

Генетика, 2019, T. 55, № 10, стр. 1231-1236
Особенности распределения регионов высокой гомозиготности в геномах представителей коренного населения Северной Евразии
Н. А. Колесников 1, *, В. Н. Харьков 1, А. А. Зарубин 1, В. А. Степанов 1
1 Научно-исследовательский институт медицинской генетики, Томский национальный исследовательский медицинский центр Российской академии наук
634050 Томск, Россия
* E-mail: nikita.kolesnikov@medgenetics.ru
Поступила в редакцию 22.01.2019
После доработки 19.03.2019
Принята к публикации 15.04.2019
Аннотация
Популяции коренных этносов Северной Евразии представляют значительный интерес для популяционной геномики как в силу относительно слабой их изученности с привлечением современных геномных технологий, так и по причине специфичности их генофондов, развивавшихся в различных генетико-демографических условиях. Мы использовали данные генотипов по 242 179 аутосомным SNP у 876 человек для поиска регионов высокой гомозиготности протяженностью более 1 Mb. Для популяций Сибири обнаружено большее общее количество и большая суммарная длина регионов высокой гомозиготности, чем для других исследованных популяций Северной Евразии.
Генетико-демографические процессы и естественный отбор в популяциях человека оказывают влияние на структурированность генетического разнообразия в геномах отдельных индивидов и популяциях в целом. Они, в частности, приводят к тому, что формируются особенности распределения регионов высокой гомозиготности (англ. ROH – runs of homozygosity) между отдельными людьми и популяциями, что обеспечивает ценный и часто неиспользуемый ресурс для изучения генетического разнообразия человечества и его эволюционной истории [1, 2].
Число и общая длина ROH у каждого человека показывают значительные различия между отдельными людьми и группами населения. Сумма длин коротких и средних ROH у индивида возрастает с увеличением расстояния популяций от Восточной Африки, коррелируя с процессами расселения человека [1]. Напротив, общий суммарный размер длинных ROH показывает большие межиндивидуальные вариации, которые, вероятно, отражают события недавнего инбридинга, причем более высокие значения чаще встречаются в популяциях с известными высокими частотами кровнородственных браков. Регионы высокой гомозиготности также связаны с эффективным размером популяции, небольшие популяции имеют тенденцию к накоплению большего числа ROH по сравнению с более многочисленными группами населения. Инбридинг добавляет небольшое количество очень длинных ROH, тогда как эффект бутылочного горлышка увеличивает количество ROH, которые, как правило, остаются относительно короткими. Наконец, наибольшая длина и количество ROH наблюдаются в популяциях с относительно небольшим эффективным размером в прошлом и недавним инбридингом [3, 4].
Территория Северной Евразии обладает уникальными географическими характеристиками с высоким уровнем этнической и лингвистической дифференциации населения, а также в этом регионе исторически имело место переплетение коренных и пришлых генетических элементов, проникавших сюда с соседних территорий. Популяции коренного населения Сибири характеризуются, как правило, небольшой численностью и значительной географической изоляцией от других популяций, что находит отражение в структуре их генетического разнообразия [5–9]. Задачей настоящей работы была оценка распределения регионов высокой гомозиготности в популяциях различных территорий Северной Евразии.
Мы использовали генотипы 876 индивидуумов по 242 179 аутосомным SNP, опубликованные в работе Triska et al. [5]. Популяции коренного населения Сибири представлены кетами (N = 26), тувинцами (N = 44), хантами (N = 29), бурятами (N = 44), якутами (N = 41). Волго-Уральский географический регион представлен популяциями коми (N = 32), чувашей (N = 28), башкир (N = 34), мордвы (N = 33), татар (N = 33). Популяции Европы представлены карелами (N = 26), белорусами (N = 28), молдаванами (N = 32), русскими (N = 77), украинцами (N = 29). Популяции карачаевцев (N = 22), чеченцев (N = 18), адыгейцев (N = 31), мегрелов (N = 34), абхазов (N = 26), кабардинцев (N = 35), черкесов (N = 32) относятся к Кавказскому региону, а популяции киргизов (N = 28), узбеков (N = 35) и казахов (N = 44) – к Средней Азии. Метисы первого поколения были исключены из анализа.
ROH были идентифицированы с помощью программы Runs of Homozygosity, реализованной в PLINK версии 1.9 [10] с использованием стандартных настроек. Для сравнения данных полученные результаты были обобщены путем расчета средних значений количества ROH и совокупной длины ROH на человека для каждой популяции. Статистическую значимость межпопуляционных различий оценивали при помощи теста Уилкоксона (уровень значимости = 0.05).
Медианное число ROH на индивидуальный геном в исследованных образцах составило 7, граница первого квартиля 4, граница третьего квартиля 12. Медианная длина регионов 15 Mb, граница первого квартиля 7.5 Mb, третьего – 35.2 Mb. Для популяций Сибири медианное число ROH составило 19, с границей для первого квартиля 13 и третьего 27, медианная длина регионов 55.7 Mb с границами 38 и 86.1 Mb для первого и третьего квартиля соответственно.
Средняя суммарная длина ROH во всех диапазонах в популяциях коренного населения Сибири значительно превышает все остальные географические регионы (табл. 1). Средняя суммарная длина ROH в популяциях Волго-Уральского региона выше в диапазонах длин 1–2, 2–3, 3–4, 4–5, 5–6, 6–8 Mb, а также по сумме всех диапазонов по сравнению с популяциями Европы, Средней Азии и Кавказа (табл. 1). Для популяций Средней Азии в диапазоне более 10 Mb средняя суммарная длина ROH выше, чем у популяций кавказского и европейского регионов, за счет популяций киргизов, близкородственных по происхождению к южносибирским этносами узбеков.
Таблица 1.
Средняя суммарная длина ROH (Mb), а также доля человек с одним или несколькими ROH в исследуемых популяциях по диапазонам длин (Mb) для каждого региона
| Диапазон длин | Сибирь | Кавказ | Средняя Азия | Европа | Волго-Уральский регион |
|---|---|---|---|---|---|
| 1–2 | 15.14 | 4.40 | 3.69 | 6.65 | 7.45 |
| 100% | 100% | 100% | 100% | 100% | |
| 2–3 | 11.40 | 3.24 | 2.34 | 3.46 | 5.09 |
| 100% | 100% | 100% | 100% | 100% | |
| 3–4 | 6.99 | 1.41 | 1.25 | 1.58 | 2.80 |
| 74% | 48% | 35% | 33% | 30% | |
| 4–5 | 5.61 | 1.03 | 0.64 | 1.09 | 2.23 |
| 66% | 37% | 17% | 16% | 15% | |
| 5–6 | 4.75 | 0.88 | 0.51 | 0.51 | 1.65 |
| 51% | 18% | 8% | 20% | 8% | |
| 6–8 | 5.78 | 1.07 | 0.62 | 1.04 | 1.71 |
| 42% | 23% | 11% | 18% | 13% | |
| 8–10 | 3.93 | 1.16 | 0.55 | 0.59 | 1.12 |
| 33% | 11% | 6% | 10% | 6% | |
| >10 | 14.27 | 1.58 | 4.09 | 2.59 | 4.01 |
| 43% | 22% | 6% | 10% | 12% | |
| Σ | 67.87 | 14.76 | 13.70 | 17.50 | 26.05 |
Популяции коренного населения Сибири по среднему количеству и средней суммарной длине ROH образуют две группы: в первую группу входят якуты, кеты и ханты, во вторую – тувинцы и буряты (рис. 1). Для кетов наблюдается наибольшее количество и длина ROH, что, вероятно, связанно с сильным инбридингом, малой эффективной численностью и географической изоляцией популяции. В популяциях кетов, хантов и бурятов наблюдается существенный вклад более протяженных регионов высокой гомозиготности, что также свидетельствует о недавнем сильном инбридинге (табл. 2). Ранее по маркерам Y-хромосомы у бурятов было обнаружено относительно низкое генетическое разнообразие по сравнению с другими популяциями Южной Сибири и сильные эффекты основателя по двум основным гаплогруппам [11]. Для якутов, несмотря на большой эффективный размер популяции, также наблюдается большое количество и длина ROH, что, вероятно, связано с сильным эффектом основателя, который выявляется на маркерах мтДНК и Y-хромосомы [12].
Рис. 1.
Количество ROH в исследуемых популяциях (а). Суммарные длины ROH (Mb) в исследуемых популяциях (б).
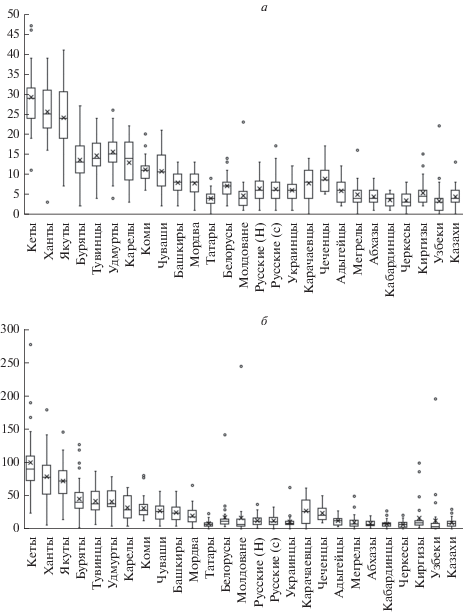
Таблица 2.
Среднee суммарное количество и длина ROH по диапазонам (Mb) в исследуемых популяциях
| Популяции | Среднее суммарное количество по диапазонам (Mb) | Средняя суммарная длина по диапазонам (Mb) | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1–2 | 2–3 | 3–4 | 4–5 | 5–6 | 6–8 | 8–10 | >10 | Σ | 1–2 | 2–3 | 3–4 | 4–5 | 5–6 | 6–8 | 8–10 | >10 | Σ | |
| Карелы | 7.38 | 2.85 | 1.04 | 0.62 | 0.27 | 0.42 | 0.15 | 0.19 | 12.90 | 10.68 | 7.07 | 3.52 | 2.72 | 1.47 | 2.91 | 1.38 | 2.90 | 32.66 |
| Абхазы | 2.58 | 1.15 | 0.31 | 0.15 | 0.04 | 0.08 | 0.00 | 0.00 | 4.30 | 3.82 | 2.73 | 1.04 | 0.72 | 0.20 | 0.57 | 0.00 | 0.00 | 9.08 |
| Адыгейцы | 3.61 | 1.26 | 0.42 | 0.16 | 0.29 | 0.06 | 0.00 | 0.03 | 5.80 | 5.37 | 3.06 | 1.39 | 0.73 | 1.62 | 0.44 | 0.00 | 0.37 | 12.97 |
| Башкиры | 3.65 | 1.79 | 0.79 | 0.50 | 0.41 | 0.26 | 0.18 | 0.32 | 7.90 | 5.22 | 4.35 | 2.77 | 2.28 | 2.24 | 1.88 | 1.59 | 5.28 | 25.62 |
| Белорусы | 4.75 | 1.25 | 0.43 | 0.25 | 0.00 | 0.11 | 0.07 | 0.14 | 7.00 | 7.03 | 2.98 | 1.48 | 1.08 | 0.00 | 0.71 | 0.59 | 3.83 | 17.70 |
| Буряты | 7.05 | 2.48 | 1.14 | 0.82 | 0.50 | 0.48 | 0.32 | 0.75 | 13.50 | 10.15 | 6.03 | 3.98 | 3.70 | 2.71 | 3.26 | 2.76 | 13.28 | 45.86 |
| Кабардинцы | 2.14 | 0.91 | 0.14 | 0.03 | 0.09 | 0.03 | 0.09 | 0.09 | 3.50 | 3.01 | 2.15 | 0.49 | 0.12 | 0.49 | 0.20 | 0.74 | 1.12 | 8.31 |
| Казахи | 2.62 | 1.09 | 0.31 | 0.13 | 0.09 | 0.04 | 0.02 | 0.04 | 4.40 | 3.85 | 2.61 | 1.06 | 0.57 | 0.50 | 0.32 | 0.22 | 0.51 | 9.86 |
| Карачаевцы | 3.26 | 1.70 | 0.65 | 0.39 | 0.39 | 0.43 | 0.57 | 0.39 | 7.80 | 4.75 | 4.12 | 2.26 | 1.73 | 2.12 | 2.96 | 5.14 | 4.94 | 29.29 |
| Кеты | 14.04 | 6.50 | 2.54 | 1.73 | 1.23 | 1.15 | 0.77 | 1.31 | 29.30 | 20.69 | 15.66 | 8.63 | 7.70 | 6.67 | 7.87 | 6.88 | 25.90 | 100 |
| Киргизы | 3.00 | 1.21 | 0.36 | 0.21 | 0.07 | 0.11 | 0.07 | 0.21 | 5.30 | 4.35 | 2.92 | 1.18 | 0.93 | 0.39 | 0.75 | 0.64 | 5.64 | 16.80 |
| Коми | 6.28 | 2.34 | 0.72 | 0.66 | 0.19 | 0.31 | 0.19 | 0.44 | 11.10 | 9.07 | 5.71 | 2.47 | 2.85 | 1.02 | 2.12 | 1.61 | 6.52 | 31.37 |
| Мегрелы | 2.92 | 1.28 | 0.36 | 0.11 | 0.11 | 0.03 | 0.06 | 0.03 | 4.90 | 4.40 | 3.20 | 1.29 | 0.52 | 0.62 | 0.21 | 0.53 | 0.88 | 11.64 |
| Молдаване | 2.59 | 0.72 | 0.38 | 0.13 | 0.19 | 0.19 | 0.09 | 0.34 | 4.60 | 3.68 | 1.76 | 1.25 | 0.60 | 1.06 | 1.38 | 0.84 | 6.30 | 16.87 |
| Мордва | 4.61 | 1.64 | 0.52 | 0.45 | 0.15 | 0.12 | 0.06 | 0.24 | 7.80 | 6.67 | 4.04 | 1.75 | 2.03 | 0.83 | 0.83 | 0.51 | 4.06 | 20.71 |
| Русские (Н) | 3.89 | 1.58 | 0.39 | 0.21 | 0.08 | 0.11 | 0.03 | 0.03 | 6.30 | 5.68 | 3.84 | 1.33 | 0.91 | 0.41 | 0.71 | 0.24 | 0.29 | 13.39 |
| Русские (с) | 4.36 | 1.15 | 0.28 | 0.21 | 0.03 | 0.08 | 0.05 | 0.05 | 6.20 | 6.31 | 2.80 | 0.95 | 0.93 | 0.14 | 0.54 | 0.48 | 0.67 | 12.81 |
| Татары | 2.50 | 0.94 | 0.12 | 0.06 | 0.06 | 0.18 | 0.03 | 0.03 | 3.90 | 3.71 | 2.39 | 0.41 | 0.28 | 0.34 | 1.33 | 0.26 | 0.32 | 9.04 |
| Тувинцы | 7.64 | 3.30 | 1.34 | 0.80 | 0.30 | 0.66 | 0.20 | 0.41 | 14.60 | 11.20 | 7.90 | 4.70 | 3.60 | 1.60 | 4.51 | 1.76 | 7.11 | 42.38 |
| Удмурты | 7.96 | 3.52 | 1.68 | 0.76 | 0.64 | 0.44 | 0.28 | 0.24 | 15.50 | 11.24 | 8.55 | 6.02 | 3.38 | 3.45 | 2.94 | 2.42 | 3.80 | 41.79 |
| Узбеки | 1.64 | 0.51 | 0.38 | 0.08 | 0.10 | 0.10 | 0.08 | 0.31 | 3.20 | 2.79 | 1.43 | 1.47 | 0.39 | 0.64 | 0.79 | 0.80 | 6.10 | 14.43 |
| Украинцы | 4.55 | 1.00 | 0.28 | 0.07 | 0.00 | 0.00 | 0.00 | 0.07 | 6.00 | 6.51 | 2.30 | 0.94 | 0.30 | 0.00 | 0.00 | 0.00 | 1.54 | 11.59 |
| Ханты | 12.79 | 5.41 | 2.52 | 1.28 | 1.07 | 1.00 | 0.48 | 0.97 | 25.50 | 18.33 | 13.00 | 8.71 | 5.75 | 5.84 | 6.84 | 4.41 | 16.27 | 79.14 |
| Черкесы | 2.03 | 0.91 | 0.18 | 0.03 | 0.18 | 0.09 | 0.00 | 0.03 | 3.50 | 2.94 | 2.28 | 0.60 | 0.13 | 1.01 | 0.67 | 0.00 | 0.32 | 7.96 |
| Чеченцы | 4.39 | 2.11 | 0.78 | 0.72 | 0.00 | 0.33 | 0.17 | 0.28 | 8.80 | 6.29 | 4.97 | 2.66 | 3.21 | 0.00 | 2.29 | 1.47 | 3.19 | 24.08 |
| Чуваши | 6.04 | 2.32 | 0.96 | 0.57 | 0.36 | 0.18 | 0.04 | 0.29 | 10.80 | 8.81 | 5.47 | 3.37 | 2.56 | 2.00 | 1.17 | 0.31 | 4.10 | 27.79 |
| Якуты | 10.66 | 5.93 | 2.56 | 1.61 | 1.27 | 0.95 | 0.44 | 0.63 | 24.0 | 15.36 | 14.43 | 8.91 | 7.28 | 6.95 | 6.43 | 3.82 | 8.79 | 71.97 |
Среди европейских популяций значительно отличаются карелы, показывая большее количество и длину, а также максимальный разброс ROH (рис. 1), что, вероятно, свидетельствует о наличии у них эффекта основателя. Карелы этнически и генетически близки к финнам, у которых также обнаружен очень сильный эффект основателя по маркерам мтДНК и Y-хромосомы [13]. По сравнению с другими исследованными европейскими этносами у карелов меньше эффективная численность популяции, следовательно, ожидается высокая степень гомозиготизации генома по сравнению с более гомогенными и обладающими высокой эффективной численностью популяциями, такими как восточные славяне, украинцы, белорусы и молдаване.
По доле образцов с одним и более ROH длиной больше 10 Mb по разным регионам выделяются три различные группы, которые явно связаны с эндогамией и изоляцией: большая часть сибирских образцов имеет длинные ROH (43% имеют ROH > 10 Mb), далее идут образцы Кавказского региона (22% имеют ROH > 10 Mb) и для среднеазиатского, европейского и Волго-Уральского регионов составляют соответственно 6, 10 и 12% (табл. 1).
Таким образом, в популяциях коренного населения Сибири количество и суммарная длина ROH значительно выше, чем в остальных исследуемых популяциях Северной Евразии, за исключением карелов и удмуртов, не показывающих значимых отличий с бурятами и тувинцами. Вероятно, это связанно с генетико-демографической историей популяций, генетическое разнообразие которых складывалось под действием эффекта основателя, инбридинга, миграций, изоляцией расстоянием.
Работа выполнена в рамках Научно-технической программы союзного государства “Разработка инновационных геногеографических и геномных технологий идентификации личности и индивидуальных особенностей человека на основе изучения генофондов регионов Союзного государства (ДНК-идентификания)”, госконтракт № 011-17 от 26.09.2017 г., а также поддержана грантом РФФИ № 18-29-13045 “Популяционная геномика и транскриптомика человека: поиск сигналов не-нейтральной эволюции”.
Все процедуры, выполненные в исследовании с участием людей, соответствуют этическим стандартам институционального и/или национального комитета по исследовательской этике и Хельсинкской декларации 1964 г. и ее последующим изменениям или сопоставимым нормам этики.
От каждого из включенных в исследование участников было получено информированное добровольное согласие.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Pemberton T.J., Absher D., Feldman M.W. et al. Genomic patterns of homozygosity in worldwide human populations // Am. J. Hum. Genet. 2012. V. 8. № 91. P. 275–292. https://doi.org/10.1016/j.ajhg.2012.06.014
Ropers H.H. New perspectives for the elucidation of genetic disorders // Am. J. Hum. Genet. 2007. V. 81. P. 199–207. https://doi.org/10.1086/520679
Ceballos F.C., Joshi P.K., Clark D.W. et al. Runs of homozygosity: windows into population history and trait architecture // Nat. Rev. Genet. 2018. V. 19(4). P. 220–234. https://doi.org/10.1038/nrg.2017.109
Triska P., Chekanov N., Stepanov V. et al. Between Lake Baikal and the Baltic Sea: Genomic history of the gateway to Europe // BMC Genetics. 2017. V. 18. (Suppl. 1). P. 110. https://doi.org/10.1186/s12863-017-0578-3
Степанов В.А., Пузырев В.П. Микросателлитные гаплогрупы Y-хромосомы демонстрируют отсутствие подразделенности и наличие нескольких компонентов в мужском генофонде тувинцев // Генетика. 2000. Т. 36. № 3. С. 377–384.
Степанов В.А., Пузырев В.П. Анализ аллельных частот семи микросателлитных локусов Y-хромосомы в трех популяциях тувинцев // Генетика. 2000. Т. 36. № 2. С. 241–248.
Степанов В.А., Харьков В.Н., Пузырев В.П. Эволюция и филогеография линий Y-хромосомы человека // Информацион. вестн. ВОГиС. 2006. Т. 10. № 1. С. 57–73.
Хитринская И.Ю., Степанов В.А., Пузырев В.П. и др. Генетическое своеобразие население Якутии по данным аутосомных локусов // Мол. биол. 2003. Т. 37. № 2. С. 234–239.
Степанов В.А. Этногеномика населения Северной Евразии. Томск: Печатная мануфактура, 2002. 244 с.
Purcel Sh., Chang Ch. PLINK [1.9]. URL: www.cog-genomics.org/plink/1.9/.
Харьков В.Н., Хамина К.В., Медведева О.Ф. и др. Генофонд бурят: клинальная изменчивость и территориальная подразделенность по маркерам Y-хромосомы // Генетика. 2014. Т. 50. № 2. С. 203–213. https://doi.org/10.1134/S1022795413110082
Харьков В.Н., Степанов В.А., Медведева О.Ф. и др. Происхождение якутов: анализ гаплотипов Y-хромосомы // Мол. биол. 2008. Т. 42. № 2. С. 226–237.
Sajantila A., Salem A.H., Savolainen P. et al. Paternal and maternal DNA lineages reveal a bottleneck in the founding of the Finnish population // PNAS USA. 1996. V. 93. № 21. P. 12035–12039.
Дополнительные материалы отсутствуют.