

Генетика, 2019, T. 55, № 3, стр. 290-299
Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L.) и дикого (Hordeum spontaneum C. Koch) ячменя в Иордании (как части Дуги Плодородия)
А. А. Поморцев 1, *, С. В. Болдырев 1, Е. В. Лялина 1
1 Институт общей генетики им. Н.И. Вавилова Российской академии наук
119991 Москва, Россия
* E-mail: Pomortsev@vigg.ru
Поступила в редакцию 30.03.2018
После доработки 03.05.2018
Принята к публикации 18.04.2018
Аннотация
Методом электрофореза в крахмальном геле изучен полиморфизм гордеинов, контролируемых локусами Hrd A и Hrd B, в 34 местных образцах культурного и в 19 образцах дикого ячменя из Иордании. Для локуса Hrd A в H. vulgare и H. spontaneum идентифицированы 32 и 26 аллелей, для локуса Hrd B – 42 и 32 аллелей соответственно. Частоты аллелей локусов Hrd A и Hrd B в культурном ячмене варьировали в пределах 0.0029–0.2707 и 0.0029–0.1824 соответственно, а в диком ячмене – в пределах 0.0105–0.1263 и 0.0105–0.0947 соответственно. В иорданских образцах H. spontaneum и H. vulgare из 52 аллелей локуса Hrd A обнаружено шесть общих аллелей, а из 70 аллелей локуса Hrd В – только четыре общих аллеля. Два общих для дикого и культурного ячменя Иордании аллеля локуса Hrd A и один аллель локуса Hrd В были обнаружены ранее и в H. spontaneum из Ирана, Турции и Сирии. Однако три аллеля локуса Hrd A и три аллеля локуса Hrd В идентифицированы одновременно в культурном и диком ячмене только в Иордании. Отмечено, что самое раннее использование ячменя в Иордании датируется 6700 г. до н. э., тогда как в Сирии – 9000 г. до н. э., а в Израиле – 17 тысячелетие до н. э. Сделан вывод, что Иорданию вряд ли можно рассматривать как центр доместикации ячменя. При этом H. spontaneum из Иордании мог являться донором отдельных аллелей гордеин-кодирующих локусов H. vulgare в результате интрогрессии при спонтанной гибридизации.
Изучение происхождения культурных растений, в том числе культурного ячменя, предполагает поиск ответов на вопросы: 1) какой дикий вид (виды) являются предками культурной формы (моно- или полифилетическое происхождение); 2) в каком месте (местах) произошла доместикация (моно- или политопное введение в культуру)? В настоящее время получены убедительные доказательства, что диким предком культурного ячменя является Hordeum spontaneum C. Koch [1]. Однако второй вопрос дискутируется до сих пор на протяжении уже более 130 лет. О центре доместикации ячменя выдвинут целый ряд гипотез: 1) Южная Туркмения; 2) Египет [2]; 3) Юго-Западноазиатский центр [3]; 4) Тибет [4, 5]; 5) Марокко [6]; 6) Эфиопия [7]; 7) Юго-Восточная Турция и Северная Сирия [8]; 8) Дуга Плодородия [9]; 9) долина реки Иордан [10]; 10) Западный Пакистан [11]. Обсуждается так же гипотеза о полицентрическом происхождении культурного ячменя [12]. Некоторые из этих гипотез основываются на имевшихся в то или иное время археологических данных. Другие – на изучении существующего разнообразия культурного и дикого ячменя по полиморфным генетическим маркерам (генам, контролирующим морфологические признаки, белковым и ДНК-маркерам). В настоящее время наиболее популярна гипотеза о введении ячменя в культуру в Дуге Плодородия, проходящей от Хузистана по горам Загрос и Тавр в Иране, Ираке, в Центральной и Западной Анатолии, и южной части Палестины (современная территория Израиля).
У ячменя одной из наиболее полиморфных белковых систем являются спирторастворимые белки эндосперма – гордеины. Известно, что гордеины, выявляемые методом электрофореза в крахмальном геле, контролируются семью локусами – Hrd A, Hrd B, Hrd C, Hrd D, Hrd E, Hrd F, Hrd G, наследуемыми сцепленно и локализованными в коротком плече хромосомы 5 [13–15]. Локусы Hrd A и Hrd B являются высокополиморфными (по локусу Hrd A у культурного ячменя выявлено более 150 аллелей, по локусу Hrd B – более 250 аллелей). Аллели этих локусов кодируют группы совместно наследуемых компонентов гордеинов (блоки компонентов), различающиеся по числу, подвижности и интенсивности входящих в них белковых полос [13, 16]. В силу своего уникального полиморфизма гордеин-кодирующие локусы по эффективности в оценке изменчивости генетических ресурсов не уступают некоторым типам молекулярных маркеров [17]. Ранее нами при сравнительном анализе полиморфизма запасных белков у культурного и дикого ячменя из Израиля, Ирана и Юго-Восточной Турции были получены данные, не позволяющие рассматривать эти территории в качестве центра или центров доместикации ячменя [18–20].
В настоящем сообщении представлены результаты сравнительного анализа аллельного разнообразия локусов Hrd A и Hrd В в образцах культурного и дикого ячменя из северо-западной части Иордании к востоку от р. Иордан, входившей в состав Палестины как части Дуги Плодородия.
МАТЕРИАЛЫ И МЕТОДЫ
Гордеины изучали в 34 местных образцах H. vulgare и в 19 образцах H. spontaneum из северных и западных районов Иордании, которые были любезно предоставлены доктором Яном Валькуном (J. Valkoun) из International Center for Agricultural Research in the Dry Areas (Aleppo, Syria), а также доктором Бенджамином Килианом (B. Kilian) из IPK (Gatersleben, Germany). Для всех образцов известны географические координаты сборов (рис. 1, табл. 1). Электрофорез гордеинов проводили в столбиках 12–14%-ного крахмального геля с 3 М мочевиной в алюминий-лактатном буфере с рН 3.1 по методике А.А. Созинова и Ф.А. Поперели [21] с некоторыми модификациями [16]. От каждого образца культурного и дикого ячменя анализировали гордеины в 10 и 5 индивидуальных зерновках соответственно. Варианты блоков компонентов, контролируемые аллелями локусов Hrd A и Hrd B, и генетические формулы гордеина определяли, пользуясь каталогами, составленными нами ранее в результате анализа более 1600 местных и коммерческих сортов ячменя из 25 стран мира [16]. Аллели этих локусов и соответствующие им варианты блоков компонентов гордеинов А и В, которые нами были обнаружены исключительно у H. spontaneum, обозначали цифрами с индексом “s”.
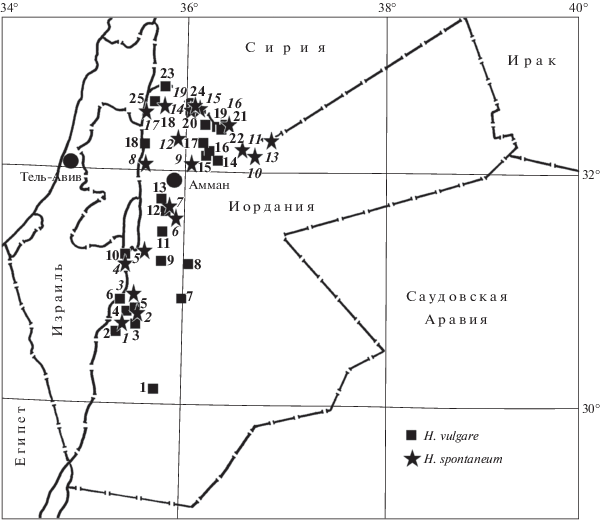
Таблица 1.
Генетические формулы гордеина и координаты мест сбора образцов культурного и дикого ячменя в Иордании
| № п/п | № каталога |
Формула гордеина по локусам | Место сбора образцов | ||
|---|---|---|---|---|---|
| Hrd A | Hrd B | сев. широта | вост. долгота | ||
| H. vulgare | |||||
| 1 | IG 31530 | 33 + 127 | 5 + 65 + 70 + 260 | 30°11′ | 35°50′ (1) |
| 2 | IG 31531 | 33 + 74 + 97 | 18 + 203 | 30°30′ | 35°30′ (2) |
| 3 | IG 31532 | 33 + 114 + 127 | 5 + 70 + 71 + 259 | То же | |
| 4 | IG 31533 | 33 | 79 + 260 | » | |
| 5 | IG 31534 | 13 + 33 | 70 + 79 + 259 + 260 | 30°37′ | 35°40′ (3) |
| 6 | IG 36044 | 33 + 127 | 65 + 70 + 203 + 264 | 30°46′ | 35°32′ (4) |
| 7 | IG 31536 | 5 + 33 + 114 + 127 | 5 + 65 + 70 + 260 + + 263 + 264 + 265 |
30°47′ | 35°40′ (5) |
| 8 | IG 31537 | 33 + 101 + 114 + 127 + 145 | 65 + 71 + 251 + 260 + 263 | То же | |
| 9 | IG 36039 | 33 + 127 | 65 + 70 + 203 + 263 | 30°51′ | 35°32′ (6) |
| 10 | IG 31404 | 1 | 6 + 8 | 30°51′ | 36°00′ (7) |
| 11 | IG 31462 | 20 + 33 + 127 | 5 + 65 + 70 | 31°14′ | 36°03′ (8) |
| 12 | IG 31402 | 1 | 6 | 31°15′ | 35°46′ (9) |
| 13 | IG 31403 | 1 + 3 | 8 + 155 | То же | |
| 14 | IG 31459 | 4 + 33 + 127 | 65 + 70 + 162 + 203 | 31°18′ | 35°38′ (10) |
| 15 | IG 36022 | 24 + 33 + 127 | 8 + 65 + 70 + 83 + 203 | 31°28′ | 35°46′ (11) |
| 16 | IG 36023 | 33 + 127 | 65 + 70 + 203 | То же | |
| 17 | IG 31401 | 1 + 3 | 10 + 15 | 31°41′ | 35°50′ (12) |
| 18 | IG 36043 | 33 + 133 | 65 | 31°47′ | 36°48′ (13) |
| 19 | IG 36019 | 24 + 33 + 127 | 66 + 70 + 162 + 183 + 203 + 263 | 32°03′ | 36°22′ (14) |
| 20 | IG 35223 | 2 + 33 + 146 | 71 + 87 + 263 | 32°06′ | 36°13′ (15) |
| 21 | IG 36016 | 127 | 70 | 32°07′ | 36°16′ (16) |
| 22 | IG 36018 | 20 + 33 + 133 | 65 | То же | |
| 23 | IG 35226 | 2 + 4 + 5 + 9 + 12 + 20 + + 100 + 141 | 65 + 251 + 254 + 257 + 264 | 32°10′ | 36°10′ (17) |
| 24 | IG 36000 | 2 + 32 | 14 + 65 | 32°10′ | 35°35′ (18) |
| 25 | IG 35224 | 5 + 22 + 33 + 114 + 127 | 2 + 65 + 254 + 256 | 32°17′ | 36°15′ (19) |
| 26 | IG 35225 | 13 + 14 | 65 + 263 | 32°18′ | 36°10′ (20) |
| 27 | IG 31553 | 33 + 127 | 70 + 141 | 32°21′ | 36°12′ (21) |
| 28 | IG 31549 | 2 | 1 + 14 + 61 + 130 + + 161 + 216 |
32°25′ | 36°10′ (22) |
| 29 | IG 31551 | 33 + 127 | 65 + 70 | То же | |
| 30 | IG 31541 | 2 + 28 + 65 + 106 | 12 + 17 + 65 + 208 | 32°29′ | 35°54′ (23) |
| 31 | IG 31546 | 2 + 62 + 68 + 76 | 1 + 14 + 24 + 142 | 32°32′ | 36°04′ (24) |
| 32 | IG 31548 | 2 + 20 + 136 + 141 | 2 + 65 + 79 + 206 257 |
То же | |
| 33 | IG 31544 | 2 + 23 + 28 + 106 | 8 + 12 + 17 + 19 + 29 + 65 | 32°32′ | 35°47′ (25) |
| 34 | IG 31545 | 2 + 23 | 8 + 19 + 29 | То же | |
| H. spontaneum | |||||
| 1 | IG 40064 | 62 | 236s | 30°35′ | 35°34′ (1) |
| 2 | IG 40063 | 77s + 82s | 79s + 247s + 259s | 30°42′ | 35°42′ (2) |
| 3 | IG 40055 | 59s + 62s | 9s + 238s | 30°53′ | 35°40′ (3) |
| 4 | IG 40045 | 81s + 82s | 9s + 239s | 31°11′ | 35°37′ (4) |
| 5 | IG 40040 | 32s + 83s | 141 + 234s | 31°17′ | 35°45′ (5) |
| 6 | IG 39821 | 22 + 33 + 82s | 101s + 243s + 253s | 31°42′ | 35°57′ (6) |
| 7 | IG 39828 | 62 + 53s | 239s + 251s | 31°47′ | 35°54′ (7) |
| 8 | IG 40014 | 41s + 81s | 19 + 242s + 252s | 32°01′ | 35°44′ (8) |
| 9 | IG 115780 | 1s + 63s + 75s | 65 + 253s | 32°01′ | 36°01′ (9) |
| 10 | IG 40035 | 33 | 101s | 32°03′ | 36°22′ (10) |
| 11 | IG 40034 | 33 + 86s | 243s + 245s | 32°07′ | 36°16′ (11) |
| 12 | IG 40012 | 1s + 25s | 257s + 258s | 32°10′ | 35°55′ (12) |
| 13 | IG 115782 | 22 + 32s | 101s + 235s | 32°15′ | 36°45′ (13) |
| 14 | IG 39850 | 45s + 60s | 248s + 249s | 32°19′ | 36°01′ (14) |
| 15 | IG 39827 | 32s + 74s + 78s | 241s + 246s | 32°20′ | 36°05′ (15) |
| 16 | IG 115795 | 22 + 97 + 53s + 78s | 235s + 244s + 254s | 32°20′ | 36°32′ (16) |
| 17 | IG 40002 | 1 + 24 | 240s + 250s | 32°29′ | 35°39′ (17) |
| 18 | IG 39886 | 62 + 30s | 256s + 265 | 32°32′ | 35°50′ (18) |
| 19 | IG 39825 | 78s + 82s | 141 + 90s + 234s | 32°32′ | 36°04′ (19) |
Примечание. В скобках указаны номера мест сбора на рис. 1; символом “s” обозначены аллели, обнаруженные исключительно у H. spontaneum.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
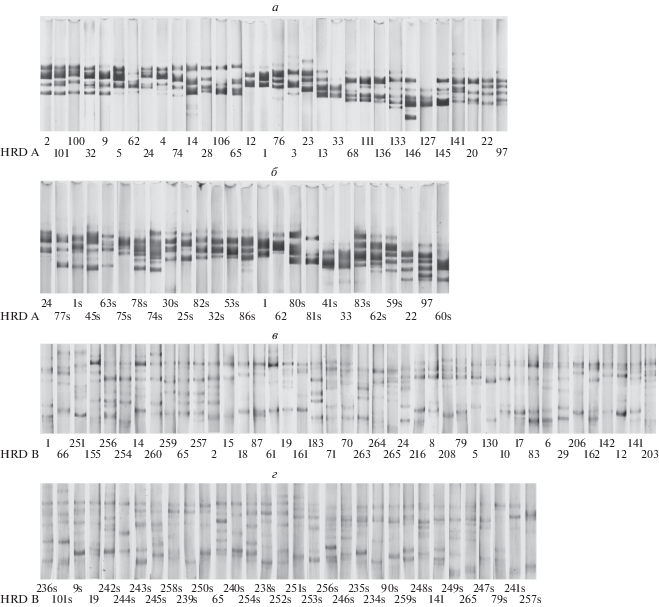
При изучении полиморфизма гордеинов методом электрофореза в 34 местных образцах культурного ячменя из Иордании было обнаружено 32 аллеля локуса Hrd A и 42 аллеля локуса Hrd B (табл. 1, рис. 2,а, в). В результате электрофоретического анализа гордеинов в 19 образцах H. spontaneum из Иордании было выявлено 26 аллелей локуса Hrd A и 32 аллеля локуса Hrd B (рис. 2,б, г).
Рис. 2.
Варианты блоков компонентов гордеинов, контролируемые аллелями локусов Hrd A и Hrd B в культурном (a, в) и диком (б, г) ячмене из Иордании. Символом “s” обозначены блоки компонентов гордеинов, обнаруженные исключительно у H. spontaneum.
Обнаруженные в образцах культурного ячменя из Иордании аллели локусов Hrd A и Hrd B значительно различались по частотам (табл. 2). Среди аллелей локуса Hrd A по частоте выделялись аллели А33 (0.2707), А127 (0.2412), А2 (0.1382) и А1 (0.0911). Частоты остальных аллелей варьировали в пределах 0.0029–0.0266. Из 26 аллелей этого локуса, присутствовавших в образцах H. spontaneum, шесть аллелей (А1, А22, А24, А33, А62, А97) ранее были идентифицированы и у культурного ячменя из мировой коллекции [16]. Эти аллели были обнаружены и в местных иорданских образцах культурного ячменя Иордании (табл. 2). Остальные 20 аллелей этого локуса, выявленные в образцах H. spontaneum, в образцах культурного ячменя из мировой коллекции не встречались. Среди аллелей локуса Hrd A у дикого ячменя из Иордании наибольшую частоту имели аллели А82s (0.1263), A22 (0.1053) и А62 (0.0947). Сравнительно высокой частотой характеризовались аллели А32s (0.0632), а также А41s, A53s и A77s (0.0526). Частоты остальных аллелей этого локуса варьировали в пределах 0.0105–0.0421.
Таблица 2.
Частоты аллелей локусов Hrd A и Hrd B в образцах культурного (H. vulgare) и дикого (H. spontaneum) ячменя из Иодании
| Аллели Hrd A | H. vulgare | H. spontaneum | Аллели Hrd A | H. vulgare | H. spontaneum |
| 1 | 0.0911 ± 0.001 | 0.0421 ± 0.014 | 127 | 0.2412 ± 0.016 | 0.0 |
| 2 | 0.1382 ± 0.002 | 0.0 | 133 | 0.0176 ± 0.005 | 0.0 |
| 3 | 0.0266 ± 0.006 | 0.0 | 136 | 0.0029 ± 0.002 | 0.0 |
| 4 | 0.0059 ± 0.002 | 0.0 | 141 | 0.0059 ± 0.002 | 0.0 |
| 5 | 0.0118 ± 0.004 | 0.0 | 145 | 0.0029 ± 0.002 | 0.0 |
| 9 | 0.0029 ± 0.002 | 0.0 | 146 | 0.0088 ± 0.004 | 0.0 |
| 12 | 0.0029 ± 0.002 | 0.0 | 1s | 0.0 | 0.0105 ± 0.007 |
| 13 | 0.0147 ± 0.005 | 0.0 | 25s | 0.0 | 0.0316 ± 0.012 |
| 14 | 0.0176 ± 0.005 | 0.0 | 30s | 0.0 | 0.0211 ± 0.010 |
| 20 | 0.0147 ± 0.005 | 0.0 | 32s | 0.0 | 0.0632 ± 0.017 |
| 22 | 0.0029 ± 0.002 | 0.1053 ± 0.022 | 41s | 0.0 | 0.0526 ± 0.016 |
| 23 | 0.0266 ± 0.006 | 0.0 | 45s | 0.0 | 0.0105 ± 0.007 |
| 24 | 0.0088 ± 0.004 | 0.0105 ± 0.007 | 53s | 0.0 | 0.0526 ± 0.016 |
| 28 | 0.0118 ± 0.004 | 0.0 | 59s | 0.0 | 0.0421 ± 0.014 |
| 32 | 0.0118 ± 0.004 | 0.0 | 60s | 0.0 | 0.0421 ± 0.014 |
| 33 | 0.2707 ± 0.02 | 0.0105 ± 0.007 | 62s | 0.0 | 0.0105 ± 0.007 |
| 62 | 0.0029 ± 0.002 | 0.0947 ± 0.021 | 63s | 0.0 | 0.0316 ± 0.012 |
| 65 | 0.0059 ± 0.002 | 0.0 | 74s | 0.0 | 0.0105 ± 0.007 |
| 68 | 0.0059 ± 0.002 | 0.0 | 75s | 0.0 | 0.0105 ± 0.007 |
| 74 | 0.0088 ± 0.004 | 0.0 | 77s | 0.0 | 0.0526 ± 0.016 |
| 76 | 0.0059 ± 0.002 | 0.0 | 78s | 0.0 | 0.0421 ± 0.014 |
| 97 | 0.0059 ± 0.002 | 0.0211 ± 0.010 | 80s | 0.0 | 0.0105 ± 0.007 |
| 100 | 0.0029 ± 0.002 | 0.0 | 81s | 0.0 | 0.0105 ± 0.007 |
| 101 | 0.0029 ± 0.002 | 0.0 | 82s | 0.0 | 0.1263 ± 0.024 |
| 106 | 0.0059 ± 0.002 | 0.0 | 83s | 0.0 | 0.0421 ± 0.014 |
| 114 | 0.0147 ± 0.005 | 0.0 | 86s | 0.0 | 0.0421 ± 0.014 |
| Аллели Hrd B |
H. vulgare | H. spontaneum | Аллели Hrd B |
H. vulgare | H. spontaneum |
| 1 | 0.0088 ± 0.004 | 0.0 | 256 | 0.0118 ± 0.004 | 0.0 |
| 2 | 0.0088 ± 0.004 | 0.0 | 257 | 0.0235 ± 0.006 | 0.0 |
| 5 | 0.0441 ± 0.008 | 0.0 | 259 | 0.0059 ± 0.003 | 0.0 |
| 6 | 0.0324 ± 0.007 | 0.0 | 260 | 0.0765 ± 0.011 | 0.0 |
| 8 | 0.0618 ± 0.009 | 0.0 | 263 | 0.0471 ± 0.008 | 0.0 |
| 10 | 0.0235 ± 0.006 | 0.0 | 264 | 0.0176 ± 0.005 | 0.0 |
| 12 | 0.0118 ± 0.004 | 0.0 | 265 | 0.0029 ± 0.002 | 0.0105 ± 0.007 |
| 14 | 0.0235 ± 0.006 | 0.0 | 9s | 0.0 | 0.0526 ± 0.016 |
| 15 | 0.0059 ± 0.002 | 0.0 | 79s | 0.0 | 0.0105 ± 0.007 |
| 17 | 0.0059 ± 0.002 | 0.0 | 90s | 0.0 | 0.0105 ± 0.007 |
| 18 | 0.0088 ± 0.004 | 0.0 | 101s | 0.0 | 0.0947 ± 0.021 |
| 19 | 0.0118 ± 0.004 | 0.0105 ± 0.007 | 234s | 0.0 | 0.0526 ± 0.016 |
| 24 | 0.0176 ± 0.005 | 0.0 | 235s | 0.0 | 0.0737 ± 0.019 |
| 29 | 0.0266 ± 0.006 | 0.0 | 236s | 0.0 | 0.0526 ± 0.016 |
| 61 | 0.0029 ± 0.002 | 0.0 | 238s | 0.0 | 0.0421 ± 0.014 |
| Аллели Hrd B |
H. vulgare | H. spontaneum | Аллели Hrd B |
H. vulgare | H. spontaneum |
| 65 | 0.1824 ± 0.015 | 0.0105 ± 0.007 | 239s | 0.0 | 0.0526 ± 0.016 |
| 66 | 0.0029 ± 0.002 | 0.0 | 240s | 0.0 | 0.0105 ± 0.007 |
| 70 | 0.1765 ± 0.015 | 0.0 | 241s | 0.0 | 0.0211 ± 0.010 |
| 71 | 0.0147 ± 0.005 | 0.0 | 242s | 0.0 | 0.0105 ± 0.007 |
| 79 | 0.0088 ± 0.004 | 0.0 | 243s | 0.0 | 0.0211 ± 0.010 |
| 83 | 0.0029 ± 0.002 | 0.0 | 244s | 0.0 | 0.0105 ± 0.007 |
| 87 | 0.0088 ± 0.004 | 0.0 | 245s | 0.0 | 0.0421 ± 0.014 |
| 130 | 0.0059 ± 0.003 | 0.0 | 246s | 0.0 | 0.0316 ± 0.012 |
| 141 | 0.0176 ± 0.005 | 0.0421 ± 0.014 | 247s | 0.0 | 0.0211 ± 0.010 |
| 142 | 0.0029 ± 0.002 | 0.0 | 248s | 0.0 | 0.0105 ± 0.007 |
| 155 | 0.0029 ± 0.002 | 0.0 | 249s | 0.0 | 0.0421 ± 0.014 |
| 161 | 0.0118 ± 0.004 | 0.0 | 250s | 0.0 | 0.0421 ± 0.014 |
| 162 | 0.0118 ± 0.004 | 0.0 | 251s | 0.0 | 0.0105 ± 0.007 |
| 183 | 0.0029 ± 0.002 | 0.0 | 252s | 0.0 | 0.0316 ± 0.012 |
| 203 | 0.0413 ± 0.008 | 0.0 | 253s | 0.0 | 0.0421 ± 0.014 |
| 206 | 0.0029 ± 0.002 | 0.0 | 254s | 0.0 | 0.0211 ± 0.010 |
| 208 | 0.0059 ± 0.003 | 0.0 | 256s | 0.0 | 0.0421 ± 0.014 |
| 216 | 0.0029 ± 0.002 | 0.0 | 257s | 0.0 | 0.0316 ± 0.012 |
| 251 | 0.0059 ± 0.003 | 0.0 | 258s | 0.0 | 0.0211 ± 0.010 |
| 254 | 0.0088 ± 0.004 | 0.0 | 259s | 0.0 | 0.0211 ± 0.010 |
Из 42 аллелей локуса Hrd В, выявленных в образцах культурного ячменя Иордании, самую высокую частоту имели аллели B65 (0.1824) и B70 (0.1765). С относительно высокой частотой встречались аллели В260 (0.0765), В8 (0.0618), В263 (0.0471) и В203 (0.0413). Частоты остальных 36 аллелей находились в пределах 0.0029–0.0266. Среди 32 аллелей локуса Hrd B, идентифицированных в H. spontaneum из Иордании, три аллеля (В19, В65 и В141) были выявлены как в образцах культурного ячменя из мировой коллекции, так и в иорданских образцах. При этом аллель В265 к настоящему времени был найден в культурном ячмене только в Иордании. Среди аллелей этого локуса в исследованных образцах дикого ячменя наибольшую частоту имели аллели В101s (0.0947), В235s (0.0737), а также аллели B9s, B234s, B236s, B239s, присутствовавшие у изученных образцов с частотой 0.0526. Частоты остальных аллелей этого локуса варьировали в пределах 0.0105–0.0421.
Рассматривая Иорданию как возможное место введения ячменя в культуру и исследуя здесь полиморфизм гордеинов в H. vulgare и H. spontaneum, логично было ожидать присутствие в образцах культурного и дикого ячменя одних и тех же аллелей локусов Hrd А и Hrd В, а также присутствие в диком ячмене самых широко распространенных аллелей этих локусов не только в местных образцах культурного ячменя в Сирии, но и в других странах мира. Из 52 аллелей локуса Hrd A, идентифицированных у образцов культурного и дикого ячменя из Иордании, только шесть аллелей – А1, А22, А24, А33, А62 и А97 оказались общими. Их частоты в H. vulgare варьировали в пределах 0.0029 (А22, А62)–0.2707 (А33) и у H. spontaneum – в пределах 0.0105 (А24, А33)–0.1053 (А22). Из 70 аллелей локуса Hrd B, обнаруженных в образцах дикого и культурного ячменя из Иордании общих аллелей оказалось всего четыре – В19, В65, В141 и В265. При этом частоты аллеля В19 в культурном и диком ячмене были близкими – 0.0118 и 0.0105 соответственно. Частоты остальных аллелей в культурном и диком ячмене существенно различались. Так, частота аллеля В65 в H. vulgare составила 0.1824, а в H. spontaneum – 0.0105. Частоты аллелей В141 и В265 в культурном ячмене Иордании были 0.0176 и 0.0029 соответственно, а в диком ячмене – 0.0421 и 0.0105 соответственно.
В табл. 3 приведены частоты “культурных” аллелей локуса Hrd A и Hrd B, обнаруженных в образцах H. spontaneum из Иордании и в исследованных нами ранее образцах местных культурных ячменей из 21 страны мира, входящих в центры разнообразия ячменя и сопредельных с ними стран. Из таблицы видно, что каждый из аллелей А24 и А62 был обнаружен в местном культурном ячмене 14 стран с различными частотами. Так, аллель А24 с максимальной частотой 0.4572 присутствовал в эфиопских образцах ячменя. С относительно высокими частотами этот аллель был обнаружен в образцах культурного ячменя из Йемена (0.1180), Палестины (0.1000) и Марокко (0.0889). В образцах из других стран, где этот аллель был идентифицирован, его частота варьировала в пределах 0.0032 (Иран)–0.0527 (Турция). Аллель А62 с максимальной частотой встречался в образцах культурного ячменя из Палестины (0.1250). Относительно высокая частота этого аллеля (0.0667) отмечена в Марокко. В образцах H. vulgare из остальных 12 стран частота аллеля А62 варьировала в пределах 0.0017 (Эфиопия)–0.0386 (Египет). Аллели А1 и А33 с различными частотами были обнаружены в образцах местного культурного ячменя из 10 стан. Максимальные частоты аллеля А1 отмечены в Сирии (0.1400) и Иране (0.0730). В образцах культурного ячменя из остальных восьми стран частота этого аллеля варьировала от 0.0020 (Туркмения) до 0.0200 (Израиль). Аллель А33 с наибольшими частотами встречался в образцах H. vulgare из Сирии (0.2250), Марокко (0.1222) и Эфиопии (0.0718). В образцах культурного ячменя из остальных семи стран его частота находилась в пределах 0.0011 (Турция)–0.0438 (Йемен). Аллели А22 и А97 не имеют широкого распространения и были обнаружены в культурном ячмене отдельных стран с невысокими частотами. Среди “культурных” аллелей локуса Hrd B, идентифицированных в H. spontaneum из Иордании, наиболее распространенным в H. vulgare является аллель В19, обнаруженный с различными частотами в исследованных образцах культурного ячменя из 15 стран (табл. 3). Наибольшая частота этого аллеля отмечена в образцах из Киргизии (0.2127), Палестины (0.1000), Алжира (0.0910), Туркмении (0.0882), Турции (0.0753) и Афганистана (0.0738). В образцах H. vulgare из остальных девяти стран этот аллель встречался с частотами от 0.0030 (Узбекистан) до 0.0400 (Израиль). Аллель В65 был идентифицирован в образцах культурного ячменя только из пяти стран. Он с высокой частотой присутствовал в ячмене из Сирии (0.2182) и Ирака (0.0706), а в образцах из Узбекистана, Индии и Турции его частота варьировала в пределах 0.0011–0.0213. Аллель В141 не имеет широкого распространения и был найден в образцах H. vulgare только из трех стран, где его частота была в пределах от 0.0133 (Сирия) до 0.0539 (Йемен). Аллель В265, как отмечалось выше, был обнаружен только в образцах культурного ячменя из Иордании. Следует заметить, что некоторые “культурные” аллели локусов Hrd A и Hrd B, обнаруженные в H. spontaneum из Иордании, были нами обнаружены в образцах дикого ячменя из других стран. Так, аллель А1 с частотой 0.0349 встречался в исследованных образцах H. spontaneum из Ирана, аллель А22 с частотой 0.0465 обнаружен в диком ячмене из Ирана, Турции (0.0630) и Сирии (0.0261) [19, 20, 29]. Аллель В19 с частотой 0.0089 присутствовал в исследованных образцах H. spontaneum Юго-Восточной Турции [20]. При этом аллели А24, А62, А97, В65, В141 и В265 были найдены одновременно в культурном и диком ячмене только в Иордании (см. табл. 2).
Таблица 3.
Частоты аллелей локусов Hrd A и Hrd B, обнаруженных у H. spontaneum из Иордании в образцах местных культурных ячменей из стран Северной и Северо-Восточной Африки, Ближнего Востока, Передней, Центральной и Восточной Азии
| Страна | Аллели локуса Hrd A | Аллели локуса Hrd B | Ссылка | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| А1 | А22 | А24 | А33 | А62 | А97 | В19 | В65 | В141 | В265 | ||
| Марокко | 0.0 | 0.0222 | 0.0889 | 0.1222 | 0.0667 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | [22] |
| Тунис | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | » |
| Алжир | 0.0 | 0.0226 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0910 | 0.0 | 0.0 | 0.0 | » |
| Египет | 0.0 | 0.0 | 0.0333 | 0.0 | 0.0386 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | » |
| Эфиопия | 0.0068 | 0.0 | 0.4572 | 0.0718 | 0.0017 | 0.0 | 0.0290 | 0.0 | 0.0 | 0.0 | [23] |
| Йемен | 0.0 | 0.0 | 0.1180 | 0.0438 | 0.0045 | 0.0 | 0.0146 | 0.0 | 0.0539 | 0.0 | [25] |
| Афганистан | 0.0095 | 0.0 | 0.0226 | 0.0309 | 0.0059 | 0.0024 | 0.0738 | 0.0 | 0.0 | 0.0 | [24] |
| Ирак | 0.0059 | 0.0 | 0.0088 | 0.0 | 0.0 | 0.0 | 0.0029 | 0.0706 | 0.0 | 0.0 | [26] |
| Сирия | 0.1400 | 0.0 | 0.0067 | 0.2250 | 0.0117 | 0.0050 | 0.0 | 0.2182 | 0.0133 | 0.0 | » |
| Палестина | 0.0 | 0.0 | 0.1000 | 0.0 | 0.1250 | 0.0 | 0.1000 | 0.0 | 0.0 | 0.0 | » |
| Израиль | 0.0200 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0400 | 0.0 | 0.0 | 0.0 | » |
| Иран | 0.0730 | 0.0159 | 0.0032 | 0.0190 | 0.0 | 0.0 | 0.0159 | 0.0 | 0.0 | 0.0 | [27] |
| Туркмения | 0.0020 | 0.0 | 0.0 | 0.0118 | 0.0059 | 0.0 | 0.0882 | 0.0 | 0.0 | 0.0 | » |
| Узбекистан | 0.0 | 0.0 | 0.0059 | 0.0049 | 0.0010 | 0.0 | 0.0030 | 0.0020 | 0.0 | 0.0 | » |
| Киргизия | 0.0170 | 0.0 | 0.0 | 0.0 | 0.0213 | 0.0 | 0.2127 | 0.0 | 0.0 | 0.0 | » |
| Таджикистан | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0241 | 0.0 | 0.0 | 0.0 | » |
| Китай | 0.0125 | 0.0125 | 0.0146 | 0.0 | 0.0021 | 0.0 | 0.0291 | 0.0 | 0.0416 | 0.0 | [28] |
| Непал | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | » |
| Пакистан | 0.0 | 0.0 | 0.0214 | 0.0143 | 0.0357 | 0.0 | 0.0143 | 0.0 | 0.0 | 0.0 | » |
| Индия | 0.0 | 0.0 | 0.0255 | 0.0 | 0.0383 | 0.0 | 0.0 | 0.0213 | 0.0 | 0.0 | » |
| Турция | 0.0022 | 0.0 | 0.0527 | 0.0011 | 0.0140 | 0.0 | 0.0753 | 0.0011 | 0.0 | 0.0 | [20] |
Таким образом, сравнительный анализ аллельного разнообразия двух высокополиморфных локусов Hrd A и Hrd B в образцах H. spontaneum и H. vulgare из Иордании показал, что 11.5% аллелей локуса Hrd A и 5.7% аллелей локуса Hrd B являются общими. При этом два из общих аллелей локуса Hrd A и один – локуса Hrd B были найдены в диком ячмене и из других стран. Заметим также, что наиболее древнее использование ячменя в Иордании по археологическим данным датируется 6700 г. до н. э. в Beidha, тогда как в Сирии – Tell Abu Hureyra, наиболее ранние свидетельства использования ячменя относятся к 9000 г. до н. э. [30]. Добавим к этому, что самое раннее использование зерна ячменя на Ближнем Востоке зафиксировано в Охало II около Галилейского моря и датируется 17 тыс. до н. э. [31]. Все это свидетельствует о том, что район Северо-Западной Иордании не является местом доместикации ячменя, но одновременно не исключает того, что H. spontaneum в Иордании мог являться донором некоторых аллелей гордеин-кодирующих локусов для H. vulgare в результате интрогрессии при спонтанной гибридизации.
Работа выполнена при поддержке программы Президиума РАН № 41 “Биоразнообразие природных систем и биологические ресурсы России”.
Список литературы
Provan J., Russell J.R., Booth A., Powell W. Polymorphic chloroplast simple sequence repeat primers for systematic and population studies in the genus Hordeum // Mol. Ecol. 1999. № 8. P. 505–511. doi 10.1046/j.1365-294X.1999.00545.x
Бахтеев Ф.Х. К истории культуры ячменя в СССР // Материалы по истории земледелия СССР: Сб. II. М.; Л. Изд-во АН СССР, 1956. С. 204–257.
Вавилов Н.И. Учение о происхождении культурных растений после Дарвина // Сов. наука. 1940. № 2. С. 55–75.
Åberg E. Hordeum agriocrithon nova sp. a wild six rowed barley // Ann. Roy. Agr. Col. Sweden. 1938. V. 6. P. 159–216.
Zhang Q., Saghai Maroof M.A., Yang P.G. Ribosomal DNA polymorphisms and the Oriental-Occidental genetic differentiation in cultivated barley // Theor. Appl. Genet. 1992. V. 84. № 5. P. 682–687. doi 10.1007/ BF00224168
Molino-Cano J.-L., Fra-Mon P., Salcedo G. et al. Marocco as possible domestication centre for barley: Biochemical and agromorphological evidence // Theor. Appl. Genet. 1987. V. 73. № 4. P. 531–536. doi 10.1007/BF00289190
Negassa M. Patterns of phenotypic diversity in an Ethiopian barley collection, and the Arussi-Bale Higland as a center of origin ob barley // Hereditas. 1985. V. 102. № 1. P. 139–150. doi 10.1111/j.1601-5223.1985.tb00474.x
Lev-Yadum S., Gopher A., Abbo S. The cradle of agriculture // Science. 2000. V. 288. P. 1602–1603. doi 10.1126/science.288.5471.1602
Harlan J.R., Zohary D. Distribution of wild wheats and barley // Science. 1966. V. 153. № 3740. P. 1074–1080. doi 10.1126/science.153.3740.1074
Kilian B., Özkan H., Kohl J. et al. Haplotype structure at seven barley genes: relevance to gene pool bottlenecks, phylogeny of ear type and site of barley domestication // Mol. Gen. Genomics. 2006. V. 276. P. 230–241. doi 10.1007/s00438-006-0136-6
Morrell P.L., Clegg M.T. Genetic evidence for a second domestication of barley (Hordeum vulgare) east of the Fertile Crescent // Proc. Natl Acad. Sci. USA. 2007. V. 104. № 9. P. 3289–3294.
Allaby R.G. Barley domestication: the end of central dogma? // Genome Biol. 2015. V. 16. № 117. https://doi.org/10.1186/s13059-015-0743-9.
Созинов А.А., Нецветаев В.П., Григорян Э.М., Образцов И.С. Картирование локусов Hrd у ячменя (Hordeum vulgare L. emed. Vav. et Bach.) // Генетика. 1978. Т. 14. № 9. С. 1610–1619.
Поморцев А.А., Нецветаев В.П., Попереля Ф.А., Созинов А.А. Идентификация шестого локуса, контролирующего синтез гордеина у озимого ячменя // Докл. ВАСХНИЛ. 1983. № 1. С. 7–9.
Netsvetaev V.P., Sozinov A.A. Location of a hordein G locus, Hrd G, on chromosome 5 of barley // Barley Genetics Newsletter. 1984. V. 14. P. 4–6.
Поморцев А.А., Лялина Е.В. Оценка сортовой принадлежности и сортовой чистоты семян ячменя методом электрофоретического анализа запасных белков зерна // Методич. пособие к практикуму “Белковые маркеры для генетической паспортизации и улучшения геномов растений хозяйственно ценных видов”. М., 2011. 86 с.
Dakir El-H., Ruiz M.-L., Garcia P., de la Vega P. Genetic variability evaluation in Moroccan collection of barley, Hordeum vulgare L., by means of storage proteins and RAPDs // Genet. Res. and Crop Evol. 2002. V. 49. P. 619–631. doi 10.1023/A:1021228730714
Поморцев А.А., Лялина Е.В. Аллельное разнообразие гордеин-кодирующего локуса Hrd A у Hordeum spontaneum C. Koch – дикого родича культурного ячменя из Израиля (как части Дуги Плодородия) // Тр. по прикл. бот., генет. и селек. 2009. Т. 166. С. 450–457.
Поморцев А.А., Лялина Е.В. Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L.) и дикого (H. spontaneum C. Koch) ячменя в Иране (как части Дуги Плодородия) // Генетика. 2016. Т. 52. № 10. С. 1146–1158. doi 10.7868/S001667581610009X
Поморцев А.А., Болдырев С.В., Лялина Е.В. Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L.) и дикого (H. spontaneum C. Koch) ячменя в Турции (как части Дуги Плодородия) // Генетика. 2017. Т. 53. № 4. С. 449–459. doi 10.7868/S0016675817030092
Созинов А.А., Попереля Ф.А. Методика вертикального дискового электрофореза в крахмальном геле и генетический принцип классификации глиадинов. Одесса, 1978. 16 с.
Поморцев А.А., Калабушкин Б.А., Терентьева И.А. Полиморфизм гордеинов у ячменей Северной Африки // Генетика. 2002. Т. 38. № 11. С. 1498–1510.
Поморцев А.А. Исследование полиморфизма гордеинов у ячменей Эфиопии // Генетика. 2001. Т. 37. № 10. С. 1371–1382.
Поморцев А.А., Мартынов С.П., Ковалева О.Н., Лялина Е.В. Полиморфизм по гордеин-кодирующим локусам культурного ячменя (Hordeum vulgare L.) Афганитстана // Генетика. 2010. Т. 46. № 11. С. 1507–1515. doi 10.1134/S1022795410110086
Поморцев А.А., Лялина Е.В. Полиморфизм культурного ячменя (Hordeum vulgare L.) Южной Аравии по гордеин-кодирующим локусам // Генетика. 2007. Т. 43. № 5. С. 660–667. doi 10.1134/ S1022795407050092
Поморцев А.А., Мартынов С.П., Лялина Е.В. Полиморфизм гордеин-кодирующих локусов в местных популяциях культурного ячменя (Hordeum vulgare L.) стран Ближнего Востока // Генетика. 2008. Т. 44. № 6. С. 709–721. doi 10.1124/S1022795408060112
Поморцев А.А., Мартынов С.П., Ковалева О.Н., Лялина Е.В. Полиморфизм по гордеин-кодирующим локусам в популяциях ячменя (Hordeum vulgare L.) Ирана и стран Средней Азии // Генетика. 2011. Т. 47. № 11. С. 1545–1564. doi 10.1134/S1022795411110147
Поморцев А.А., Мартынов С.П., Лялина Е.В. Полиморфизм по гордеин-кодирующим локусам популяций ячменя (Hordeum vulgare L.) стран Восточной Азии (Китай, Непал, Пакистан, Индия) // Генетика. 2012. Т. 48. № 8. С. 934–950. doi 10.1134/ S1022795412050183
Поморцев А.А., Болдырев С.В., Лялина Е.В. Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L.) и дикого (H. spontaneum C. Koch) ячменя в Сирии (как части Дуги Плодородия) // Генетика. 2018. Т. 54. № 11. С. 1275–1284.
Zohary D., Hopf M. Domestication of Plants in the Old World. The Origin and Spread of Cultivated Plants in West Asia, Europe, and Nile Valley. Oxford: Clarendon Press, 1988. 245 p.
Willcox G. Archeobotanical evidence for the beginnings of agriculture in Southwest Asia // Proc. Harlan Symp. The Origins of Agriculture and Crop Domestication. Aleppo, 1997. P. 25–38.
Дополнительные материалы отсутствуют.