. А. Темботова 1, З. А. Берсекова 1, З. Х. Боттаева 1, А. Х. Чапаев 1
1 Институт экологии горных территорий им. А.К. Темботова РАН
Нальчик, Россия
* E-mail: emkugeva_m@mail.ru
Поступила в редакцию 01.12.2020
После доработки 26.02.2021
Принята к публикации 18.03.2021
Аннотация
Исследован комплекс показателей системы крови широко распространенного вида – малой лесной мыши (Apodemus uralensis Pall.) в горах Кавказа (1800 м над ур. м.) с охватом всех сезонов года. Исследование было проведено на взрослых животных, отловленных зимой, весной, летом и осенью в сосновом лесу (Национальный парк Приэльбрусье, КБР). При действии хронической гипоксии у вида показана сезонная динамика эритропоэза костного мозга, поступления молодых эритроцитов (ретикулоцитов) в периферическую кровь и особенности поддержания кислородной емкости крови в разные сезоны года. Кроветворная функция костного мозга A. uralensis в среднегорьях сохраняется на высоком уровне в течение всего годового цикла, при этом наиболее активное кроветворение наблюдается в зимний и летний периоды, и менее активное – в весенний и осенний. Обитание в условиях хронической гипоксии обусловливает необходимость высокого оснащения организма A. uralensis кислородом независимо от сезона года. В способах поддержания кислородной емкости крови в разные сезоны года имеются существенные различия: в холодный осенне-зимний период она обеспечивается увеличением общей поглотительной поверхности для кислорода, в весенне-летний – поступлением в кровоток эритроцитов с качественно иными свойствами. Выявленная сезонная изменчивость в показателях эритрона малой лесной мыши обеспечивает поддержание как оптимальной кислородной емкости крови, так и гомеостаза всего организма в горах Центрального Кавказа.
ВВЕДЕНИЕ
Как известно, гипоксия является одним из главных факторов эволюционной адаптации, которая приводит к появлению большого разнообразия адаптивных стратегий [1]. Исследование физиологических, а в последние десятилетия и молекулярно-генетических основ адаптаций к гипоксии у разных видов, остается одним из актуальных вопросов современной биологии [2–6]. Эти адаптации направлены на увеличение подачи кислорода к тканям и/или снижение потребности в кислороде на клеточном уровне [5, 7]. Адаптивные реакции малой лесной мыши (Apodemus uralensis Pall.) к условиям гор относятся к первому типу. Увеличение пропускной способности кислорода в крови у вида обеспечивается изменениями в эритропоэтической активности костного мозга и соответствующим поступлением в периферическую кровь большого числа молодых эритроцитов – ретикулоцитов – и качественными особенностями этих клеток: более высокое содержание гемоглобина при меньших размерах [8]. В результате высокой кислородной емкости крови увеличивается нагрузка на сердце, что выражается в относительно больших размерах сердца [8].
Горные условия характеризуются комбинированным воздействием гипоксии и холода, что создает серьезные физиологические проблемы для млекопитающих, поскольку реакция метаболизма на эти два фактора противоположна [5, 9]. Эти стрессовые факторы накладываются на сезонную цикличность сдвигов условий существования (как климатических, так и кормовых). При значительном объеме исследований по различным аспектам адаптации к гипоксии нет четкого понимания того, каким образом в разные сезоны года обеспечивается поддержание кислородного запроса организма в экстремальных условиях гор.
Цель работы состояла в исследовании особенностей обеспечения кислородного запроса организма широко распространенного вида – Apodemus uralensis, в разные сезоны года в горах Центрального Кавказа.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЙ
Исследования, согласно типизации А.К. Темботова [10], проводись в условиях среднегорий субальпийского пояса эльбрусского варианта Центрального Кавказа. Эльбрусский вариант относится к континентальному подтипу восточно-северокавказского (полупустынного) типа поясности. Здесь Главный и Боковой хребты состоят из больших вершин, в силу чего влажные средиземно-черноморские ветры задерживаются, их влияние здесь отсутствует. Климат континентальный, за счет свободного проникновения сухих воздушных потоков с Прикаспийской низменности в горы. Отлов животных проводили в сосновом лесу с примесью березы на территории Национального парка “Приэльбрусье”, окр. пос. Эльбрус, высота местности – 1800 м над ур. м., GPS координаты – 43°15' с.ш., 42°38' в.д.
Объект исследования (Apodemus uralensis Pall.) – доминирующий вид в сообществе мелких млекопитающих субальпийского пояса эльбрусского варианта. Ареал вида на Кавказе в высотном направлении простирается до 3000 м над ур. м. [11]. Молекулярно-генетическими исследованиями на Центральном Кавказе, в пределах КБР, установлено обитание генетически однородных животных, которые идентифицируются как Apodemus uralensis [12].
В работе использовано 170 особей малой лесной мыши с охватом всех сезонов года: зима (январь) – 11 самцов, 10 самок; весна (апрель) – 22 самца, 12 самок; лето (июль) – 32 самца, 33 самки; осень (октябрь) – 20 самцов, 30 самок. Животных отлавливали стандартным методом ловушко-линий [13]. Изучались взрослые особи, возраст которых определяли на основании комплекса признаков стертости зубов, массы тела, состояния генеративных органов. Работа выполнена с соблюдением этических стандартов, утвержденных правовыми актами РФ, международных принципов Базельской декларации о гуманном отношении к животным и правил проведения работ с использованием экспериментальных животных.
Костный мозг получали из бедренной кости. Препараты костного мозга готовили на предметных стеклах и окрашивали комбинированным методом (по Май-Грюнвальд и Романовскому-Гимза, MiniMed, Россия). Морфологический анализ клеток костного мозга (соотношение клеток лейкоцитарного и эритробластного ряда, эритробластограмма) проводился с помощью микроскопа Axio Imager, (Carl Zeiss, Германия), окуляр – 10, объектив – 100 с масляной иммерсией. Ретикулоциты крови (RET) и их парциальный состав исследовали на микропрепаратах, окрашенных 1% раствором красителя бриллиант-крезилблау. Расчет проводили на 10 тыс. эритроцитов. По степени зрелости различали 5 групп ретикулоцитов (0, I, II, III, IV). Концентрацию гемоглобина (HGB, г/л) определяли унифицированным гемоглобинцианидным методом на гемоглобинометре фотометрическом МиниГЕМ-540 (Техномедика, Россия), гематокритное число – соотношение плазмы и форменных элементов (HCT, об.%) – микрометодом на гематокритной центрифуге CM-70 (ELMI, Латвия), подсчет количества эритроцитов (RBC, млн в 1 мкл) проводился в камере Горяева. Диаметр эритроцитов (микрометры) измерялся прямым микрометрическим методом с помощью винтового окуляр-микрометра МАВ 1-16х на сухих окрашенных препаратах крови. Измеряли диаметр 100 различных эритроцитов, результаты распределяли по группам (по величине диаметра эритроцитов). Рассчитывали среднее содержание гемоглобина в эритроците (MCH) в пикограммах (пг), среднюю концентрацию гемоглобина в эритроците (MCHC) в г/л, средний объем эритроцита (MCV) в кубических микрометрах (мкм3) [14, 15].
Статистическая обработка данных проводилась с использованием пакета программ “Statistica-10 for Windows”. Влияние факторов проанализировано двухфакторным дисперсионным анализом (Factorial ANOVA). При оценке различий (половых, сезонных) использовали Tukey-тест. Кластерный анализ проводили на основе матрицы дистанций Махаланобиса методом Варда (Ward’s method, 1 – Pearson –r). Для всех статистических тестов принят 5% уровень значимости.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Двухфакторный дисперсионный анализ показал, что система крови малой лесной мыши в условиях среднегорья Центрального Кавказа претерпевает значительные сезонные изменения (F = 3.6, p = 0.000), не зависит от пола животных (F = 0.6, p = 0.867) и совместного действия рассматриваемых факторов (F = 1.3, p = 0.104).
Согласно результатам дискриминантного анализа 82% животных соответствуют классификационной матрице, однородность зимней, весенней и осенней выборок составляет 80–88%, летней – 77%.
Система крови A. uralensis в зимний период (январь). Активная часть паренхимы костного мозга у всех изученных особей в зимний период была густо заполнена кроветворными клетками. Эритроидные клетки на препаратах равномерно располагались между другими кроветворными элементами лейкоцитарного ряда и эритроцитами. На каждое поле зрения их приходилось от 15 до 35. Среди этих клеток эритроидного ряда в стадии митоза были 1–3. Жировые клетки встречались в небольшом количестве и не в каждом поле зрения. В парциальном соотношении клеточных групп костного мозга эритроидные клетки составили 21.65% у самцов и 21.45% у самок, оставшиеся около 80% приходятся на лейкоцитарный ряд (табл. 1). В эритрограмме выявлен сдвиг в левую сторону за счет молодых клеток (эритробластов и базофильных нормобластов).
Таблица 1.
Сезонная динамика параметров костного мозга малой лесной мыши (A. uralensis) в условиях среднегорий Центрального Кавказа
| Параметры | пол | M ± SE | p < 0.05 | |||
|---|---|---|---|---|---|---|
| Зима (1) | Весна (2) | Лето (3) | Осень (4) | |||
| Эритробластный ряд в % ко всем ядерным клеткам костного мозга | ♂ | 21.65 ± 0.82 | 18.18 ± 0.63 | 22.59 ± 0.60 | 18.88 ± 0.70 | 1–2, 2–3, 3–4 |
| ♀ | 21.45 ± 0.22 | 18.81 ± 1.34 | 23.47 ± 0.60 | 20.22 ± 1.02 | 2–3, 3–4 | |
| Соотношение лейкопоэз эритропоэз |
♂ | 3.61 : 1 | 4.60 : 1 | 3.50 : 1 | 4.43 : 1 | |
| ♀ | 3.82 : 1 | 4.49 : 1 | 3.34 : 1 | 4.15 : 1 | ||
В периферической крови содержание молодых эритроцитов (ретикулоцитов) высокое и составляет 35.47‰ у самцов и 35.67‰ у самок. В парциальном их составе преимущественное содержание ретикулоцитов третьей стадии (около 20‰ у животных обоих полов). Число эритроцитов в 1 мкл крови также высоко, причем как у самок, так и у самцов малой лесной мыши (около 11 млн), при этом они небольших размеров (5.15 и 5.16 мкм соответственно). Концентрация гемоглобина в крови в среднем высокая, у особей обоих полов (табл. 2). По системе крови в зимний период по полу различия не отмечены.
Таблица 2.
Сезонная динамика параметров периферической крови малой лесной мыши (A. uralensis) в условиях среднегорий Центрального Кавказа
| Параметры | пол | M ± SE | p < 0.05 | |||
|---|---|---|---|---|---|---|
| Зима (1) | Весна (2) | Лето (3) | Осень (4) | |||
| HGB (г/л) | ♂ | 165.09 ± 3.0 | 164.90 ± 3.9 | 158.41 ± 1.8 | 157.21 ± 2.6 | |
| ♀ | 161.30 ± 3.1 | 158.82 ± 4.6 | 161.90 ± 2.0 | 154.71 ± 1.9 | ||
| RBC (млн в 1 мкл) | ♂ | 11.002 ± 0.3 | 9.380 ± 0.3 | 9.220 ± 0.1 | 9.930 ± 0.2 | 1–2, 1–3 |
| ♀ | 10.762 ± 0.4 | 9.898 ± 0.3 | 9.166 ± 0.2 | 10.203 ± 0.2 | 1–3, 3–4 | |
| HCT (об/%) | ♂ | 51.25 ± 1.4 | 51.65 ± 1.1 | 49.71 ± 0.6 | 49.50 ± 0.7 | |
| ♀ | 52.00 ± 1.1 | 50.83 ± 1.1 | 50.32 ± 0.8 | 49.31 ± 0.6 | ||
| Диаметр эритроцитов (мкм) | ♂ | 5.15 ± 0.05 | 5.50 ± 0.03 | 5.43 ± 0.04 | 5.39 ± 0.03 | 1–2, 1–3 |
| ♀ | 5.16 ± 0.04 | 5.41 ± 0.05 | 5.51 ± 0.04 | 5.35 ± 0.02 | 1–2, 1–3, 3–4 | |
| MCH (пг) | ♂ | 15.09 ± 0.4 | 17.87 ± | 17.22 ± 0.6 | 15.84 ± 0.2 | 1–2, 1–3, 3–4 |
| ♀ | 15.17 ± 0.6 | 15.88 ± | 17.82 ± 05 | 15.22 ± 0.3 | 1–3, 2–3, 3–4, | |
| MCV (мкм3) | ♂ | 47.83 ± 1.5 | 53.07 ± | 53.96 ± 1.8 | 50.18 ± 0.6 | 1–3, 3–4 |
| ♀ | 48.90 ± 1.9 | 51.62 ± | 55.20 ± 1.3 | 48.76 ± 0.7 | 1–3, 3–4, | |
| MCHC (г/л) | ♂ | 31.63 ± 0.05 | 31.49 ± | 31.90 ± 0.4 | 31.39 ± 0.2 | |
| ♀ | 31.06 ± 0.04 | 30.77 ± | 32.18 ± 0.5 | 31.25 ± 0.2 | 1–3 | |
Морфологический анализ эритроцитов на препаратах крови показал, что они нормохромные, и процесс их поступления не нарушен, о чем свидетельствует наличие 3–9 полихроматофильных эритроцитов в каждом поле зрения (рис. 1).
Рис. 1.
Микрофотография препарата крови A. ura-lensis: нормохромные (1) и полихроматофильные эритроциты (2).
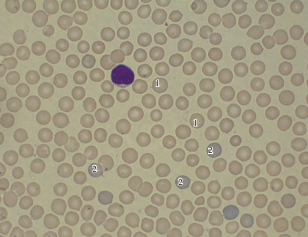
В эритроцитах обнаружены включения телец Хауэлла-Жолли (у 60% изученных животных). Все это свидетельствует о высокой активности эритропоэза в зимний период.
Система крови A. uralensis в весенний период (апрель). В весенний период эритроидные элементы на препаратах костного мозга встречались в каждом поле зрения в количестве до 30. В стадии митоза клетки красного ряда (2–3) отмечались не во всех полях зрения. Жировых клеток больше, чем зимой. Полученные результаты свидетельствуют о некотором снижении активности эритропоэза в весенний период, в сравнении с зимним (табл. 1). В парциальной эритрограмме (рис. 2) более низкое содержание эритробластов, статистически значимое в случае сравнения самок (p = 0.000).
Рис. 2.
Сезонная динамика содержания эритроидных клеток в костном мозге у малой лесной мыши (A. uralensis) в условиях среднегорий Центрального Кавказа.
1 – эритробласты, 2 – базофильные, 3 – полихроматофильные, 4 – оксифильные нормобласты.
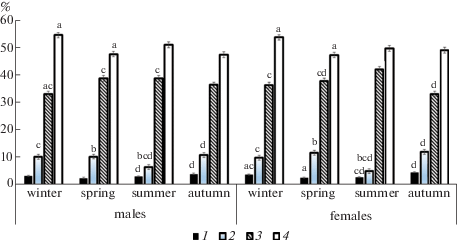
Средние значения содержания этих клеток весной составляют для самцов 2.59 ± 0.2% и 2.64 ± 0.50% для самок. Также ниже относительное количество оксифильных нормобластов – 47.93 ± 1.39% и 47.61 ± ± 3.25% (p = 0.000 и p = 0.009) соответственно.
Несмотря на это, содержание ретикулоцитов в периферической крови остается на уровне зимних значений, что свидетельствует об активном поступлении красных кровяных телец и в весенний период. Снижение показателей (рис. 3) отмечено лишь для третьей стадии (у самцов – 11.18 ± 0.8‰, у самок – 8.29 ± 0.4‰). Половые различия выражены в парциальном составе ретикулоцитов: у самок меньше зрелых клеток (III (p = 0.008) и IV (p = = 0.029) стадий) и больше молодых (II стадии (p = = 0.050), что связано с ускоренным выходом ретикулоцитов в периферическую кровь и способствует повышению дыхательной функции крови.
Рис. 3.
Сезонная динамика содержания ретикулоцитов в периферической крови Apodemus uralensis Pall.) в условиях Центрального Кавказа (пос. Эльбрус, 1800 м над ур. м.)
1 – I стадия, II – стадия, III – стадия, IV – стадия, 5 – общее количество ретикулоцитов.
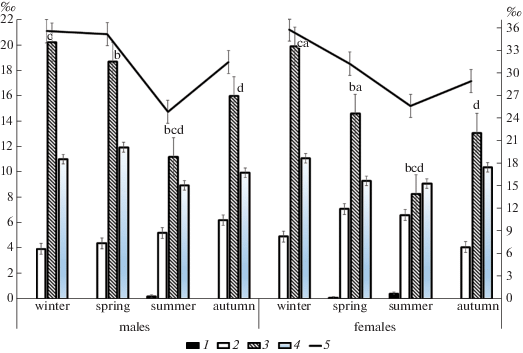
В периферической крови содержание эритроцитов весной снижается, по отношению к зиме. Размеры этих клеток весной увеличиваются (диаметр – у обоих полов, объем – MCV – достоверно у самцов). Содержание гемоглобина в 1 мкл крови и его средняя концентрация в эритроците остаются на уровне зимних показателей (табл. 2). Насыщенность эритроцитов гемоглобином (MCH) повышается у самцов, у самок остается на уровне зимних значений. Морфологический анализ препаратов крови показал, что поступающие клетки (эритроциты) из костного мозга в весенний период нормохромные. Полихроматофильные эритроциты, которые встречались в каждом поле зрения, свидетельствуют о нормальном процессе обновления эритроцитов. В эритроцитах, как и зимой, обнаружены тельца Хауэлла-Жолли, у 71.6% особей. Половые различия по параметрам периферической крови в данном сезоне выражены по среднему содержанию гемоглобина в эритроцитах (p = 0.001).
Система крови A. uralensis в летний период (июль). Эритроидные клетки на препаратах костного мозга летом встречались в количестве до 45 в каждом поле зрения. Отмечены 1–3 клетки в стадии митоза не во всех полях зрения. Летом активизируется эритропоэз костного мозга: значительно повышается общее количество эритроидных клеток у обоих полов. Их количество составило 22.59% у самцов и 23.47% – у самок. В эритрограмме меняется соотношение эритроидных клеток. Повышается количество эритробластов (3.21 ± 0.2% у самцов и 2.86 ± 0.2% у самок), значительно снижается (рис. 2) количество базофильных нормобластов (6.57 ± 0.5 и 5.09 ± 0.4% соответственно). Численные данные по полихроматофильным и оксифильным нормобластам – близки с весенними показателями. Летом в периферической крови по результатам дисперсионного анализа наблюдается снижение поступления ретикулоцитов, по сравнению с весной (рис. 3), за счет более зрелых клеток (в основном III стадии). При этом возрастает (рис. 3) доля молодых стадий (отмечены клетки первой стадии и несколько увеличивается количество второй). Число эритроцитов в периферической крови летом проявляет тенденцию к снижению. У самцов A. uralensis среднее содержание гемоглобина в эритроците сохраняется на уровне весенних значений, а у самок наблюдается его рост (до уровня самцов). Содержание гемоглобина в крови, гематокритный показатель остаются близкими с весенними данными (табл. 2). У самцов близки также летние и весенние показатели по средней концентрации гемоглобина и среднему объему эритроцитов. Морфологический анализ эритроцитов на препаратах крови показал, что они нормохромные, как и в предыдущие сезоны, процесс поступления молодых эритроцитов (полихроматофильных) увеличен. В эритроцитах 83.3% изученных животных обнаружены тельца Хауэлла-Жолли. Половые различия по изученным параметрам системы крови в данном сезоне выражены (по полихроматофильным нормобластам, ретикулоцитам III стадии).
Рис. 4.
Кластерный анализ по системе крови (20 параметров костного мозга, периферической крови) малой лесной мыши (Apodemus uralensis Pall.) в условиях Центрального Кавказа (пос. Эльбрус, 1800 м над ур. м.).
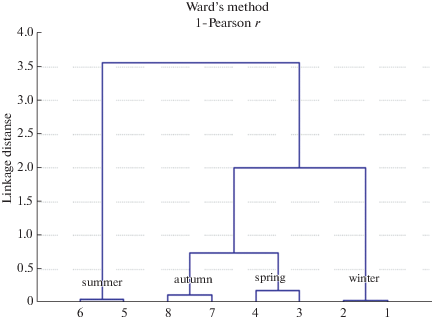
Система крови A. uralensis в осенний период (октябрь). Кроветворные клетки костного мозга осенью на препаратах размещались равномерно. Эритроидные клетки встречались в каждом поле зрения в количестве до 40, отмечены клетки в стадии митоза (1–2 не в каждом поле зрения). Дисперсионный анализ показал, что осенью количество клеток эритроидного ряда в костном мозге малой лесной мыши значительно снижается (у самцов и самок p = 0.01), по сравнению с летним периодом. При этом в парциальной эритрограмме у особей обоих полов повышается содержание эритробластов (у самцов – 4.07 ± 0.3%, у самок – 4.61% ± ± 0.2%) и базофильных нормобластов (11.10 ± 0.5 и 12.13 ± 0.7% соответственно). Доля более зрелых клеток – полихроматофильных (достоверно у самок) снижается. Процесс поступления молодых эритроцитов (ретикулоцитов) в кровь осенью у Apodemus uralensis более интенсивен по сравнению с летним периодом, однако различия у самок не достигают уровня достоверных различий (у самцов p = 0.01, у самок p = 0.264). Изменения наблюдаются в парциальном составе ретикулоцитов: увеличивается число клеток третьей стадии (у самцов 11.18 ± 0.8, p = 0.03, у самок 8.29 ± 0.4, p = 0.01).
Данные, полученные по ретикулоцитам, вполне согласуются с увеличением содержания эритроцитов в периферической крови. Осенью изменяются не только количественные, но и качественные характеристики этих клеток: они меньшего диаметра и объема и менее насыщены гемоглобином, по сравнению с летними (табл. 2). Морфологический анализ эритроцитов на препаратах крови показал, что они нормохромные и процесс поступления молодых эритроцитов (полихроматофильных) не нарушен. Тельца Хауэлла-Жолли в эритроцитах встречаются, но меньше, чем летом – 60%.
В крови осенью содержание гемоглобина, гематокритный показатель близки к летним значениям.
Как видно из рис. 4, кластерный анализ по расстоянию Махаланобиса (Ward’s method, 1 Pearson r) на основе показателей эритропоэза, ретикулоцитов периферической крови показал разделение выборок на 3 кластера. Выборки весны и осени наиболее сходны и образуют один кластер, который обладает большим сходством с зимними выборками, чем с летними.
Летние выборки отличаются от выборок других сезонов и образуют отдельный кластер. По изученным параметрам системы крови выявлена параллельная изменчивость самцов и самок малой лесной мыши.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ сезонной изменчивости эритропоэза свидетельствует о том, что у A. uralensis в среднегорьях эльбрусского варианта Центрального Кавказа кроветворная функция костного мозга сохраняется на высоком уровне в течение всего годового цикла, и подвержена сезонной изменчивости. Наиболее активное обновление крови наблюдается в зимний и летний периоды, и менее активное – в весенний и осенний. Процесс поступления молодых эритроцитов (ретикулоцитов) из костного мозга в периферическую кровь наиболее интенсивен зимой, сохраняется на высоком уровне в весенний период, а минимален – летом. Концентрация гемоглобина в крови A. uralensis в среднегорьях Центрального Кавказа стабильно поддерживается на высоком уровне в течение всего годового цикла. По всей вероятности, обитание в условиях хронической гипоксии обусловливает необходимость высокого оснащения организма A. uralensis кислородом независимо от сезона года. В способах поддержания необходимой кислородной емкости крови в разные сезоны года имеются существенные различия, что подтверждается результатами кластерного анализа.
Так, в летний период в периферическую кровь, при активном эритропоэзе, из костного мозга поступает относительно небольшое количество молодых эритроцитов (ретикулоцитов). Есть мнение [16], что ретикулоцит, который не прошел барьер, образованный стенкой сосудистого эндотелия, созревает в паренхиме, утрачивает способность диапедеза, остается блокированным и фагоцитируется макрофагами. Эти эритроциты входят в состав неэффективного эритропоэза.
Эритроциты A. uralensis в среднегорьях Центрального Кавказа в летний период имеют качественные особенности: они более крупные по своим размерам (диаметр, объем), с большим содержанием гемоглобина. При этом их количество летом падает до минимальных значений в годовом цикле. Высокая эритропоэтическая активность костного мозга при одновременном разжижении крови может способствовать увеличению кислородной емкости крови, при расчете на единицу массы тела [17].
Лесные мыши характеризуются относительным несовершенством физической терморегуляции и энергетические затраты их организма весьма значительны [18]. В зимний сезон, с низкими температурами внешней среды, который предполагает более высокий уровень метаболизма для поддержания постоянной температуры тела, эритропоэтическая активность костного мозга A. uralensis также высока, но с выходом большого количества ретикулоцитов (молодых эритроцитов) в периферическую кровь. Число эритроцитов в крови достигает максимальных значений в годовом цикле, причем размеры (диаметр и объем) этих клеток – минимальны. Все это обусловлено увеличением общей поглотительной поверхности для кислорода, а соответственно, более эффективным обеспечением организма кислородом. Известно [19], что меньшие по размеру эритроциты имеют более эффективный обмен кислорода. Увеличение доли “мелких” эритроцитов в период значительного снижения температуры среды [20, 21] объяснимо и с физико-химических позиций: чем более мелкодисперсная среда, тем больше площадь соприкосновения двух фаз, выше скорость диффузии. Большое число мелких эритроцитов в крови зимой обнаружено у ряда грызунов [11, 22–24]. Существует мнение, что уменьшение размеров эритроцитов зимой может быть генетически детерминированным [25]. Так, у высокогорных Phyllotis xanthopygus rupestris, выдерживающих сезонный перепад температур по меньшей мере 20°C, потомство рожденное и выросшее в условиях постоянной температуры, сохраняет сезонную изменчивость размеров эритроцитов [25]. Все выше сказанное позволяет предположить, что изменение размеров эритроцитов в первую очередь обусловлено изменением температуры среды.
В весенний период кроветворная функция костного мозга Apodemus uralensis снижается. Однако содержание ретикулоцитов, причем и более молодых стадий, в крови высокое, при довольно значительном снижении количества эритроцитов в периферической крови. Можно полагать, что это обусловлено увеличением эритродиереза, т.е. наблюдается усиленный распад красных клеток крови. В этот период года, по нашему мнению, и происходит замещение “мелких” зимних эритроцитов на крупные клетки.
В осенний сезон изученные показатели периферической крови, за исключением диаметра эритроцитов, ближе в основном к зимним значениям, при более низкой кроветворной активности костного мозга и соответствующих более низких показателях содержания ретикулоцитов в крови.
В эритроцитах малой лесной мыши (у более 70% особей летом, около 60% в другие сезоны) в условиях гор Центрального Кавказа отмечены тельца Хауэлла-Жолли, которые являются мелкими остатками ядерного материала. У желтогорлой мыши в Словакии [26] этих включений в эритроцитах зимой меньше, чем в другие сезоны. Присутствие их, возможно, объясняется повышенной скоростью продуцирования эритроцитов в момент регенеративного ответа [16, 17].
ЗАКЛЮЧЕНИЕ
Анализ комплекса гематологических показателей широко распространенного вида – A. uralensis в условиях среднегорий Центрального Кавказа позволил выявить сезонную динамику как в кроветворении, так и в показателях периферической крови.
Эритропоэтическая функция костного мозга вида в течение всего года поддерживается на высоком уровне, с выделением двух пиков: зимнего и летнего. При этом сезонные тренды поступления молодых эритроцитов (ретикулоцитов) отличны (максимальное – зимой, минимальное – летом). Динамика поступивших в кровь эритроцитов и содержания в них гемоглобина сходны: высокие показатели в холодный зимний сезон и низкие – в теплый летний. Изменчивость размеров эритроцитов – диаметра и объема – в противоположном направлении: максимальные значения – в весенне-летний период, минимальные – осенне-зимний. Наиболее изменчивыми в системе красной крови в сезонном аспекте являются эти показатели. В течение года стабильны – показатели содержания гемоглобина в крови и его концентрация в эритроцитах, гематокритный показатель. Относительно постоянные значения гематокритного числа A. uralensis обусловлены разнонаправленной сезонной динамикой размера и количества эритроцитов. Так, количество эритроцитов увеличивается в зимний период (что дает рост гематокритного числа), но одновременно средний диаметр эритроцита уменьшается. Соответственно, гематокритное число сохраняет относительно стабильные значения. Постоянство гематокрита позволяет избегать увеличения вязкости крови [27] при увеличении транспорта кислорода, что особенно важно в условиях действия хронической гипоксии.
Полученные нами данные свидетельствуют о высокой кислородной емкости крови A. uralensis в условиях гор Центрального Кавказа (1800 м над ур. м.) в течение всего годового цикла. При этом обеспечение высокого кислородного запроса вида в горах в разные сезоны года осуществляется разными способами. В осенне-зимний период – увеличением общей поглотительной поверхности для кислорода, обусловленное увеличением количества эритроцитов и уменьшением их размеров. В весенне-летний – поступлением в кровоток эритроцитов с качественно иными свойствами: более “крупных” с повышенным содержанием связывающего кислород гемоглобина (при относительно небольшом их количестве). Обнаруженные качественные изменения эритроцитов обусловливаются сезонными перестройками в эритропоэзе костного мозга.
У самцов и самок A. uralensis сезонная изменчивость по большинству изученных показателей синхронна. Половые различия по показателям системы красной крови у вида в условиях среднегорий Центрального Кавказа выражены слабо, вероятной причиной чего могут являться суровые климатические условия [28]. В отдельных случаях на сезонные изменения климатических факторов самцы реагируют быстрее самок, что согласуется с эволюционной теорией пола [29]. Так, объем эритроцитов и насыщение их гемоглобином достигают максимальных значений у самок летом, тогда как у самцов уже весной. В целом половые различия по показателям системы крови более выражены в весенне-летний период. Вероятнее всего это связано с процессами размножения и соответствующими изменениями гормонального фона.
Выявленная сезонная изменчивость в показателях костномозгового кроветворения и периферической крови малой лесной мыши обеспечивает поддержание, как оптимальной кислородной емкости крови, так и гомеостаза всего организма в горах Центрального Кавказа.
Список литературы
Pamenter ME, Hall JE, Tanabe Y, Simonson TS (2020) Cross-Species Insights Into Genomic Adaptations to Hypoxia. Frontiers in Genetics 11: 743. https://doi.org/10.3389/fgene.2020.00743
Storz JF (2007) Hemoglobin function and physiological adaptation to hypoxia in high-altitude mammals. J Mammal 88: 24–31. https://doi.org/10.1644/06-MAMM-S-199R1.1
Storz JF, Runck AM, Sabatino SJ, Kelly JK, Nuno F, Hideaki M, Weber RE, Angela F (2009) Evolutionary and functional insights into the mechanism underlying high-altitude adaptation of deer mouse hemoglobin. Proc Natl Acad Sci USA 106: 14450–14455. https://doi.org/10.1073/pnas.0905224106
Dzal YA, Sprenger RJ, Milsom WK (2020) Postnatal changes in O2 and CO2 sensitivity in rodents. Respiratory Physiology & Neurobiology. 103313. https://doi.org/10.1016/j.resp.2019.1033131
Dzal YA, Jenkin SEM, Lague SL, Reichert MN, York JM, Pamenter ME (2015) Oxygen in demand: How oxygen has shaped vertebrate physiology. Comparative Biochemistry and Physiology-Part A: Molecular and Integrative Physiology 186: 4–26. https://doi.org/10.1016/j.cbpa.2014.10.029
Pamenter ME, Powell FL (2016) Time domains of the hypoxic ventilatory response and their molecular basis. Comprehensive Physiology 6 (3): 1345–1385. https://doi.org/10.1002/cphy.c150026
Dzal YA, Milsom WK (2019) Hypoxia alters the thermogenic response to cold in adult homeothermic and heterothermic rodents. J Physiology. 597. 4809–4829. https://doi.org/10.1016/j.cbpa.2014.10.029
Темботов АК, Темботова ЭЖ, Берсекова ЗА, Емкужева ММ (2005) Влияние градиента высоты местности на гематологические показатели одного из широкораспространенных видов грызунов – малой лесной мыши (Muridae, Rodentia) на Центральном Кавказе. В кн.: Млекопитающие горных территорий М: КМК. 169–174. [Tembotov AK, Tembotova EZh, Bersekova ZA, Emkuzheva MM (2005) Vliyanie gradienta vysoty mestnosti na gematologicheskie pokazateli odnogo iz shirokorasprostranennyh vidov gryzunov – maloi lesnoi myshi (Muridae, Rodentia) na Central’nom Kavkaze [Effect of the altitudinal gradient on hematological indexes of the wide-spread rodent – small common field mouse (Murida, Rodentia) from the Caucasus ]. In: Mlekopitayushchie gornyh territorii. Moscow: KMK. 169–174. (In Russ)].
Cadena V, Tattersall GJ (2014) Body temperature regulation during acclimation to cold and hypoxia in rats. J Thermal Biol 46: 56–64. https://doi.org/10.1113/j.p277183
Соколов ВЕ, Темботов АК (1989) Млекопитающие Кавказа: Насекомоядные. М.: Наука. [Sokolov V.E., Tembotov A.K. (1989) Mlekopitayushchie Kavkaza: Nasekomoyadnye [Mammals of the Caucasus: Insectivores]. Moscow: Nauka (In Russ)].
Темботов АК, Темботова ЭЖ, Темботова ФА, Емкужева ММ (2009) Изменчивость гематологических параметров домовой мыши (Mus musculus L.) в предгорьях Центрального Кавказа. Усп. совр. биол. 129 (4): 370–378. [Tembotov AK, Tembotova EZh, Temboto-va FA, Emkuzheva MM (2009) Variation of hematological parameters in the house mouse (Mus musculus) in the Central Caucasus piedmont. Usp. sovr. biol. 129: 370–378. (In Russ)].
Amshokova AKh, Tembotova FA (2016) Differentiation of cryptic wood mice species of the subgenus Sylvaemus under different ecogeographic conditions in the Northern Caucasus. Russ J of Ecol 47: 376–382. https://doi.org/10.1134/S1067413616040044
Карасева ЕВ, Телицина АВ, Жигальский ОА (2008) Методы изучения грызунов в полевых условиях. М.: ЛКИ. [Karaseva E.V., Telitsyna A.Yu., Zhigal’ski O.A. (2008) Metody izucheniya gryzunov v polevykh usloviyakh [Methods of study of rodents in natural habitats] Moscow: LKI. (In Russ)].
Меньшиков ВВ (ред.) (2012) Стандартизация аналитических технологий лабораторной медицины. М.: Лабора. [Men’shikov V.V. (ed) (2012) Standartizaciya analiticheskih tekhnologij laboratornoj mediciny [Standardization of analytical technologies in laboratory medicine] Moscow: Labora. (In Russ)].
Риган В, Сандерс Т, Деникола Д (2000) Атлас ветеринарной гематологии. М.: Аквариум ЛТД. [Reagan W J, Sanders TG, DeNicola DB (1998) Veterinary hematology atlas of common domestic species. Iowa State University Press ].
Липунова ЕА, Скоркина МЮ (2004) Система красной крови: Сравнительная физиология: монография. Белгород: БелГУ. [Lipunova E.A., Skorkina M.Yu. (2004) Sistema krasnoj krovi: Sravnitel’naya fiziologiya: monografiya [Red blood system: Comparative physiology]. Belgorod: BelGU. (In Russ)].
Барбашова ЗИ (1981) Дыхательная функция крови. В кн.: Слоним А.Д. (ред) Экологическая физиология животных. Ч. II. Физиологические системы в процессе адаптации и факторы среды обитания: Руководство по физиологии. Л.: Наука [Barbashova ZI (1981) Dyhatel’naya funkciya krovi. In: Slonim A.D. (ed) Ekologicheskaya fiziologiya zhivotnyh. CH. II. Fiziologicheskie sistemy v processe adaptacii i faktory sredy obitaniya: Rukovodstvo po fiziologii. Leningrad: Nauka. (In Russ)].
Калабухов НИ (1969) Периодические (сезонные и годичные) изменения в организме грызунов, их причины и последствия. Л.: Наука, Ленингр. отд. [Kalabuhov NI (1969) Periodic changes in the body of rodents (seasons and consequences) Leningrad: Nauka (In Russ)].
Holland RA, Forster RE (1966) The effect of size of red cells on the kinetics of their oxygen uptake. J Gen Physiol 49 (4): 727–742. https://doi.org/10.1085/jgp.49.4.727
Тарахтий ЭА, Сумин МН, Давыдова ЮА (2009) Изменчивость показателей “красной” крови рыжей полевки (Clethrionomys glareolus) в зависимости от сезона и репродуктивного состояни. Усп соврем биол 129 (2): 191–197. [Tarakhtii E.A., Sumin M.N., Davydova Yu.A. (2009) Variability of red blood characteristics in bank vole (Clethriomys glareolus) related to season and reproductive status of its individuals. Usp sovrem biol 129 (2): 191–197. (In Russ)].
Тарахтий ЭА, Мухачева СB (2018) Химическое и радиационное загрязнение природной среды: эффекты в клетках системы крови мелких млекопитающих. Радиационная биология. Радиоэкология. 58 (3): 293–304. [Tarahtii EA, Muhacheva SV (2018) Chemical contamination and radiation environment: effects in the cells of the blood system of small mammals. Radiacionnaya biologiya. Radioekologiya. 58. (3): 293–304. (In Russ)]. https://doi.org/10.7868/S0869803118030098
Tarakhtii EA, Davydova YuA (2007) Seasonal variation in hematological indices in bank vole (Clethrionomys glareolus) in different reproductive states. Biol Bull Russ Acad Sci 34: 9–19. https://doi.org/10.1134/S1062359007010025
Rewkieweir-Dziarska A (1975) Seasonal changes in hemoglobin and erythrocyte indexes in Microtus arvalis (Pallas, 1779). Bull Acad Polon Sci. Ser Sci Biol 23: 481–486.
Perez-Suarei G, Arevalo F, Upez-Caballero, Lopez-Luna EP (1990) Seasonal variations in hematological values and heart weight in two small mammals, a mouse: Apodemus sylvaticus, and a vole: Pitymys duodecimcostatus. Acta teriologica 35: 33–40.
Ruiz G, Rosenmann M, Cortes A (2004) Thermal acclimation and seasonal variations of erythrocyte size in the Andean mouse Phyllotis xanthopygus rupestris. Comparative biochemistry and physiology a-molecular & integrative physiology. 139. 405–409. https://doi.org/10.1016/j.cbpb.2004.03.003
Husáriková M, Janiga M, Kufelová M (2015) Howell-Jolly bodies in peripheral blood of Yellownecked mouse (Apodemus flavicollis) living in the vicinity of pulp mill industrial complex. Oecologia Montana 24: 9–20.
Birchard GF (1997) Optimal hematocrit: theory, regulation and implications. Amer Zool 37: 65–72. https://doi.org/10.1093/icb/37.1.65
De Lisle SP, Goedert D, Reedy AM, Svensson EI (2018) Climatic factors and species range position predict sexually antagonistic selection across taxa. Phil Trans R Soc B 373: 20170415. https://doi.org/10.1098/rstb.2017.0415
Geodakian VA (2000) Evolutionary chromosomes and sex dimorphism. Biol Bull. 27: 99–113.
Дополнительные материалы отсутствуют.
Инструменты
Журнал эволюционной биохимии и физиологии