

Генетика, 2019, T. 55, № 11, стр. 1298-1311
Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у дикого (Hordeum spontaneum C. Koch) и культурного (Hordeum vulgare L.) ячменя в Израиле и Палестине
А. А. Поморцев 1, *, А. В. Рубанович 1, О. Н. Ковалева 2, Е. В. Лялина 1
1 Институт общей генетики им. Н.И. Вавилова Российской академии наук
119991 Москва, Россия
2 Федеральный исследовательский центр Всероссийский институт генетических ресурсов растений
им. Н.И. Вавилова
190000 Санкт-Петербург, Россия
* E-mail: a.pomortsev@gmail.com
Поступила в редакцию 28.03.2019
После доработки 07.05.2019
Принята к публикации 15.05.2019
Аннотация
Методом электрофореза в крахмальном геле изучен полиморфизм гордеинов, контролируемых локусами Hrd A и Hrd B, в 13 местных образцах культурного и в 31 популяции дикого ячменя из Израиля и Палестины. Для локуса Hrd A в H. spontaneum и H. vulgare идентифицированы 114 и 11 аллелей, для локуса Hrd B – 114 и 22 аллелей соответственно. Частоты аллелей локусов Hrd A и Hrd B в образцах дикого ячменя варьировали в пределах 0.0019–0.0910 и 0.0019–0.0743 соответственно, а культурного ячменя – в пределах 0.0143–0.4215 и 0.0071–0.200 соответственно. В израильских популяциях H. spontaneum и H. vulgare из 120 аллелей локуса Hrd A обнаружено шесть общих аллелей, а из 135 аллелей локуса Hrd В – только один. При этом в H. spontaneum присутствовали 18 аллелей локуса Hrd A и 15 аллелей локуса Hrd В, найденные нами ранее также в местных образцах культурного ячменя из различных стран, входящих и сопредельных с центром разнообразия ячменя. “Культурные” аллели локусов Hrd A и Hrd B обнаружены в основном в популяциях дикого ячменя из района озера Кинерет. В популяции H. spontaneum из Суситы присутствовали генотипы Hrd A3 B162 и Hrd A24 B65, обнаруженные и в местном культурном ячмене из Сирии и Иордании. В целом, анализ аллельного разнообразии гордеин-кодирующих локусов в популяциях израильского дикого ячменя, состава и распространения обнаруженных в H. spontaneum “культурных” аллелей и отдельных генотипов в местных образцах культурного ячменя из 20 стран показал, что некоторые популяции H. spontaneum из Израиля могли быть донорами отдельных аллелей и генотипов гордеин-кодирующих локусов для H. vulgare в результате интрогрессии при спонтанной гибридизации в ходе распространения культурного ячменя.
Ячмень является одним из первых одомашненных человеком растений. Предполагается, что ячмень был введен в культуру на Ближнем Востоке в VIII–VII тыс. до н.э. [1, 2]. Однако о месте доместикации ячменя другими авторами в различное время выдвигались иные гипотезы, согласно которым культурный ячмень впервые появился в тех или иных районах от Северной Африки до Тибета: Южной Туркмении [3], Юго-Западноазиатском центре разнообразия растений [4], Тибете [5, 6], Египте [3], Марокко [7], Эфиопии [8], Юго-восточной Турции [9], в долине р. Иордан [10], Западном Пакистане [11]. Одни гипотезы основываются на имевшихся в определенный момент времени данных археологических раскопок, другие – на изучении существующего морфологического разнообразия, исследовании полиморфных белков и ДНК-маркеров. Кроме этого, предложена гипотеза, что культурный ячмень – результат множественной спонтанной интрогрессии генетического материала от местных популяций H. spontaneum в культурный ячмень при его распространении человеком в ареале дикого ячменя [12]. Авторы этой гипотезы считают, что такой взгляд на вопрос о происхождении культурного ячменя предпочтительнее, чем дискретные центры происхождения. Проведенные нами ранее исследования аллельного разнообразия высокополиморфных локусов Hrd A и Hrd B, контролирующих синтез гордеина в зерне, в образцах местного культурного и дикого ячменя из Западного Ирана [13], Юго-Восточной Турции [14], Сирии [15] и Иордании [16] позволили сделать вывод, что территории каждой из этих стан вряд ли могут рассматриваться как центры доместикации ячменя. Однако нельзя исключить интрогрессию отдельных аллелей гордеин-кодирующих локусов в H. vulgare от H. spontaneum в результате спонтанной гибридизации при распространении культурного ячменя.
Исследование аллельного разнообразия высокополиморфных локусов Hrd A и Hrd B в образцах местного культурного и дикого ячменя из Израиля и Палестины в свете изучения вопроса о центре происхождения представляет особый интерес. С одной стороны, эта территория входит в состав Дуги Плодородия, с другой стороны, на Ближнем Востоке самое раннее использование человеком зерна ячменя, датируемое XVII тыс. до н.э., зафиксировано в Охало II около Галилейского моря [17]. Кроме этого, археоботанические исследования свидетельствуют, что климатические условия на территории современного Израиля и Палестины существенно не менялись за последние 10–12 тыс. лет [18]. Это позволяет надеяться на отсутствие значительного влияния внешних условий на генетическую структуру популяций как дикого, так и культурного ячменя.
МАТЕРИАЛЫ И МЕТОДЫ
Аллельное разнообразие локусов Hrd A и Hrd B изучали в 31 популяции H. spontaneum и 13 образцах местного культурного ячменя. Популяции дикого ячменя представляли собой оригинальные сборы в районах Голанских высот, Галилейского моря (оз. Кинерет), Хайфы и границы с Палестиной (рис. 1, табл. 1). Материал был собран О. Раскиной, А. Беляевым и Е. Бадаевой в 2000–2008 гг. Всего из популяций H. spontaneum гордеины были исследованы в зернах от 503 растений. Образцы H. vulgare, собранные в 1922–1926 гг., были получены из коллекции генетических ресурсов ВИР им. Н.И. Вавилова. От каждого образца культурного ячменя гордеины анализировали в 10 отдельных зернах. Варианты блоков компонентов, контролируемые аллелями локусов Hrd A и HrdB, и генетические формулы гордеина по этим локусам определяли по каталогам, составленным нами ранее на основе изучения более 1600 образцов культурного ячменя из стран, входящих в центры разнообразия и граничащих с ними [19]. Аллели этих локусов и соответствующие им варианты блоков компонентов гордеинов А и В, которые были выявлены исключительно в H. spontaneum, обозначали цифрами с индексом “s”.
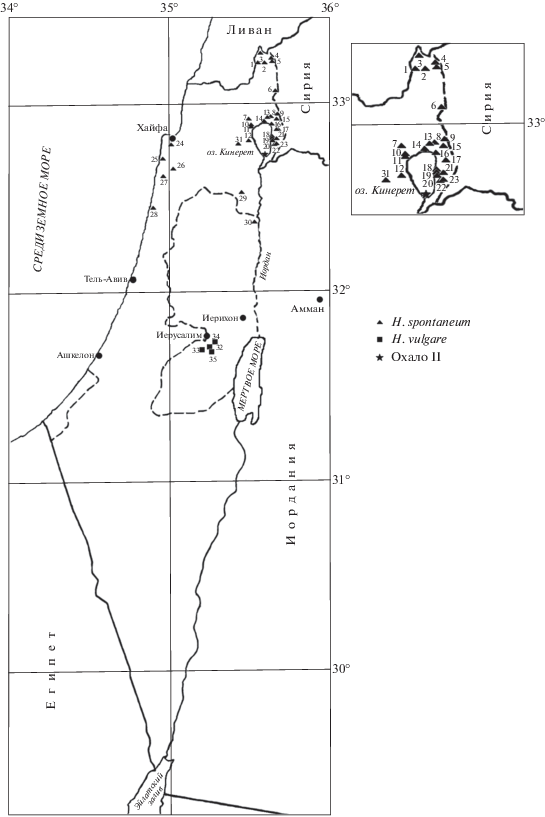
Таблица 1.
Генетические формулы гордеина и места сбора образцов дикого (H. spontaneum) и культурного (H. vulgare) ячменя в Израиле и Палестине
| № п/п | № каталога ВИР | Формула гордеина по локусам | Место сбора образцов, координаты | |
|---|---|---|---|---|
| Hrd A | Hrd B | |||
| H. spontaneum | ||||
| 1 | Оригинальные сборы | 162s + 175s | 211s + 213s | Мицпе Пеэр (1)* |
| 2 | То же | 131s + 155s + 166s + 167s | 203s + 204s | Нахаль Орвим (2) |
| 3 | » | 55s + 180s + 181s + 182s + + 183s + 184s + 185s | 180s + 189s + 192s + 215s + + 220s + 221s + 222s + 224s | Хермон (3) |
| 4 | » | 12 + 55s + 150s + 151s | 189s + 190s + 191s + 192s | 33°16′ с.ш., 35°45′ в.д. (4), Мигдал Шамс |
| 5 | » | 105s + 152 | 15 + 97s | 33°14′ с.ш., 35°41′ в.д., Баниас (5) |
| 6 | » | 152 + 91s + 158s + 159s + 160s | 41 + 161s + 199s + 200s + 247s | 33°06′ с.ш., 35°48′ в.д. (6), Эль Кунейтра |
| 7 | » | 106s + 116s + 134s | 182s + 183s + 187s | Рош Пина (7) |
| 8 | » | 175s + 176s + 177s | 162 + 213s | Кешет (8) |
| 9 | » | 151s + 169s + 172s + + 173s + 174s | 161s + 210s + 213s | 32°55′ с.ш., 35°45′ в.д., дорога на Джанатан (9) |
| 10 | » | 24 + 138s + 139s + 142s | 174s + 177s + 178s | Табигха, известняк (10) |
| 11 | » | 1 + 96s + 140s + 141s | 141 + 189 + 158s + 179s | Табигха, базальт (11) |
| 12 | » | 1 + 2 + 12 + 118s + 119s + + 120s + 121s + 122s + 123s + + 124s + 125s + 147s | 123s + 152s + 153s + 154s + + 155s + 156s + 157s + + 158s + 159s | 32°49′ с.ш., 35°29′ в.д. (12), Арбель |
| 13 | » | 33 + 30s | 208s + 209s + 210s | 32°59′ с.ш., 35°41′ в.д. (13), Казрин |
| 14 | » | 63 + 110 | 192s + 212s | Долина р. Иордан, мост Эрик (14) |
| 15 | » | 10s + 39s + 107s + 113s + 114s + + 115s + 116s + 161s | 87s + 140s + 141s + 142s + + 143s + 144s + 145s + 146s + + 147s + 148s + 149s + 150s + + 151s | 32°55′ с.ш., 35°41′ в.д., Йехудия (15) |
| 16 | » | 1 + 3 + 18 + 61 + 41s + 52s + + 116s + 126s | 160s + 161s + 162s + 163s + + 164s + 260s | Галма (16) |
| 17 | » | 116s + 170s + 171s | 162 + 206s + 207s | Авней Эйтан (17) |
| 18 | » | 1 + 3 + 12 + 13 + 18 + 23 + + 24 + 28 + 33 + 116s + + 127s + 128s + 129s + 130s + + 131s + 132s | 65 + 162 + 169 + 15s + 33s + + 146s + 163s + 165s + 166s + + 167s + 168s + 169s | 32°46′ с.ш., 35°39′ в.д., Сусита (18) |
| 19 | 15s | 193s | 32°47′ с.ш., 35°38′ в.д. (19), Нахаль Ен Гев | |
| 20 | » | 20s + 53s + 94s + 109s + 162s + + 163s + 164s + 165s | 47s + 150s + 165s + 196s + + 201s + 202s + 248s + 254s | Кфар Харув (20) |
| 21 | Оригинальные сборы | 16s + 152s + 153s + 154s + + 155s + 156s | 85s + 156s + 194s + 195s + + 196s + 260s | Неот а-Галан (21) |
| 22 | » | 33 + 117s | 165s + 197s + 198s | 32°40′ с.ш., 35°39′ в.д. (22), Хамат Гаден |
| 23 | » | 15s + 157s | 47s + 165s | 5 км от Хамат Гаден (23) |
| 24 | » | 1 + 12 + 30s + 64s + 112s + +148s + 149s | 165s + 171s + 184s + 185s + + 186s + 188s + 252s | Хайфа, Технион (24) |
| 25 | » | 133s + 134s | 217 + 161s + 170s + 171s + 172s | а-Боним (25) |
| 26 | » | 67s + 133s + 135s + 161s | 19s + 98s + 173s + 174s | Кармель, кибуцу Офер (26) |
| 27 | » | 33 + 63 + 67 + 1s + 25s + + 126s + 178s + 179s | 150 + 163 + 134s + 195s + + 214s + 217s + 218s | 32°37′ с.ш., 34°59′ в.д., Мицпе Офир (27) |
| 28 | » | 40 + 123 + 152 | 142 + 12s + 160s + 161s | 32°28′ с.ш., 34°53′ в.д., Кесария (28) |
| 29 | » | 35 + 136s + 137s | 1 + 175s + 176s | 32°29′ с.ш., 35°29′ в.д., Бет Шиан (29) |
| 30 | » | 143s + 144s + 145s + 146s | 29 + 228 + 181s + 245s | Мехола (30) |
| 31 | » | 1 + 168s + 169s | 16 + 200s + 205s | Эльром (31) |
| H. vulgare | ||||
| 32 | к-3453 | 2 | 1 + 19 + 53 | Палестина, 1922 г. |
| 33 | к-4606 | 2 + 3 + 24 + 32 | 35 + 53 + 82 + 277 | Палестина, 1923 г. |
| 34 | к-4620 | 17 | 18 | » |
| 35 | к-4621 | 2 + 24 | 198 + 218 | » |
| 36 | к-5272 | 18 | 18 | Палестина, 1922 г. |
| 37 | к-5273 | 2 + 3 + 18 | 17 + 18 + 31 | » |
| 38 | к-5274 | 2 + 3 + 159 | 3 + 6 + 24 | » |
| 39 | к-7417 | 2 + 28 + 159 | 3 + 17 + 19 + 24 | Вифлеем, долина Пастухов, 1926 г. (32) |
| 40 | к-7418 | 2 | 14 | Хеврон (33) |
| 41 | к-7426 | 2 + 20 + 28 | 1 + 20 + 278 + 279 | Елеонская гора (34) |
| 42 | к-7432 | 2 + 160 | 14 + 17 + 18 | гора Бейт Джала (35) |
| 43 | к-8084 | 2 | 119 + 280 + 281 | Назарет- базар |
| 44 | к-8085 | 2 + 20 + 41 | 11 + 20 + 53 | То же |
* В скобках указаны номера точек сборов на рис. 1; символом “s” обозначены варианты гордеинов А и В, к настоящему времени обнаруженные исключительно в H. spontaneum.
Электрофорез гордеинов проводили в столбиках 12–14%-ного крахмального геля с 3 М мочевиной в алюминий-лактатном буфере с рН 3.1 по методике А.А. Созинова и Ф.А. Поперели [20] с некоторыми модификациями [19]. С целью изучения возможной дифференциации или сходства исследованных популяций H. spontaneum как межу собой, так и с образцами H. vulgare, проводили иерархический кластерный анализ по методу ближайшего соседа с использованием корреляций Пирсона в качестве меры близости распределений аллельных частот [21]. Контроль за результатами анализа при подборе адекватного алгоритма кластеризации осуществляли с помощью грубой дендрограммы, построенной по матрице числа общих аллелей. Вычисления проводились с помощью пакета статистических программ SPSS 19.0.0.1.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В результате электрофоретического анализа гордеинов в образцах дикого и культурного ячменя из Израиля и Палестины нами было обнаружено 114 и 11 аллелей локуса Hrd A соответственно (рис. 2,а, в). Для локуса Hrd B в образцах H. spontaneum было выявлено также114 аллелей, а в образцах H. vulgare – 22 аллеля (рис. 2,б, г). Генетические формулы гордеина в исследованных образцах дикого и культурного ячменя представлены в табл. 1. Присутствующие в популяциях H. spontaneum из Израиля аллели локусов Hrd A и Hrd B значительно различались по частотам (табл. 2). Среди аллелей локуса Hrd A у дикого ячменя по частоте выделялся аллель А1 (0.0910). С относительно высокой частотой встречались также аллели А30s (0.0539), А33 (0.0520). Частоты остальных аллелей варьировали в пределах 0.0019–0.0427. В образцах H. spontaneum также впервые были обнаружены 74 аллеля (А112s – А185s), не встречавшиеся у исследованных нами ранее образцов дикого ячменя из Ирана [13], Турции [14], Сирии [15] и Иордании [16].
Рис. 2.
Варианты блоков компонентов гордеинов, контролируемых аллелями локусов Hrd A и Hrd B, обнаруженные в популяциях H. spontaneum (а, б) и H. vulgare (в, г) из Израиля и Палестины.

Таблица 2.
Частоты аллелей локусов Hrd A и Hrd B в образцах дикого (H. spontaneum) и культурного (H. vulgare) ячменя из Израиля и Палестины
| Аллели Hrd A |
H. spontaneum | H. vulgare | Аллели Hrd A |
H. spontaneum | H. vulgare |
| 1 | 0.0910 | 0 | 126s | 0.0037 | 0 |
| 2 | 0.0019 | 0.4215 | 127s | 0.0019 | 0 |
| 3 | 0.0074 | 0.0714 | 128s | 0.0019 | 0 |
| 12 | 0.0186 | 0 | 129s | 0.0056 | 0 |
| 13 | 0.0019 | 0 | 130s | 0.0019 | 0 |
| 17 | 0 | 0.0714 | 131s | 0.0037 | 0 |
| 18 | 0.0260 | 0.1714 | 132s | 0.0037 | 0 |
| 20 | 0 | 0.0143 | 133s | 0.0167 | 0 |
| 23 | 0.0037 | 0 | 134s | 0.0093 | 0 |
| 24 | 0.0037 | 0.05 | 135s | 0.0019 | 0 |
| 28 | 0.0019 | 0.05 | 136s | 0.0019 | 0 |
| 32 | 0 | 0.0214 | 137s | 0.0019 | 0 |
| 33 | 0.0520 | 0 | 138s | 0.0056 | 0 |
| 35 | 0.0037 | 0 | 139s | 0.0019 | 0 |
| 40 | 0.0130 | 0 | 140s | 0.0019 | 0 |
| 41 | 0 | 0.0571 | 141s | 0.0019 | 0 |
| 61 | 0.0019 | 0 | 142s | 0.0019 | 0 |
| 63 | 0.0130 | 0 | 143s | 0.0019 | 0 |
| 67 | 0.0019 | 0 | 144s | 0.0019 | 0 |
| 110 | 0.0093 | 0 | 145s | 0.0019 | 0 |
| 123 | 0.0093 | 0 | 146s | 0.0037 | 0 |
| 152 | 0.0409 | 0 | 147s | 0.0019 | 0 |
| 159 | 0 | 0.0429 | 148s | 0.0112 | 0 |
| 160 | 0 | 0.0286 | 149s | 0.0093 | 0 |
| 1s | 0.0019 | 0 | 150s | 0.0279 | 0 |
| 10s | 0.0074 | 0 | 151s | 0.0037 | 0 |
| 11s | 0.0019 | 0 | 152s | 0.0093 | 0 |
| 15s | 0.0242 | 0 | 153s | 0.0112 | 0 |
| 16s | 0.0019 | 0 | 154s | 0.0056 | 0 |
| 20s | 0.0019 | 0 | 155s | 0.0037 | 0 |
| 25s | 0.0149 | 0 | 156s | 0.0019 | 0 |
| 30s | 0.0539 | 0 | 157s | 0.0056 | 0 |
| 39s | 0.0019 | 0 | 158s | 0.0037 | 0 |
| 41s | 0.0019 | 0 | 159s | 0.0204 | 0 |
| 52s | 0.0019 | 0 | 160s | 0.0019 | 0 |
| 53s | 0.0056 | 0 | 161s | 0.0130 | 0 |
| 55s | 0.0335 | 0 | 162s | 0.0130 | 0 |
| 64s | 0.0335 | 0 | 163s | 0.0037 | 0 |
| 67s | 0.0019 | 0 | 164s | 0.0037 | 0 |
| 91s | 0.0019 | 0 | 165s | 0.0037 | 0 |
| 94s | 0.0019 | 0 | 166s | 0.0019 | 0 |
| 96s | 0.0019 | 0 | 167s | 0.0019 | 0 |
| 105s | 0.0074 | 0 | 168s | 0.0019 | 0 |
| Аллели Hrd A |
H. spontaneum | H. vulgare | Аллели Hrd A |
H. spontaneum | H. vulgare |
| 106s | 0.0019 | 0 | 169s | 0.0130 | 0 |
| 107s | 0.0019 | 0 | 170s | 0.0093 | 0 |
| 109s | 0.0056 | 0 | 171s | 0.0037 | 0 |
| 112s | 0.0427 | 0 | 172s | 0.0019 | 0 |
| 113s | 0.0056 | 0 | 173s | 0.0019 | 0 |
| 114s | 0.0074 | 0 | 174s | 0.0186 | 0 |
| 115s | 0.0037 | 0 | 175s | 0.0093 | 0 |
| 116s | 0.0316 | 0 | 176s | 0.0037 | 0 |
| 117s | 0.0149 | 0 | 177s | 0.0056 | 0 |
| 118s | 0.0056 | 0 | 178s | 0.0019 | 0 |
| 119s | 0.0037 | 0 | 179s | 0.0019 | 0 |
| 120s | 0.0019 | 0 | 180s | 0.0056 | 0 |
| 121s | 0.0019 | 0 | 181s | 0.0335 | 0 |
| 122s | 0.0019 | 0 | 182s | 0.0037 | 0 |
| 123s | 0.0019 | 0 | 183s | 0.0074 | 0 |
| 124s | 0.0019 | 0 | 184s | 0.0019 | 0 |
| 125s | 0.0019 | 0 | 185s | 0.0019 | 0 |
| Аллели Hrd B |
H. sponaneum | H. vulgare | Аллели Hrd B |
H. sponaneum | H. vulgare |
| 1 | 0.0037 | 0.0214 | 161s | 0.0558 | 0 |
| 3 | 0 | 0.0286 | 162s | 0.0074 | 0 |
| 6 | 0 | 0.0214 | 163s | 0.0149 | 0 |
| 8 | 0 | 0.0071 | 164s | 0.0037 | 0 |
| 11 | 0 | 0.0571 | 165s | 0.0650 | 0 |
| 14 | 0 | 0.1000 | 166s | 0.0056 | 0 |
| 15 | 0.0074 | 0 | 167s | 0.0019 | 0 |
| 16 | 0.0019 | 0 | 168s | 0.0019 | 0 |
| 17 | 0 | 0.0571 | 169s | 0.0019 | 0 |
| 18 | 0 | 0.2000 | 170s | 0.0037 | 0 |
| 19 | 0 | 0.0500 | 171s | 0.0112 | 0 |
| 20 | 0 | 0.0429 | 172s | 0.0056 | 0 |
| 24 | 0 | 0.0429 | 173s | 0.0019 | 0 |
| 29 | 0.0037 | 0 | 174s | 0.0093 | 0 |
| 31 | 0 | 0.0143 | 175s | 0.0019 | 0 |
| 35 | 0 | 0.0071 | 176s | 0.0019 | 0 |
| 41 | 0.0037 | 0 | 177s | 0.0019 | 0 |
| 53 | 0 | 0.0787 | 178s | 0.0019 | 0 |
| 65 | 0.0037 | 0 | 179s | 0.0019 | 0 |
| 119 | 0 | 0.0571 | 180s | 0.0093 | 0 |
| 141 | 0.0019 | 0 | 181s | 0.0019 | 0 |
| 142 | 0.0074 | 0 | 182s | 0.0112 | 0 |
| 150 | 0.0019 | 0 | 183s | 0.0019 | 0 |
| 162 | 0.0223 | 0 | 184s | 0.0093 | 0 |
| 163 | 0.0019 | 0 | 185s | 0.0669 | 0 |
| Аллели Hrd B |
H. sponaneum | H. vulgare | Аллели Hrd B |
H. sponaneum | H. vulgare |
| 169 | 0.0037 | 0 | 186s | 0.0743 | 0 |
| 189 | 0.0019 | 0 | 187s | 0.0019 | 0 |
| 198 | 0 | 0.1144 | 188s | 0.0372 | 0 |
| 217 | 0.0037 | 0 | 189s | 0.0093 | 0 |
| 218 | 0 | 0.0286 | 190s | 0.0019 | 0 |
| 228 | 0.0019 | 0 | 191s | 0.0279 | 0 |
| 277 | 0 | 0.0357 | 192s | 0.0093 | 0 |
| 278 | 0 | 0.0143 | 193s | 0.0186 | 0 |
| 279 | 0 | 0.0071 | 194s | 0.0112 | 0 |
| 280 | 0 | 0.0071 | 195s | 0.0056 | 0 |
| 281 | 0 | 0.0071 | 196s | 0.0056 | 0 |
| 12s | 0.0130 | 0 | 197s | 0.0149 | 0 |
| 15s | 0.0037 | 0 | 198s | 0.0112 | 0 |
| 19s | 0.0019 | 0 | 199s | 0.0019 | 0 |
| 33s | 0.0019 | 0 | 200s | 0.0186 | 0 |
| 47s | 0.0074 | 0 | 201s | 0.0112 | 0 |
| 85s | 0.0019 | 0 | 202s | 0.0037 | 0 |
| 87s | 0.0037 | 0 | 203s | 0.0056 | 0 |
| 97s | 0.0093 | 0 | 204s | 0.0019 | 0 |
| 98s | 0.0019 | 0 | 205s | 0.0112 | 0 |
| 123s | 0.0019 | 0 | 206s | 0.0149 | 0 |
| 134s | 0.0037 | 0 | 207s | 0.0037 | 0 |
| 140s | 0.0037 | 0 | 208s | 0.0074 | 0 |
| 141s | 0.0056 | 0 | 209s | 0.0019 | 0 |
| 142s | 0.0019 | 0 | 210s | 0.0037 | 0 |
| 143s | 0.0019 | 0 | 211s | 0.0056 | 0 |
| 144s | 0.0019 | 0 | 212s | 0.0093 | 0 |
| 145s | 0.0019 | 0 | 213s | 0.0279 | 0 |
| 146s | 0.0056 | 0 | 214s | 0.0019 | 0 |
| 147s | 0.0037 | 0 | 215s | 0.0019 | 0 |
| 148s | 0.0019 | 0 | 217s | 0.0149 | 0 |
| 149s | 0.0074 | 0 | 218s | 0.0019 | 0 |
| 150s | 0.0037 | 0 | 220s | 0.0056 | 0 |
| 151s | 0.0019 | 0 | 221s | 0.0186 | 0 |
| 152s | 0.0112 | 0 | 222s | 0.0074 | 0 |
| 153s | 0.0019 | 0 | 224s | 0.0353 | 0 |
| 154s | 0.0019 | 0 | 245s | 0.0019 | 0 |
| 155s | 0.0037 | 0 | 247s | 0.0056 | 0 |
| 156s | 0.0093 | 0 | 248s | 0.0056 | 0 |
| 157s | 0.0019 | 0 | 252s | 0.0056 | 0 |
| 158s | 0.0056 | 0 | 254s | 0.0019 | 0 |
| 159s | 0.0019 | 0 | 260s | 0.0093 | 0 |
| 160s | 0.0279 | 0 |
Из 114 аллелей локуса Hrd В, выявленных в образцах дикого ячменя из Израиля, самую высокую частоту имели четыре аллеля: B186s (0.0743), B185s (0.0669), В165s (0.0650) и B161s (0.0558). С относительно высокой частотой встречались аллели В188s (0.0372) и В224s (0.0353). Частоты остальных 108 аллелей находились в пределах 0.0019–0.0279 (табл. 2). Для этого локуса 82 аллеля (B140s – B215s, B217s, B218s, B220s, B221s, B222s и B224s) к настоящему времени были обнаружены только в израильских популяциях H. spontaneum.
В образцах культурного ячменя из Палестины и Израиля для локуса Hrd A были найдены 11 аллелей, из которых по частоте преобладал аллель А2 (0.4215). Относительно высокую частоту имели аллели А18 (0.1714), А3, А17 (0.0714) и А41 (0.0571). Частоты остальных шести аллелей этого локуса варьировали в пределах 0.0143–0.0500. В исследованных образцах для локуса Hrd B были обнаружены 22 аллеля, из которых три характеризовались наибольшей частотой: В18 (0.2000), В198 (0.1144) и В14 (0.1000). Сравнительно высокую частоту имели таке аллели В53 (0.0787), В11, В17 и В119 (0.0571). Частоты остальных 15 аллелей находились в пределах 0.0071–0.0429 (табл. 2).
В результате проведенного кластерного анализа большинство (74%) популяций дикого ячменя и представленные как одна популяция образцы культурного ячменя распределились по четырем основным кластерам (рис. 3). В первый, самый большой кластер I вошли 11 популяций дикого ячменя и объединенная популяция местных сортов культурного ячменя. Преимущественно в этот кластер объединились образцы H. spontaneum из района оз. Кинерет (точки сборов 22, 13, 18, 19, 23, 16, 31, 11, 12), одна популяция из района Эль Кунейтра на северо-востоке Израиля (6) и одна популяция из района Хайфы в северо-западной части страны – точка сбора 24 (рис. 1, 3). Только в двух популяциях, образующих этот кластер (точки сборов 19 и 23), отсутствовали аллели локусов Hrd А и Hrd В, которые нами ранее были идентифицированы в культурном ячмене различных стран [19]. При этом в образцах дикого ячменя из точек сборов 12, 16 и 18 присутствовали аллели гордеин-кодирующих локусов, обнаруженные и в образцах культурного ячменя из Израиля (табл. 1). Второй по величине кластер III составили пять популяций из района оз. Кинерет (точки сборов 7, 17, 8, 9, 20) и одна популяция из района Мицпе Пеэр на севере Израиля (точка 1). Только в образцах из района Авней Эйтан (17) и Кешет (8) были обнаружены растения с аллелем В162, который ранее мы находили в образцах местного культурного ячменя из мировой коллекции. Кластер IV включает две популяции из района оз. Кинерет (точки 10 и 15) и популяции из района а-Боним (25) и района Кармель, кибуцу Офер (26). В популяции из Табигхи (10) присутствовал аллель А24, обнаруженный также в образцах культурного ячменя из Израиля и Палестины. Кластер II представлен всего двумя популяциями: из района Баниас на севере Израиля (5) и района Кесарии около Средиземноморского побережья страны (28). В обеих популяциях найдены аллели локусов Hrd А и Hrd В, идентифицированные нами ранее в культурном ячмене из мировой коллекции. Остальные восемь популяций H. spontaneum из различных районов Израиля не входили ни в один из кластеров. В пяти из них также обнаружены аллели гордеин-кодирующих локусов, идентифицированные нами в образцах из мировой коллекции ячменя [19]. Таким образом, “культурные” аллели локусов Hrd А и Hrd В присутствовали в 21 из 31 изученной популяции H. spontaneum из Израиля и Палестины.
Рис. 3.
Дендрограмма для 32 популяций H. spontaneum и H. vulgare по методу ближайшего соседа с использованием корреляции Пирсона в качестве меры близости (SPSS). I–IV – кластеры, арабские цифры – номера точек сбора на рис. 1.

Если принять гипотезу, что доместикация ячменя произошла на территории современных Израиля и Палестины, можно было бы ожидать присутствие в образцах дикого и культурного ячменя одних и тех же аллелей локусов Hrd А и Hrd В, а также присутствие в диком ячмене представителей самых широко распространенных аллелей этих локусов не только в местных образцах культурного ячменя в Израиле и Палестине, но и в других странах мира. Среди 114 аллей локуса Hrd A, идентифицированных в образцах H. spontaneum, 18 аллелей были нами ранее обнаружены в мировой коллекции культурного ячменя [19]. Однако в исследованных образцах культурного и дикого ячменя из Израиля и Палестины общими для них являлись только пять аллелей: А2, А3, А18, А24 и А28. Их частоты значительно различались в культурном и диком ячмене и находились в пределах 0.05–0.4215 и 0.0019–0.026 соответственно (табл. 2). Для локуса Hrd B в образцах дикого ячменя было выявлено 15 “культурных” аллелей. Однако только один из них – B1, частота которого в H. spontaneum составила 0.0037, присутствовал в палестинских образцах культурного ячменя с частотой 0.0214.
В табл. 3 приведены частоты “культурных” аллелей локуса Hrd A и Hrd B, обнаруженных в образцах H. spontaneum из Израиля и Палестины и в исследованных ранее местных образцах культурного ячменя из 20 стран мира, входящих в центры разнообразия ячменя и сопредельных с ними [14, 22–28]. Как видно из таблицы, среди аллелей локуса Hrd A самым распространенным является аллель А2, обнаруженный в местных образцах культурного ячменя из 19 стран мира с высокими частотами – от 0.0719 в Йемене до 0.8000 в Тунисе. В Израиле этот аллель идентифицирован только в одном растении H. spontaneum популяции из Арбеля. В образцах культурного ячменя из 13 различных стран были найдены аллели А3 и А24, где их частоты варьировали от 0.0266 (Иордания) до 0.2334 (Марокко) и от 0.0032 (Иран) до 0.4572 (Эфиопия) соответственно. В диком ячмене из Израиля аллель А3 был найден только в двух популяциях: из районов Галмы (16) и Сусита (18) около оз. Кинерет (табл. 1, рис. 1). Аллель А18 присутствовал в образцах H. vulgare из 12 стран Северной Африки, Средней и Восточной Азии с частотой от 0.0307 (Египет) до 0.6441 (Узбекистан). В Израиле этот аллель был найден только в одной популяции H. spontaneum из района Галмы (16). Каждый из аллелей А1, А12 и А33 был обнаружен в культурном ячмене из 10 стран. При этом аллели А1 и А12 отсутствовали в образцах из стран Северной Африки. Максимальную частоту 0.14 аллель А1 имел в культурном ячмене из Сирии, а аллель А12 (0.1362) – в Киргизии. Для аллеля А33 самые высокие частоты отмечены в Иордании (0.2707), Сирии (0.2250) и Марокко (0.1222). Аллель А1 обнаружен в израильских популяциях дикого ячменя из районов Табигха (11), Арбель (12), Галма (16), Сусита (18) и Эльром (31), находящихся около оз. Кинерет, и из района Хайфы (табл. 1, рис. 1). Аллель А12 был найден в двух популяциях H. spontaneum, собранных в районе оз. Кинерет–Сусита (18), Арбель (12), и в популяции из окрестностей Хайфы (24). Аллель А33 присутствовал в популяциях израильского дикого ячменя из Казрин (13), Сусита (18), Хамат Гаден (22) – район оз. Кинерет, а также в образце из Мицпе Офир (27). Аллели А13, А23, А28, А61, А63, А67 А110, А123 и А152 в культурном ячмене из различных стран распространены на значительно меньшей территории и с низкими частотами. Эти аллели присутствовали в единичных популяциях израильского H. spontaneum, преимущественно из района оз. Кинерет, за исключением аллелей А63, А67 и А123, найденных и в образцах из Мицпе Офир (27), и Кесария (28). Мы не рассматривали распространение аллелей А35 и А40, поскольку они были нами ранее обнаружены только в отдельных современных сортах ячменя европейской селекции.
Таблица 3.
Частоты аллелей локусов Hrd A и Hrd B, обнаруженных в H. spontaneum из Израиля и Палестны, также присутствующие в образцах местных культурных ячменей из стран Северной и Северо-Восточной Африки, Ближнего Востока, Передней, Центральной и Восточной Азии
| Страна | Аллели локуса Hrd A | |||||||||||||||
| 1 | 2 | 3 | 12 | 13 | 18 | 23 | 24 | 28 | 33 | 61 | 63 | 67 | 110 | 123 | 152 | |
| Марокко | 0.0 | 0.1000 | 0.2334 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.1222 | 0.0 | 0.0 | 0.0778 | 0.0 | 0.0 | 0.0 |
| Тунис | 0.0 | 0.8000 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Алжир | 0.0 | 0.2545 | 0.1455 | 0.0 | 0.1273 | 0.0455 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0636 | 0.0091 | 0.0 | 0.0 | 0.0 |
| Египет | 0.0 | 0.3872 | 0.0 | 0.0 | 0.0256 | 0.0307 | 0.0 | 0.0333 | 0.0256 | 0.0 | 0.0 | 0.0 | 0.0256 | 0.0 | 0.0 | 0.0 |
| Турция | 0.0022 | 0.2934 | 0.0946 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0527 | 0.0172 | 0.0419 | 0.0 | 0.0 | 0.0011 | 0.0043 | 0.0 | 0.015 |
| Эфиопия | 0.0068 | 0.2174 | 0.0 | 0.0085 | 0.0 | 0.0 | 0.0 | 0.4572 | 0.0 | 0.0718 | 0.0308 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Йемен | 0.0 | 0.0719 | 0.0 | 0.0 | 0.0 | 0.0011 | 0.0 | 0.1180 | 0.0112 | 0.0438 | 0.0011 | 0.0 | 0.0 | 0.0 | 0.009 | 0.0 |
| Афганистан | 0.0095 | 0.1405 | 0.0667 | 0.0167 | 0.0012 | 0.3273 | 0.0036 | 0.0226 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ирак | 0.0059 | 0.2412 | 0.0589 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0088 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Иордания | 0.0911 | 0.1382 | 0.0266 | 0.0029 | 0.0147 | 0.0 | 0.0266 | 0.0088 | 0.0118 | 0.2707 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Сирия | 0.1400 | 0.1466 | 0.0567 | 0.0033 | 0.0 | 0.0 | 0.0 | 0.0067 | 0.0 | 0.2250 | 0.0017 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Иран | 0.0730 | 0.02826 | 0.0968 | 0.0159 | 0.0127 | 0.054 | 0.0 | 0.0032 | 0.0063 | 0.0190 | 0.0 | 0.0 | 0.0 | 0.0365 | 0.0 | 0.0 |
| Туркмения | 0.0020 | 0.2116 | 0.0784 | 0.0314 | 0.002 | 0.2019 | 0.0 | 0.0 | 0.0 | 0.0018 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Узбекистан | 0.0 | 0.0227 | 0.0394 | 0.0 | 0.0 | 0.6441 | 0.0039 | 0.0059 | 0.0 | 0.0049 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Киргизия | 0.0170 | 0.3360 | 0.0511 | 0.1362 | 0.0 | 0.1276 | 0.0 | 0.0 | 0.0213 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Таджикистан | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0739 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Китай | 0.0125 | 0.2854 | 0.0187 | 0.0333 | 0.0 | 0.1417 | 0.0 | 0.0146 | 0.0021 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Непал | 0.0 | 0.1800 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Пакистан | 0.0 | 0.1214 | 0.0 | 0.0143 | 0.0 | 0.3929 | 0.0 | 0.0214 | 0.0 | 0.0143 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Индия | 0.0 | 0.2 | 0.0467 | 0.0042 | 0.0123 | 0.0383 | 0.0 | 0.0255 | 0.0 | 0.0 | 0.0042 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Страна | Аллели локуса Hrd В | Литературная ссылка | ||||||||||||||
| 1 | 15 | 16 | 29 | 41 | 65 | 141 | 142 | 150 | 162 | 163 | 169 | 189 | 217 | 228 | ||
| Марокко | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 22 |
| Тунис | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Алжир | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Египет | 0.0 | 0.0 | 0.0256 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Турция | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0011 | 0.0 | 0.0096 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 14 |
| Эфиопия | 0.0250 | 0.0 | 0.0068 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 23 |
| Йемен | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0539 | 0.0011 | 0.0011 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0067 | 28 |
| Афганистан | 0.0048 | 0.0095 | 0.0024 | 0.0036 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0059 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 24 |
| Ирак | 0.0091 | 0.0061 | 0.0 | 0.0 | 0.0 | 0.0706 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 25 |
| Иордания | 0.0088 | 0.0059 | 0.0 | 0.0266 | 0.0 | 0.1824 | 0.0176 | 0.0029 | 0.0 | 0.0118 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Сирия | 0.0050 | 0.2670 | 0.0 | 0.0 | 0.0 | 0.2182 | 0.0133 | 0.0217 | 0.0 | 0.0 | 0.0 | 0.0133 | 0.0 | 0.0 | 0.0 | |
| Иран | 0.0095 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0254 | 0.0 | 0.0 | 26 |
| Туркмения | 0.0059 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0176 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Узбекистан | 0.0020 | 0.0 | 0.0 | 0.0039 | 0.0 | 0.0020 | 0.0 | 0.0 | 0.0 | 0.0 | 0.001 | 0.0 | 0.001 | 0.0039 | 0.0 | |
| Киргизия | 0.2510 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0043 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Таджикистан | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Китай | 0.0396 | 0.0 | 0.1146 | 0.0 | 0.0 | 0.0 | 0.0416 | 0.0021 | 0.0146 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 27 |
| Непал | 0.0 | 0.0 | 0.1000 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Пакистан | 0.0 | 0.0 | 0.0714 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Индия | 0.0 | 0.0 | 0.0213 | 0.0 | 0.029 | 0.0213 | 0.0 | 0.0213 | 0.0128 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
Среди 15 “культурных” аллелей локуса Hrd B, найденных в образцах H. spontaneum из Израиля, относительно широкое распространение в культурном ячмене имел аллель В1, который с низкими частотами (от 0.002 до 0.025) присутствовал в культурном ячмене из 10 стран Северо-Восточной Африки Средней Азии, Ближнего Востока (табл. 3). Самые высокие частоты этого аллеля отмечены в Киргизии (0.2510) и Китае (0.0396). Аллель B16 с наибольшими частотами встречался в культурном ячмене из Китая (0.1146), Непала (0.1000) и Пакистана (0.0714), а в образцах из остальных четырех стран его частота была в пределах 0.0024 (Афганистан) – 0.0256 (Египет). В израильском H. spontaneum этот аллель обнаружен только в одной популяции из Эльрома (31). Аллель В65 преимущественно распространен в культурном ячмене стран Ближнего Востока, где его частота варьировала в пределах 0.0706 (Ирак) – 0.2182 (Сирия). С низкими частотами он присутствовал в культурном ячмене из Турции (0.0011), Узбекистана (0.0020) и Индии (0.0213). При этом в диком ячмене из Израиля этот аллель обнаружен только в одной популяции из Суситы (18). Остальные “культурные” аллели этого локуса характеризовались мозаичным характером распространения в H. vulgare и низкими частотами в различных странах мира. В диком израильском ячмене эти аллели встречались в единичных популяциях в различных районах страны. Обращает на себя внимание популяция H. spontaneum из Суситы (18), в которой обнаружено девять “культурных” аллелей для локуса Hrd A и три – для локуса Hrd B. Кроме этого, генотипы Hrd A3 B162 и Hrd A24 B65, присутствующие в этой популяции, обнаружены нами ранее в отдельных образцах культурного ячменя только из Иордании и Сирии [25]. Учитывая, что гордеин-кодирующие локусы наследуются сцепленно – величина рекомбинации между ними составляет 12.6% [29], а также близкое расположение района оз. Кинерет к границам Сирии и Иордании, можно предположить, что указанные генотипы могли появиться в культурном ячмене в результате интрогресси популяции H. spontaneum из Суситы или каких-то других популяций дикого ячменя из этого района Израиля. Следует отметить, что подавляющее большинство “культурных” аллелей локусов Hrd A и Hrd B в исследованных популяциях H. spontaneum встречаются в одном генотипе с аллелями этих локусов, обнаруженных до сих пор только в диком ячмене.
Таким образом, представленные данные об аллельном разнообразии гордеин-кодирующих локусов в популяциях израильского дикого ячменя, составе и распространении обнаруженных “культурных” аллелей и отдельных генотипов в местных образцах культурного ячменя из 20 стран позволяют сделать вывод, что некоторые популяции H. spontaneum из Израиля могли быть донорами отдельных аллелей и генотипов гордеин-кодирующих локусов для H. vulgare в результате интрогрессии при спонтанной гибридизации при распространении культурного ячменя.
Авторы выражают благодарность О. Раскиной, А. Беляеву и Е. Бадаевой за предоставленный материал H. spontaneum из Израиля.
Работа выполнена по государственному заданию № 0112-2019-0002.
Настоящая статья не содержит каких-либо исследований с использованием в качестве объекта животных.
Настоящая статья не содержит каких-либо исследований с участием в качестве объекта людей.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Harlan J.R., Zohary D. Distribution of wild wheats and barley // Science. 1966. V. 153. № 3740. P. 1074–1080. https://doi.org/10.1126/science.153.3740.1074
Zohary D., Hopf M. Domestication of Plants in the Old World. The Origin and Spread of Cultivated Plants in West Asia, Europe, and Nile valley. Oxford: Clarendon Press, 1988. 245 p.
Бахтеев Ф.Х. К истории культуры ячменя в СССР // Матер. по истории земледелия СССР: Cб. II. М.-Л.: Изд-во АН СССР, 1956. С. 204–257.
Вавилов Н.И. Учение о происхождении культурных растений после Дарвина // Сов. наука. 1940. № 2. С. 55–75.
Åberg E. Hordeum agriocrithon nova sp. a wild six rowed barley // Ann. Roy. Agr. Col. Sweden. 1938. V. 6. P. 159–216.
Zhang Q., Saghai Maroof M.A., Yang P.G. Ribosomal DNA polymorphisms and the Oriental-Occidental genetic differentiation in cultivated barley // Theor. Appl. Genet. 1992. V. 84. № 5. P. 682–687. https://doi.org/10.1007/BF00224168
Molino-Cano J.-L., Fra-Mon P., Salcedo G. et al. Marocco as possible domestication centre for barley: biochemical and agromorphological evidence // Theor. Appl. Genet. 1987. V. 73. № 4. P. 531–536. https://doi.org/10.1007/BF00289190
Negassa M. Patterns of phenotypic diversity in an Ethiopian barley collection, and the Arussi-Bale Higland as a center of origin ob barley // Hereditas. 1985. V. 102. № 1. P. 139–150. https://doi.org/10.1111/j.1601-5223.1985.tb00474.x
Lev-Yadum S., Gopher A., Abbo S. The cradle of agriculture // Science. 2000. V. 288. P. 1602–1603. https://doi.org/10.1126/science.288.5471.1602
Kilian B., Özkan H., Kohl J. et al. Haplotype structure at seven barley genes: relevance to gene pool bottlenecks, phylogeny of ear type and site of barley domestication // Mol. Gen. Genomics. 2006. V. 276. P. 230–241. https://doi.org/10.1007/s00438-006-0136-6
Morrell P.L., Clegg M.T. Genetic evidence for a second domestication of barley (Hordeum vulgare) east of the Fertile Crescent // Proc. Natl Acad. Sci. USA. 2007. V. 104. № 9. P. 3289–3294.
Poets A.M., Fang Z., Clegg M.T., Morrell P.L. Barley landraces are characterized by geographically heterogeneous genomic origin // Genome Biol. 2015. V. 16. P. 173. https://doi.org/10.1186/s13059-015-0712-3
Поморцев А.А., Лялина Е.В. Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L) и дикого (H. spontaneum C. Koch) ячменя в Иране (как части Дуги Плодородия) // Генетика. 2016. Т. 52. №10. С. 1146–1158. https://doi.org/10.7868/S001667581610009X
Поморцев А.А., Болдырев С.В., Лялина Е.В. Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L.) и дикого (H. spontaneum C. Koch) ячменя в Турции (как части Дуги Плодородия) // Генетика. 2017. Т. 53. № 4. С. 449–459. https://doi.org/10.7868/S0016675817030092
Поморцев А.А., Болдырев С.В., Лялина Е.В. Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L.) и дикого (Hordeum spontaneum C. Koch) ячменя в Сирии (как части Дуги плодородия) // Генетика. 2018. Т. 54. № 11. С. 1275–1284. https://doi.org/10.1134/S0016675818110127
Поморцев А.А., Болдырев С.В., Лялина Е.В. Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L.) и дикого (Hordeum spontaneum C. Koch) ячменя в Иордании (как части Дуги плодородия) // Генетика. 2019. Т. 55. № 3. С. 290–299. https://doi.org/10.1134/S0016675819030135
Willcox G. Archeobotanical evidence for the beginnings of agriculture in Southwest Asia // Proc. Harlan Symp. The Origins of Agriculture and Crop Domestication. Aleppo, 10–14 May, 1997. P. 25–38.
Лисицына Г.Н., Прищепенко Л.В. Палеоботанические находки Кавказа и Ближнего Востока. М.: Наука, 1977. 115 с.
Поморцев А.А., Лялина Е.В. Оценка сортовой принадлежности и сортовой чистоты семян ячменя методом электрофоретического анализа запасных белков зерна // Методич. пособие к практикуму “Белковые маркеры для генетической паспортизации и улучшения геномов растений хозяйственно ценных видов”. М.: Цифровичок, 2011. 86 с.
Созинов А.А., Попереля Ф.А. Методика вертикального дискового электрофореза в крахмальном геле и генетический принцип классификации глиадинов. Одесса, 1978. 16 с.
Hennig C., Meila M., Murtagh F., Rocci R. Handbook of Cluster Analysis. CRC Press, a Chapman & Hall book. 2016. 730 p.
Поморцев А.А., Калабушкин Б.А., Терентьева И.А. Полиморфизм гордеинов у ячменей Северной Африки // Генетика. 2002. Т. 38. № 11. С. 1498–1510.
Поморцев А.А. Исследование полиморфизма гордеинов у ячменей Эфиопии // Генетика. 2001. Т. 37. № 10. С. 1371–1382.
Поморцев А.А., Мартынов С.П., Ковалева О.Н., Лялина Е.В. Полиморфизм по гордеин-кодирующим локусам культурного ячменя (Hordeum vulgare L.) Афганистана // Генетика, 2010. Т. 46. № 11. С. 1507–1515. https://doi.org/10.1134/S1022795410110086
Поморцев А.А., Мартынов С.П., Лялина Е.В. Полиморфизм гордеин-кодирующих локусов в местных популяциях культурного ячменя (Hordeum vulgare L.) стран Ближнего Востока // Генетика. 2008. Т. 44. № 6. С. 709–721. https://doi.org/10.1134/S1022795408060112
Поморцев А.А., Мартынов С.П., Ковалева О.Н., Лялина Е.В. Полиморфизм по гордеин-кодирующим локусам в популяциях ячменя (Hordeum vulgare L.) Ирана и стран Средней Азии // Генетика. 2011. Т. 47. № 11. С. 1545–1564. https://doi.org/10.1134/S1022795411110147
Поморцев А.А., Мартынов С.П., Лялина Е.В. Полиморфизм по гордеин-кодирующим локусам популяций ячменя (Hordeum vulgare L.) стран Восточной Азии (Китай, Непал, Пакистан, Индия) // Генетика. 2012. Т. 48. № 8. С. 934–950. https://doi.org/10.1134/S1022795412050183
Поморцев А.А., Лялина Е.В. Полиморфизм культурного ячменя (Hordeum vulgare L.) Южной Аравии по гордеин-кодирующим локусам // Генетика. 2007. Т. 43. № 5. С. 660–667.
Созинов А.А., Нецветаев В.П., Григорян Э.М., Образцов И.С. Картирование локусов Hrd у ячменя (Hordeum vulgare L. emed. Vav/ et Bach.) // Генетика. 1978. Т. 14. № 9. С. 1610–1619.
Дополнительные материалы отсутствуют.