

Генетика, 2019, T. 55, № 4, стр. 426-438
Гибридная зона между хромосомными расами Сок и Серов у обыкновенной бурозубки Sorex araneus (Lipotyphla, Mammalia) на территории Европейской России
Н. А. Щипанов 1, А. А. Калинин 1, А. В. Бобрецов 2, 3, **, С. В. Павлова 1, *
1 Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
119071 Москва, Россия
2 Печоро-Илычский государственный природный заповедник
169436 Республика Коми, пос. Якша, Россия
3 Институт биологии Коми научного центра Уральского отделения
Российской академии наук
167982 Сыктывкар, Россия
** E-mail: avbobr@mail.ru
* E-mail: swpavlova@mail.ru
Поступила в редакцию 07.05.2018
После доработки 01.07.2018
Принята к публикации 07.08.2018
Аннотация
Анализ распределения частот кариотипических вариантов в гибридной зоне между хромосомными расами Сок и Серов у обыкновенной бурозубки Sorex araneus показал, что зона имеет сложную конфигурацию. Значительная часть зоны контакта расположена на участке однотипной средней тайги в отсутствие каких-либо барьеров, препятствующих расселению зверьков, и частично проходит вдоль р. Печора (Республика Коми). Анализ распределения диагностических хромосом на трансекте, ориентированном вдоль р. Печора, показывает, что конфигурацию гибридной зоны можно описать встречными (opposing) клинами. Особи обеих чистых рас Сок и Серов, а также межрасовые гибриды F1 обнаружены только на левом берегу р. Печора. На правом берегу реки были найдены только кариотипы расы Серов и единичные гибриды. Число межрасовых гибридов в центре зоны не отличается от ожидаемого при свободном скрещивании, что позволяет охарактеризовать зону как унимодальную. Стандартная ширина зоны составляет около 40 км, что значительно превышает этот показатель для известных случаев гибридизации рас с таким же уровнем различий в кариотипе (по двум парам диагностических метацентриков с монобрахиальной гомологией). Обсуждаются возможные причины этого явления.
По выражению Г. Хьюитта [1], гибридные зоны являются “природными лабораториями для эволюционных исследований”. Конфигурация и ширина гибридных зон, с одной стороны, позволяют судить о степени и факторах изоляции внутривидовых форм, с другой – увидеть возникающие вследствие гибридизации новые генетические варианты [2, 3]. Гибридные зоны между различными внутривидовыми кариотипическими формами наиболее интересны, так как в этом случае принадлежность к определенной форме однозначно маркирована кариотипом, а гибриды надежно определяются по сочетанию у особи вариантов хромосом обеих скрещивающихся форм.
Одной из наиболее удобных моделей для изучения гибридизации кариотипических форм является мелкое насекомоядное млекопитающее – обыкновенная бурозубка Sorex araneus. Кариотип этого вида представлен инвариантной частью и вариабельной, состоящей из 10 хромосомных плеч, которые могут оставаться в акроцентрическом состоянии или сливаться за счет робертсоновских (Rb) транслокаций в различные комбинации, образуя метацентрические хромосомы [4]. Дальнейшая эволюция полностью метацентрических кариотипов S. araneus возможна за счет полноплечевых реципрокных транслокаций (WART), при этом оба типа перестроек рассматривают как “один эволюционный шаг”, так как для образования нового варианта кариотипа достаточна одна мутация Rb или WART [5]. Все хромосомы в кариотипе, ранжированные по размеру, принято обозначать строчными буквами латинского алфавита, начиная с а [6].
Акроцентрические и метацентрические хромосомы вариабельной части кариотипа называются “диагностическими” и определяют диагноз различных кариотипических форм S. araneus. Эти формы распределены парапатрично и называются “хромосомные расы” – группы популяций с одинаковым набором акроцентрических и метацентрических хромосом, передаваемым по наследству, и занимающие общее неразделенное пространство [7]. Всего к настоящему времени на ареале вида известно более 75 хромосомных рас и их число продолжает увеличиваться [8–10]. В местах контакта ареалов хромосомных рас образуются гибридные зоны. Изучение известных гибридных зон позволило показать, что их стандартная ширина находится в обратной зависимости от количества хромосом, различающих расы, и, соответственно, от сложности конфигурации, возникающей в мейозе у гибридов [11]. Большинство известных гибридных зон S. araneus характеризуются дефицитом гибридов, т.е. их можно рассматривать как “бимодальные”. Такие зоны позволяют предполагать, что эти формы встают на путь видообразования [12]. На данный момент известно около 35 мест межрасовой гибридизации на всем ареале вида от Британских островов до Байкала и не менее 14 из них локализованы на территории России. Однако данные о стандартной ширине российских гибридных зон получены лишь в двух случаях: между расами Москва–Селигер и Кириллов–Печора [11, 13], и еще в двух случаях подробно описано распределение гибридов в зоне контакта [14–16]. Таким образом, изученность хромосомных гибридных зон у обыкновенной бурозубки на территории России нельзя назвать исчерпывающей.
Одним из интересных аспектов формирования гибридных зон является их “притягивание” в места с пониженной плотностью особей – попадание в “ловушки низкой плотности” [1]. Такие “ловушки” могут возникать во всех случаях, когда миграция между локальными популяциями ослаблена вследствие низкой общей численности, либо при наличии некоторого препятствия, затрудняющего миграцию. Следует заметить, что в большинстве известных случаев локализация гибридных зон не связана с какой-либо внешней преградой (см. обзор [4]). Анализ распределения хромосомных рас на российской части ареала показал [8], что в своем распространении расы не ограничены физическими барьерами, в том числе большими реками, не связаны с определенными экологическими или климатическими особенностями местности и в большинстве случаев в зоне предполагаемого контакта рас невозможно предположить ожидаемого физического барьера. Однако в том же исследовании показано, что если одна из рас занимает пространство пусть даже за сравнительно небольшим препятствием, то она сама по себе блокирует внедрение сюда чужой расы [8]. С одной стороны, наличие барьера заметно уменьшает ширину гибридной зоны [13, 17–19], а с другой, при определенной ориентации зоны к препятствию, например, если раса ограничена с трех сторон, может возникать эффект “затекания” расы на территорию другой расы – “карманный” эффект, который увеличивает стандартную ширину гибридной зоны [20].
Ранее, при изучении распределения хромосомных рас на севере европейской части России, мы указывали на то, что вероятным месторасположением зоны контакта между расами Сок и Серов может являться верхнее течение р. Печоры около впадения в нее р. Илыч (Республика Коми). Обнаружение межрасового гибрида в этом месте позволило предположить, что речное русло может внести свой вклад в формирование направления (курса) гибридной зоны [21]. К настоящему времени накоплено достаточное количество данных, позволяющее провести предварительный анализ этой гибридной зоны.
Целью данной публикации является описание локализации и курса гибридной зоны между хромосомными расами Сок и Серов на территории Республики Коми и первичный анализ распределения частот встречаемости особей чистых (родительских) рас и их гибридов в этой гибридной зоне.
МАТЕРИАЛЫ И МЕТОДЫ
Территория и населяющие ее расы. Распределение кариотипических вариантов S. araneus рассмотрено на участке между городами Ухта и Сыктывкар (Республика Коми) и далее на восток, до верховьев рек Илыч и Печора с притоком р. Унья. Северные и горные области на этом участке представлены северно-таежными, южнее – среднетаежными еловыми лесами с преобладающим участием сибирской ели (Picea abovata), пихты (Abies sibirica) и кедра (Pinus sibirica). Для территории характерны значительные по площади сосняки на бедных песчаных почвах и обширные заболоченные участки [22]. Средняя ширина р. Печоры, измеренная троекратно на соответствующем отрезке по космическим фотографиям Google Earth, составляет около 90 м от впадения р. Уньи до пос. Якша, около 150 м ниже до устья р. Илыч и около 300 м ниже до пос. Вуктыл.
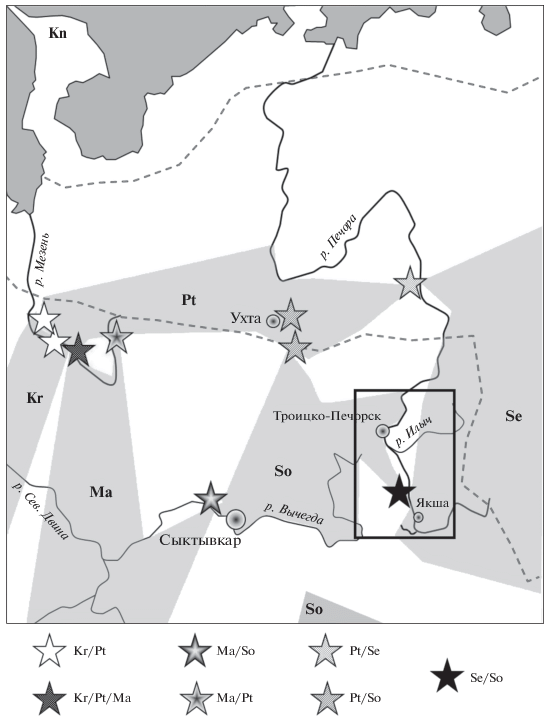
В рассматриваемом регионе известны шесть хромосомных рас. На севере распространена раса Печора, которая формирует зону гибридизации с расой Сок в районе г. Ухта [21]. Раса Серов пересекает Уральские горы с востока, спускается по западному макросклону и выходит на равнину к р. Печора [23]. Раса Сок обитает в районе среднего и южного Тимана [24], однако ее присутствие к югу от р. Вычегда до Кировской области пока не подтверждено кариотипированием [21]. Далее на юг раса Сок известна в Заволжье, где доходит до лесных островов в саратовской степи [25]. На западе региона распространена раса Мантурово, которая занимает обширный ареал и формирует зоны контакта и гибридизации с расой Сок на р. Вычегда, а с расой Печора на р. Мезень [26]. Ареалы рас Канин и Кириллов отделены от рас Сок и Серов ареалом расы Печора (рис. 1).
Рис. 1.
Распространение хромосомных рас в исследуемом регионе (по [21] с учетом новых данных табл. 2). Известные ареалы рас обозначены серым цветом, территория с неизвестной расовой принадлежностью S. araneus – белым. В рамке – территория, представленная на рис. 3. Места обнаружения межрасовых гибридов F1 обозначены звездочками. Пунктиром отмечены границы северной тайги. Обозначения рас: So – Сок, Se – Серов, Ма – Мантурово, Kr – Кириллов, Pt – Печора, Kn – Канин.
Для расчета клины диагностических хромосом использованы данные распределения кариотипических вариантов на участке от окрестностей г. Ухты до устья левого притока Печоры – р. Волосница. Выборки взяты и вдоль трассы Ухта– Троицко-Печорск, далее вдоль дороги на пос. Якша, по местным проезжим дорогам и вдоль р. Печора по доступным притокам. Локализация мест выборки определялась только их доступностью.
Стандартную ширину клины определяли при подгонке кривой tanh [27]:
где y – частота встречаемости диагностических хромосом в выборке, х – расстояние от начала трансекта (Волосница), В – координата центра, А – коэффициент максимального наклона кривой. Ширина клины получена как удвоенный обратный коэффициент A [28].Выборки и кариотипирование. В анализе использованы опубликованные ранее (табл. 1, 2005–2010 гг.) и новые данные (табл. 2, 2011–2017 гг.) о распределении хромосомных рас на исследуемом участке. Всего проанализировано 146 кариотипированных особей из 59 локалитетов, из них данные по распределению 92 особей в 35 локалитетах приводятся впервые (табл. 2).
Таблица 1.
Распределение кариотипических вариантов Sorex araneus на изучаемой территории
| № п.п. | Локалитет | Координаты | Кариотип | Se | F1 | So | |
|---|---|---|---|---|---|---|---|
| с.ш. | в.д. | ||||||
| 1 | с. Дутово, 147 км восточнее г. Ухта (лев. берег р. Печора, переправа на Вуктыл) | 63°41′ | 56°39′ | Pt-Se | 0 | 1 | 0 |
| 2 | г. Ухта | 63°33′ | 53°48′ | Pt-So im | 0 | 2 | 0 |
| 3 | с. Седью | 63°27′ | 53°57′ | Pt-So | 0 | 1 | 0 |
| 4 | пос. Комсомольск-на-Печоре, 15 км к северу | 62°17′ | 56°36′ | So-Se | 0 | 1 | 2 |
| 5 | р. Чомкосаель | 62°44′ | 54°38′ | So | 0 | 0 | 2 |
| 6 | пос. Верхняя Омра | 62°48′ | 55°25′ | So | 0 | 0 | 4 |
| 7 | пос. Троицко-Печорск | 62°42′ | 56°11′ | So | 0 | 0 | 2 |
| 8 | с. Пузла | 62°26′ | 54°39′ | So | 0 | 0 | 6 |
| 9 | г. Эжва (лев. берег р. Вычегда) | 61°47′ | 50°44′ | Ma-So | 0 | 1 | 0 |
| 10 | г. Ухта, 42 км к югу | 63°14′ | 54°21′ | Se | 2 | 0 | 0 |
| 11 | р. Березовка | 61°41′ | 56°31′ | Se | 2 | 0 | 0 |
| 12 | р. Нем | 61°40′ | 56°14′ | Se | 2 | 0 | 0 |
| 13 | р. Волосница | 61°42′ | 56°44′ | Se | 1 | 0 | 0 |
| 14 | пос. Якша | 61°49′ | 56°50′ | Se | 1 | 0 | 0 |
| 15 | Верховья р. Печора | 62°03′ | 58°25′ | Se | 4 | 0 | 0 |
| 16 | Верховья р. Печора | 62°05′ | 59°05′ | Se | 1 | 0 | 0 |
| 17 | Верховья р. Илыч | 63°03′ | 58°36′ | Se | 1 | 0 | 0 |
| 18 | р. Илыч | 62°38′ | 58°55′ | Se | 1 | 0 | 0 |
| 19 | р. Илыч | 62°28′ | 58°58′ | Se | 1 | 0 | 0 |
| 20 | р. Ропча | 63°00′ | 52°19′ | So | 0 | 0 | 2 |
| 21 | пос. Малая Слуда (прав. берег р. Вычегда) | 62°00′ | 50°35′ | So | 0 | 0 | 3 |
| 22 | пос. Приозерный (прав. берег р. Вычегда) | 61°47′ | 51°49′ | So | 0 | 0 | 5 |
| 23 | пос. Сторожевск (прав. берег р. Вычегда) | 61°57′ | 52°20′ | So | 0 | 0 | 5 |
| 24 | пос. Усть-Нем (прав. берег р. Вычегда) | 61°40′ | 54°53′ | So | 0 | 0 | 1 |
Таблица 2.
Новые данные по распределению кариотипических вариантов Sorex araneus на изучаемой территории
| № п.п. |
Локалитет | Координаты | Se | F1 | So | |
|---|---|---|---|---|---|---|
| с.ш. | в.д. | |||||
| 25 | Правый берег р. Печора, 10 км ниже пос. Усть-Илыч | 62°35′16′′ | 56°43′14′′ | 2 | 0 | 0 |
| 26 | Правый берег р. Печора, 12 км ниже пос. Усть-Илыч | 62°36′07′′ | 56°40′06′′ | 3 | 0 | 0 |
| 27 | Правый берег р. Илыч, 16 км выше устья | 62°35′05′′ | 56°54′36′′ | 3 | 0 | 0 |
| 28 | Правый берег р. Илыч, напротив р. Чолыч | 62°34′08′′ | 57°04′33′′ | 4 | 0 | 0 |
| 29 | Правый берег р. Печора, 12 км выше пос. Усть-Илыч | 62°25′12′′ | 56°46′11′′ | 1 | 1 | 0 |
| 30 | Правый берег р. Печора, 18 км выше пос. Усть-Илыч | 62°22′05′′ | 56°45′48′′ | 2 | 0 | 0 |
| 31 | Правый берег р. Печора, у острова | 62°11′30′′ | 56°40′45′′ | 4 | 1 | 0 |
| 32, 33 | Остров на р. Печора | 62°11′43′′ | 56°40′11′′ | 1 | 1 | 0 |
| 34 | Правый берег р. Печора, устье р. Дозмер | 62°01′37′′ | 56°47′51′′ | 3 | 0 | 0 |
| 35 | Правый берег р. Печора, р. Дозмер 1 км | 62°01′28′′ | 56°48′57′′ | 4 | 0 | 0 |
| 36 | пос. Якша (правый берег р. Печора) | 61°49′19′′ | 56°51′54′′ | 1 | 0 | 0 |
| 37 | пос. Якша (левый берег р. Печора) | 61°48′59′′ | 56°50′15′′ | 3 | 0 | 0 |
| 38 | Левый берег р. Печора, 18 км ниже пос. Якши | 61°55′16′′ | 56°49′08′′ | 4 | 0 | 0 |
| 39 | Левый берег р. Печора, напротив р. Дозмер | 62°01′38′′ | 56°46′21′′ | 5 | 1 | 0 |
| 40 | Левый берег р. Печора, между дер. Мамыль и пос. Комсомольск-на-Печоре, 1.03 км от берега | 62°01′57′′ | 56°43′01′′ | 1 | 0 | 0 |
| 41 | Левый берег р. Печора, между дер. Мамыль и пос. Комсомольск-на-Печоре | 62°04′27′′ | 56°45′28′′ | 2 | 0 | 1 |
| 42 | Дорога между пос. Комсомольск-на-Печоре и пос. Якша | 62°05′04′′ | 56°38′38′′ | 2 | 0 | 0 |
| 43 | Дорога между пос. Комсомольск-на-Печоре и пос. Троицко-Печорск | 62°09′23′′ | 56°34′53′′ | 0 | 0 | 1 |
| 44 | Левый берег р. Печора, у острова | 62°11′42′′ | 56°39′39′′ | 2 | 0 | 0 |
| 45 | Левый берег р. Печора, 16 км ниже пос. Комсомольск-на-Печоре | 62°13′14′′ | 56°40′43′′ | 1 | 1 | 0 |
| 46 | Левый берег р. Печора, 23 км ниже пос. Комсомольск-на-Печоре | 62°15′56′′ | 56°45′35′′ | 2 | 0 | 0 |
| 47 | Левый берег р. Печора, 30 км ниже пос. Комсомольск-на-Печоре | 62°18′45′′ | 56°44′15′′ | 1 | 2 | 0 |
| 48, 53 | Левый берег р. Печора, 9 км выше пос. Усть-Илыч | 62°26′37′′ | 56°47′10′′ | 1 | 0 | 1 |
| 49 | пос. Белый Бор | 62°34′08′′ | 56°23′07′′ | 0 | 2 | 2 |
| 50 | Левый берег р. Печора, 10 км ниже пос. Усть-Илыч | 62°35′06′′ | 56°41′36′′ | 3 | 1 | 1 |
| 51 | Левый берег р. Печора, 15 км ниже пос. Усть-Илыч | 62°36′26′′ | 56°35′48′′ | 0 | 1 | 3 |
| 52 | Левый берег р. Печора, 50 км ниже пос. Усть-Илыч | 63°02′43′′ | 56°03′03′′ | 0 | 0 | 1 |
| 54 | р. Велью, 4 км от устья (левый берег р. Печора) | 63°13′17′′ | 55°52′06′′ | 0 | 0 | 2 |
| 55 | р. Безволосица | 62°05′30′′ | 56°34′45′′ | 1 | 0 | 0 |
| 56 | Верховья р. Илыч (правый берег) | 62°38′10′′ | 58°55′11′′ | 4 | 0 | 0 |
| 57 | р. Унья, 68 км от устья (левый берег) | 61°31′35′′ | 58°13′06′′ | 3 | 0 | 0 |
| 58 | р. Унья, 68 км от устья (правый берег) | 61°37′27′′ | 57°56′44′′ | 4 | 0 | 0 |
| 59 | Левый берег р. Печора, оз. Лешкатское, 1 км ниже | 61°42′30′′ | 57°27′19′′ | 2 | 0 | 0 |
Примечание. Se – раса Серов, So – раса Сок, F1 – гибриды между расами Сок и Серов. Порядковый номер локалитетов соответствует рис. 3.
Зверьки были отловлены в специализированные ловушки-живоловки [29] по специально разработанной методике отлова землероек-бурозубок [30].
Кариотипы большинства особей были получены по стандартной методике с предварительным внутрибрюшинным колхицинированием (0.2 мл, 0.04%, 1–1.5 ч), гипотонией клеток костного мозга и селезенки раствором хлорида калия (0.56%, 25–30 мин) и последующей трехкратной метанол–уксусной (3 : 1) фиксацией.
Кариотипы зверьков из локалитетов 31–33 (табл. 2) были получены посредством фиксации суспензий хромосом из первичной культуры фибробластов по описанной ранее методике [13].
Идентификацию гомологичных хромосом проводили с помощью G-окраски после обработки раскапанных на предметные стекла суспензий хромосом раствором 0.25%-ного трипсина с последующей 2-минутной окраской в 2%-ном красителе Гимза. Расовая принадлежность особей установлена в соответствии с международной номенклатурой для стандартного кариотипа S. araneus [6].
РЕЗУЛЬТАТЫ
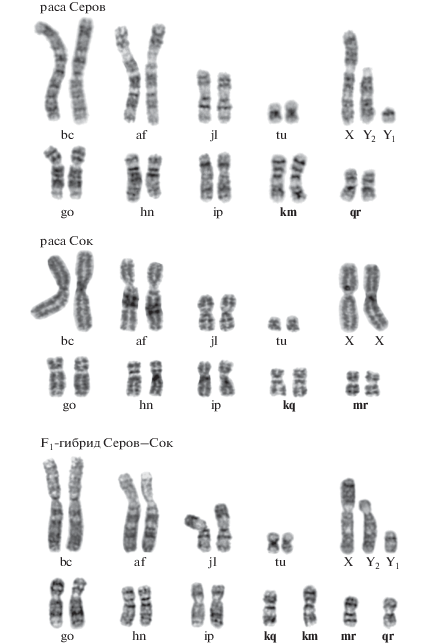
Цитогенетический анализ бурозубок, отловленных в 2011–2017 гг. (табл. 2), выявил наличие трех кариотипических вариантов – “чистая” раса Сок с диагностическими метацентриками go, hn, ip, kq, mr (N = 12), “чистая” раса Серов go, hn, ip, km, gr (N = 69) и межрасовые гибриды F1 (N = 11). Кариотипы этих рас различаются по двум парам хромосом с монобрахиальной гомологией (kq, mr-km, gr) и, как следовало ожидать, в кариотипе гибрида F1 были выявлены три пары гомологичных хромосом и четыре непарных метацентрика – go, hn, ip, qr-kq-km-mr (рис. 2). Диплоидное число хромосом у “чистых” рас и гибридов было постоянным 2n = 20 у самок и 2n = 21 у самцов (за счет полового тривалента XY1Y2) и NFa = 18, за исключением трех особей, полиморфных по метацентрику j/l. Данная гетерозиготность была обнаружена в кариотипах обеих “чистых” рас и гибрида F1, NFa = 19 (локалитеты 37, 48, 53). В мейозе у гибридов F1 ожидается формирование мейотической фигуры в виде кольца из четырех элементов – RIV подобно гетерозиготам из других гибридных зон со схожим уровнем кариотипических различий между родительскими расами [4].
Рис. 2.
G-окрашенные кариотипы обыкновенной бурозубки S. araneus из зоны контакта хромосомных рас Сок и Серов. Половые хромосомы обозначены XX (самка) и XY1Y2 (самец). Жирным шрифтом выделены метацентрики с монобрахиальной гомологией.
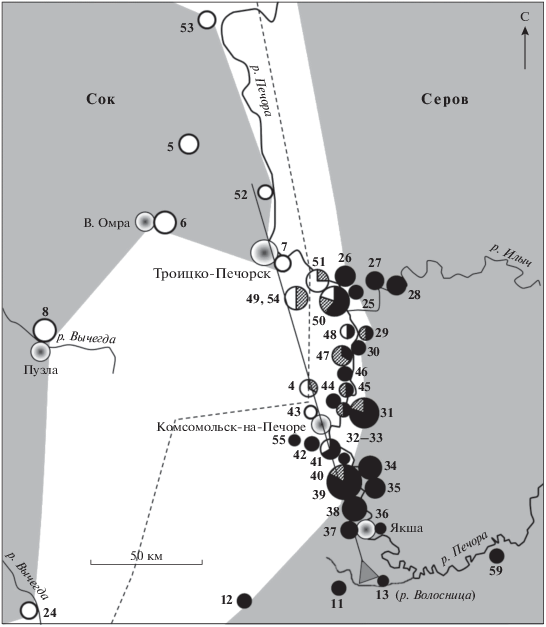
Совокупность всех данных (табл. 1 и 2) дает представление о локализации гибридной зоны Сок–Серов в исследуемом районе (рис. 3). Зона контакта образуется в междуречье Печоры и Вычегды. В районе пос. Комсомольск-на-Печоре ареал расы Сок доходит до р. Печора, которая далее разделяет зону. На левом берегу Печоры выявлены обе родительские расы и гибриды (локалитеты 4, 11–13, 37–51, 54), однако ниже пос. Троицко-Печорск обнаружены только кариотипы “чистой” расы Сок (локалитеты 5–7, 52, 53). Отметим, что еще ниже по течению на левом берегу Печоры был обнаружен кариотип гибрида F1 между расами Серов и Печора (локалитет 1, табл. 1). На правом берегу Печоры выявлены только кариотипы расы Серов (локалитеты 25–31, 34–36) и два гибрида F1 (локалитеты 29 и 31). На большом острове, отделенном от левого берега сравнительно неширокой протокой (локалитет 32, 33 “остров”), обнаружен один гибрид F1 и раса Серов (табл. 2, рис. 3).
Рис. 3.
Распределение кариотипических вариантов в зоне контакта хромосомных рас Сок и Серов и положение трансекта относительно локалитетов с известным кариотипом. Размер круга пропорционален величине выборки (макс. N = 6); черный цвет – раса Серов, белый – раса Сок, штриховка – гибриды. Области распространения родительских рас, исключая гибриды, показаны заливкой. Пунктир – изоклина (0.5) частот диагностических хромосом kq, mr (Сок)/km, /qr (Серов). Начало трансекта обозначено серым треугольником (локалитет 13).
Таким образом, общая картина зоны контакта Сок–Серов выглядит довольно сложной. В междуречье Печоры и Вычегды встреча этих рас возможна лишь в направлении, перпендикулярном реке, а от места встречи (Комсомольск-на-Печоре) их ареалы следуют вдоль разных берегов (рис. 3). Очевидно, что река вносит существенный вклад в распределение особей родительских рас, однако имеющиеся выборки не позволяют корректно оценить ширину гибридной зоны в зависимости от ее ориентации относительно речного русла. В связи с этим расчет ширины зоны был проведен только на трансекте вдоль реки.
Для построения клины были использованы только данные, полученные на левом берегу Печоры. Трансект общей длиной 180 км проложен вдоль левого берега реки с основанием в локалитете 13 и до локалитета 52 (рис. 3). Такая протяженность позволяет захватить крайние южные и северные участки, где надежно были обнаружены только кариотипы “чистых” родительских рас. Для расчета частот кариотипических вариантов общее расстояние было разделено на “подзоны” по 10 км, перпендикулярные курсу трансекта, с целью объединения локалитетов в одну “точку” и избегания малого числа особей (учитывалось не менее 4 особей). При отсутствии кариотипированных особей “подзону” исключали из расчета и, таким образом, анализ был проведен по 12 “точкам” (табл. 3, рис. 4).
Таблица 3.
Число особей Sorex araneus различных кариотипических категорий и частоты диагностических хромосом в гибридной зоне
| Дистанция от локалитета 13, км | Особи (N) | Частоты хромосом | ||||
|---|---|---|---|---|---|---|
| Se | F1 | So | всего | kq, mr (So) |
km, qr (Se) |
|
| 0 | 5 | 0 | 0 | 5 | 1 | 0 |
| 10 | 5 | 0 | 0 | 5 | 1 | 0 |
| 20 | 5 | 0 | 0 | 5 | 1 | 0 |
| 30 | 8 | 1 | 1 | 10 | 0.85 | 0.15 |
| 40 | 0 | |||||
| 50 | 4 | 0 | 1 | 5 | 0.80 | 0.2 |
| 60 | 3 | 2 | 2 | 7 | 0.57 | 0.43 |
| 70 | 2 | 2 | 1 | 5 | 0.60 | 0.4 |
| 80 | 0 | 2 | 2 | 4 | 0.25 | 0.75 |
| 90 | 3 | 2 | 4 | 9 | 0.44 | 0.56 |
| 100 | 0 | 0 | 4 | 4 | 0 | 1 |
| 110 | 0 | |||||
| 120 | 0 | |||||
| 130 | 0 | |||||
| 140 | 0 | 0 | 4 | 4 | 0 | 1 |
| 150 | 0 | |||||
| 160 | 0 | |||||
| 170 | 0 | 0 | 9 | 9 | 0 | 1 |
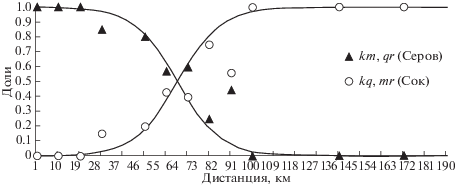
Кривая ожидаемого распределения лучше всего совпадает с фактическим распределением частот встречаемости диагностических хромосом для положения центра на расстоянии около 70 км от опорной точки (локалитет 13) к северу от пос. Комсомольск-на-Печоре и коэффициента A = 0.05. Соответственно, стандартная ширина клины составила 40 км (рис. 4). Фактическое число гибридов в центре зоны (см. табл. 3) не отличалось от ожидаемого для свободного скрещивания 50% (с поправкой Йетса χ2 = 0.24, p > 0.6).
ОБСУЖДЕНИЕ
В отличие от большинства изученных гибридных зон, зона Сок–Серов расположена на территории, которая подверглась минимальной антропогенной трансформации, что позволяет изучать процессы, не осложненные антропогенным изменением ландшафта. Заметим, что у обыкновенной бурозубки и популяционная структура в зоне гибридизации, и сама локализация гибридной зоны часто определяются изменениями ландшафта вследствие хозяйственной деятельности [18, 38]. Однако такое положение гибридной зоны имеет существенный минус. Труднодоступность большей части равнинной территории требует специальных исследований и в настоящее время у нас нет таких возможностей. Вместе с тем, даже предварительное обследование этой гибридной зоны в районе верхней Печоры выявило ряд вопросов, решение которых может оказаться значимым для понимания ранних этапов дивергенции вида.
Гибридная зона Сок–Серов имеет сложную конфигурацию. На бóльшей части территории расы встречаются в однотипном ландшафте средней тайги, и их ареалы не разделены существенными физическими преградами. Граница между расами Сок и Серов доходит до р. Печора и далее продолжается вдоль ее русла. По-видимому, реку пересекают только особи расы Серов, поскольку чистая раса Сок не отмечена на правом берегу реки. Заметим, что реки такого размера не являются непреодолимой преградой для бурозубок. В специальном исследовании при отлове зверьков на заякоренных на воде плотиках в верховьях р. Илыч (Печоро-Илычский заповедник, Республика Коми) было показано, что землеройки, в том числе обыкновенные бурозубки, довольно часто попадаются в ловушки на плотиках, т.е. плавают через реку [31]. Косвенным свидетельством интенсивности процесса миграции через реку является довольно высокая частота обнаружения этого вида в желудках европейского хариуса [32]. Наконец, появление гибридов F1 Сок–Серов на правом берегу Печоры возможно лишь при непосредственном пересечении ими реки, поскольку локалитеты, где встречены обе родительские расы, известны лишь с левого берега.
Можно предположить, что дистанция расселения гибридов больше, чем особей родительских рас. Так, в гибридной зоне между расами Кириллов и Печора, которая расположена на правом берегу р. Мезень (дер. Кысса Архангельской обл.), “чистая” раса Печора также не была обнаружена на левом берегу. Лишь один гибрид “пересек” реку и еще одна гибридная особь была поймана на речном острове [13]. Выше по течению Мезени, в районе д. Вожгора, гибрид Кириллов–Печора был пойман на правом берегу, где отсутствовали “чистые” особи расы Кириллов [21].
Аналогичная ситуация наблюдается и в гибридной зоне между расами Нерусса и Пенза, где гибриды отмечены лишь на одном берегу р. Аза в отсутствие одной из рас [33]. В гибридной зоне Москва–Селигер гибриды также встречены на краях зоны, там, где не были обнаружены особи обеих родительских рас [11]. В связи с этим уместно вспомнить, что, изучая заселение островков на озере в Финляндии, Хански и коллеги с помощью индексов асимметрии показали, что вселяющиеся на них обыкновенные бурозубки представлены социальными субординантами [34]. С учетом того, что на правом берегу Печоры, противоположном ареалу расы Сок, и на острове хромосомы этой расы встречены только у гибридных особей, можно предполагать, что и в этом случае лишь гибриды Сок–Серов пересекают реку и, следовательно, с большей вероятностью являются социальными субординантами. Разумеется, материал, которым мы располагаем, недостаточен даже для предварительной оценки подобного явления. Однако нам представляется важным обратить внимание на этот факт, так как социальная субординантность гибридов может являться одним из существенных факторов формирования гибридной зоны.
В этом случае поддержание парапатрии между контактирующими формами может быть обеспечено за счет перераспределения в пространстве особей с разными стратегиями расселения. Ранее нами было показано, что такая модель достаточна для объяснения стабилизации гибридной зоны Москва–Селигер, причем не требуется включать в нее отбор против гибридов [35]. Заметим, что этот же принцип мог бы объяснить подразделение обыкновенных бурозубок, как морфологическое [36, 37], так и генетическое [38, 39], на хорошо различимые локальные популяции внутри рас.
Расы Сок и Серов различаются двумя парами метацентриков с монобрахиальной гомологией, и в мейозе у гибридов ожидается формирование фигуры в виде кольца из четырех элементов (RIV). Известно несколько подобных гибридных зон и их стандартная ширина составляет от 1 до 14 км. Из них узкие зоны обнаружены между расами Кириллов и Печора – 1 км [21] и шведскими Uppsala и Hällefors – 3 км [19], которые разделены реками. В случае польских рас Drnholec и GuzowyMłyn [18] и Drnholec и Łęgucki Młyn [40, 41] ширина составляет 13 и 10–14 км соответственно. К сожалению, у нас не хватает данных для построения нескольких разнонаправленных клин. Вместе с тем следует обратить внимание на то, что стандартная ширина гибридной зоны Сок–Серов, полученная в нашем исследовании (40 км), явно больше, чем можно было бы ожидать для такого уровня различий кариотипов. Мы можем предложить несколько объяснений такого результата.
Во-первых, увеличение протяженности зоны гибридизации рас может происходить благодаря “растеканию” расселяющихся зверьков вдоль реки. В этом случае в излучинах могут образовываться “карманы”, в которых накапливаются особи родительских рас. Это объяснение отличается от “классического” карманного эффекта, обнаруженного у рас Łęgucki Młyn и Popielno, где последняя оказывается ограниченной с трех сторон и накапливается в излучине реки [20]. Однако если граница не перпендикулярна речному руслу, такой эффект вполне допустим.
Вместе с тем обратим внимание на то, что, гибриды Сок–Серов обнаружены не только в непосредственной близости от реки, но и в двух локалитетах на удалении 8 и 12 км от русла Печоры, и именно они оказывают влияние на ширину клины. На этом расстоянии влияние реки, а соответственно и “карманный эффект”, не должно было бы сказываться. На протяженность зоны вдоль трансекта может сказываться приток особей расы Серов, мигрирующих через реку или проходящих вдоль левого берега. Эта раса, по-видимому, заселяет правый берег Печоры как минимум до переправы на Вуктыл (локалитет 1), где на левом берегу обнаружен гибрид Серов–Печора [21]. Однако и в этом случае продвижение расы по берегу не объясняет наличия гибридов на удалении 8 и 12 км от реки.
Во-вторых, можно допустить, что ширина гибридной зоны определяется не только различиями в кариотипах, но и временем существования зоны. В процессе контакта обе формы, подверженные отбору против гибридов, приобретают дополнительные механизмы, препятствующие свободному скрещиванию. Такие механизмы возникают как усиление изоляции (reinforcement) при продолжительном ограничении потока генов. Именно приобретение дополнительных механизмов изоляции ответственно за формирование бимодальных зон (с преобладанием родительских форм) и именно на этом основании бимодальность гибридных зон свидетельствует, что формы “встали на путь видообразования” [12].
Можно предположить, что скрещивание недавно разделившихся рас не осложнено дополнительными механизмами изоляции. В случае гибридизации рас Сок и Серов мы пока не обнаруживаем дефицита гибридов, характерного для большинства известных гибридных зон у этого вида. Есть ли у нас основания считать, что гибридная зона Сок–Серов – одна из наиболее молодых гибридных зон?
Прежде всего об этом свидетельствует родственность кариотипов. Кариотип каждой из рас может произойти один из другого в результате одной транслокации WART. Такой вариант был первоначально предложен для эволюции группы финских рас [42]. Кариотип расы Серов был также предсказан на основе общей логики возможных трансформаций кариотипов хромосомных рас в результате WART [43] и впоследствии действительно обнаружен в природе [44]. Позже было показано, что в результате ряда последовательных изменений типа WART хромосомные расы, обитающие в Сибири, могут быть связаны со Скандинавскими расами [44]. Рассматривая распределение хромосомных рас по территории России, мы выделили четыре “кариотипические цепочки”, в которых кариотип одной расы может быть трансформирован из другой в результате одной мутации WART, причем, как и у других авторов [44, 45], расы Сок и Серов являются последовательными звеньями одной цепочки [8]. Анализ распределения и расселения рас S. araneus по территории в послеледниковый период (или в постледниковое время) позволяет говорить о недавнем (менее 8 тыс. лет назад) разделении этих рас. Причем, как считают многие авторы, расообразование – продолжающийся процесс [4, 7, 45–49] и судя по положению рас Сок и Серов в “цепочке” – это одно из наиболее молодых событий [8].
Помимо прочего, представляет интерес и тот факт, что распространение расы Сок на север ограничено сравнительно неширокой речной преградой, пространство за которой заполнено другой расой (Серов). Для сравнения заметим, что для того, чтобы заселить свой современный ареал, расе Кириллов нужно было пересечь р. Северная Двина в среднем течении, расе Мантурово – р. Вычегда в нижнем течении, а расе Сок – р. Кама [13]. Анализируя распространение хромосомных рас по современному ареалу, мы рассматривали случаи пересечения крупных рек и обнаружили, что если противоположный берег не занят иной расой, то водная преграда не имеет существенного значения в качестве препятствия для расселения особей другой расы [8]. Рассмотренная в настоящей публикации гибридная зона – один из таких примеров.
Авторы признательны к. б. н. В.М. Анискину (ИППЭ РАН) и д. б. н. А.В. Буракову (МГУ) за помощь в сборе полевого материала.
Работа частично была поддержана грантами РФФИ 15-04-04759 (С.П.) и 15-04-02531 (А.К.), а также Программой Президиума РАН “Биоразнообразие и биологические ресурсы России” (Программа 41) (С.П.).
Список литературы
Hewitt G.M. Hybrid zones – natural laboratories for evolutionary studies // Trends Ecol. Evol. 1988. V. 3. P. 158–165. doi 10.1016/0169-5347(88)90033-X
Barton N.H., Gale K.S. Genetic analysis of hybrid zones // Hybrid Zones and the Evolutionary Process. N.Y.: Oxford Univ. Press, 1993. P. 13–45.
Barton N.H., Hewitt G.M. Adaptation, speciation and hybrid zones // Nature. 1989. V. 341. P. 497–503.
Searle J.B., Wójcik J.M. Chromosomal evolution: the case of Sorex araneus. Survey of hybrid zones // Evolution of shrews. Białowieża: Mammal Research Institute Polish Acad. Sci., 1998. P. 243–253.
White T.A., Bordewich M., Searle J.B. A network approach to study karyotypic evolution: the chromosomal races of the common shrew (Sorex araneus) and house mouse (Mus musculus) as model systems // System. Biol. 2010. V. 59. P. 262–276. doi 10.1093/sysbio/syq004
Searle J.B., Fedyk S., Fredga K. et al. Nomenclature for the chromosomes of the common shrew (Sorex araneus) // Mem. Soc. Vaud. Sci. Nat. 1991. V. 19. P. 13–22.
Häusser J., Fedyk S., Fredga K. et al. Definition and nomenclature of the chromosome races of S. araneus // Folia Zool. 1994. V. 43. Suppl. 1. P. 1–9.
Shchipanov N.A., Pavlova S.V. Density-dependent processes determine the distribution of chromosomal races of the common shrew Sorex araneus (Lipotyphla, Mammalia) // Mammal. Res. 2017. V. 62. № 3. P. 267–282. doi 10.1007/s13364-017-0314-4
Pavlova S.V., Borisov S.A., Timoshenko A.F., Sheftel B.I. “European” race-specific metacentrics in East Siberian common shrews (Sorex araneus): a description of two new chromosomal races, Irkutsk and Zima // Comparative Cytogenet. 2017. V. 11. № 4. P. 797–806. doi 10.3897/CompCytogen.v11i4.19800
Павлова С.В., Анискин В.М., Щипанов Н.А. Новая хромосомная раса Sorex araneus (Lipotyphla, Mammalia) обнаружена на территории России // Генетика. 2017. Т. 53. № 12. С. 1453–1457.
Bulatova N., Jones R.M., White T.A. et al. Natural hybridization between extremely divergent chromosomal races of the common shrew (Sorex araneus, Soricidae, Soricomorpha): hybrid zone in European Russia // J. Evol. Biol. 2011. V. 24. P. 573–586. https://doi.org/ 10.1111/j.1420-9101.2010.02191.x
Jiggins C.D., Mallet J. Bimodal hybrid zones and speciation // Trends. Ecol. Evol. 2000. V. 15. P. 250–255. https://doi.org/10.1016/S0169-5347(00)01873-5
Pavlova S.V., Shchipanov N.A. A hybrid zone between the Kirillov and Petchora chromosomal races of the common shrew (Sorex araneus L., 1758) in northeastern European Russia. Preliminary description // Acta Theriol. 2014. V. 59. P. 415–426. doi 10.1007/s13364-014-0183-z
Polyakov A.V., White T.A., Jones R.M. et al. Natural hybridization between extremely divergent chromosomal races of the common shrew (Sorex araneus, Soricidae, Soricomorpha): hybrid zone in Siberia // J. Evol. Biol. 2011. V. 24. P. 1393–1402. doi 10.1111/j.1420-9101.2011.02266.x
Орлов В.Н., Борисов Ю.М., Ирхин С.Ю., Ковалева А.А. Особенности зоны контакта трех хромосомных рас обыкновенной бурозубки Sorex araneus L. (Mammalia) как показатель конкуренции популяций // Экология. 2010. № 6. С. 459–463.
Орлов В.Н., Борисов Ю.М., Черепанова Е.В. и др. Узкая гибридная зона хромосомных рас Москва и Западная Двина и особенности изоляции популяций обыкновенной бурозубки Sorex araneus (Mammalia) // Генетика. 2012. Т. 48. № 1. С. 80–88.
Mercer S.J., Searle J.B. Preliminary analyses of a contact zone between karyotypic races of the common shrew (Sorex araneus) in Scotland // Mémoir. Soc. Vaudoise Sci. Natur. 1991. V. 19. P. 73–78.
Szałaj K.A., Fedyk S., Banaszek A. et al. A hybrid zone between two chromosome races of the common shrew, Sorex araneus, in eastern Poland: preliminary results // Hereditas. 1996. V. 125. P. 169–176. https://doi.org/ 10.1111/j.1601-5223.1996.00169.x
Narain Y., Fredga K. A hybrid zone between the Hällefors and Uppsala chromosome races of Sorex araneus in central Sweden // Hereditas. 1996. V. 125. P. 137–145.
Moska M. A hybrid zone between the chromosomal races Łęgucki Młyn and Popielno of the common shrew in north-eastern Poland: preliminary results // Acta Theriol. 2003. V. 48. P. 441–455. https://doi.org/ 10.1007/BF03192491
Shchipanov N.A., Pavlova S.V. Contact zones and ranges of chromosomal races of the common shrew, Sorex araneus, in northeastern European Russia // Folia Zool. 2013. V. 62. P. 24–35. https://doi.org/ 10.25225/fozo.v62.i1.a4.2013
Дегтева С. Растительные сообщества // Девственные леса Коми / Ред. Таскаев А.И. М.: Феория, 2005. С. 108–133.
Щипанов Н.А., Бобрецов А.В., Булатова Н.Ш. и др. Уральская хромосомная раса Серов обыкновенной бурозубки (Sorex araneus L.: Insectivora, Mammalia) в полидоминантной тайге на Севере европейской части России // Докл. Акад. наук. 2005. Т. 404. № 2. С. 281–285.
Щипанов Н.А., Булатова Н.Ш., Бобрецов А.В., Демидова Т.Б. Хромосомные расы обыкновенной бурозубки (Sorex araneus L.) на Европейском северо-востоке. Ограничивают ли физические преграды их распространение? // Докл. Акад. наук. 2008. Т. 422. № 5. С. 714–717.
Щипанов Н.А., Булатова Н.Ш., Опарин М.Л. Островная популяция расы Сок обыкновенной бурозубки (Investivora: Мammalia) на южной границе ареала // Докл. Акад. наук. 2002. Т. 386. № 3. С. 427–429.
Shchipanov N.A., Bobretsov A.V., Pavlova S.V., Demidova T.B. Inhabited areas and new hybrid zones of the common shrew (Sorex araneus L.) at the European north-eastern part of Russia // Proceedings of International Conference “Problems of animal study and protection in the North”. Syktyvkar: Komi Sci. Centre UrB RAS, 2009. P. 142–144.
Searle J.B. Factors responsible for a karyotypic polymorphism in the common shrew, Sorex araneus // Proc. R. Soc. L. B. 1986. V. 229. P. 227–298.
Fedyk S. Geographic chromosomal differentiation and hybrid zones between chromosome races of Sorex araneus L. in north-eastern Poland: Dissertat. Univ. Varsov. Białystok (in Polish), 1995.
Щипанов Н.А. К экологии малой белозубки (Crocidura suaveolens) // Зоол. журн. 1986. Т. 66. № 7. С. 1051–1060.
Щипанов Н.А., Калинин А.А., Олейниченко В.Ю. и др. К методике изучения использования пространства землеройками-бурозубками // Зоол. журн. 2000. Т. 79. № 3. С. 362–371.
Калинин А.А., Куприянова И.Ф. Методика количественного учета мелких млекопитающих при миграциях через водные преграды // Зоол. журн. 2015. Т. 94. № 3. С. 365–369.
Калинин А.А., Куприянова И.Ф. Мелкие млекопитающие в питании европейского хариуса (Thymallusthymallus, Thymallidae, Salmoniformes) // Зоол. журн. 2016. Т. 95. № 6. С. 712–719.
Черепанова Е.В., Кривоногов Д.М., Щегольков А.В. и др. Формирование зон контакта хромосомных рас обыкновенной бурозубки (Sorex araneus, Soricomorpha) в бассейне Bолги // Зоол. журн. 2018. Т. 97. № 6. С. 723–734.
Hanski I., Peltonen A., Kaski L. Natal dispersal and social dominance in the common shrew Sorex araneus // Oicos. 1991. V. 62. P. 48–58. doi 10.2307/3545445
Щипанов Н.А., Булатова Н.Ш., Павлова С.В. Распределение обыкновенных бурозубок (Sorex araneus L.) двух хромосомных рас в зоне интерградации. Может ли изменение типа расселения поддерживать независимость генных частот // Генетика. 2008. Т. 44. № 6. С. 734–745.
Polly P.D. Phylogeographic differentiation in Sorex araneus: morphology in relation to geography and karyotype // Rus. J. Theriol. 2007. V. 6. P. 73–84.
Щипанов Н.А., Бобрецов А.В., Куприянова И.Ф., Павлова С.В. Межрасовая и межпопуляционная изменчивость фенотипических (краниальных) признаков обыкновенных бурозубок (Sorex araneus L., 1758) // Генетика. 2011. Т. 47. № 1. С. 76–86.
Ratkiewicz M., Banaszek A., Jadwiszczak K.A. et al. Genetic diversity, stability of population structure and barriers to gene flow in a hybrid zone between two Sorex araneus chromosome races // Mammalia. 2003. V. 67. P. 275–284.
Horn A., Basset P., Yannic G. et al. Chromosomal rearrangements do not seem to affect the gene flow in hybrid zones between karyotypic races of the common shrew (Sorex araneus) // Evolution. 2012. V. 66. P. 882–889. https://doi.org/10.1111/j.1558-5646.2011.01478.x
Fedyk S., Chętnicki W., Banaszek A. Genetic differentiation of Polish populations of Sorex araneus L. III. Interchromosomal recombination in a hybrid zone // Evolution. 1991. V. 45. P. 1384–92. https://doi.org/ 10.1111/j.1558-5646.1991.tb02643.x
Fedyk S., Chętnicki W. Whole–arm reciprocal translocation in a hybrid population of Sorex araneus // Chromosome Res. 2009. V. 17. P. 451–454. https://doi.org/ 10.1007/s10577-009-9036-z
Halkka L., Soderlund V., Skaren U., Heikkila J. Chromosomal polymorphism and racial evolution of Sorex araneus // Hereditas. 1987. V. 106. P. 257–275. doi 10.1111/j.1601-5223.1987.tb00260.x
Polyakov A.V., Borodin P.M., Lukáčová L. et al. The hypothetical Old-Northern chromosome race of Sorex araneus found in the Ural Mts // Ann. Zool. Fennica. 1997. V. 34. P. 139–142. http://www.jstor.org/stable/23735690.
Polyakov A.V., Zima J., Searle J.B. et al. Chromosome races of the common shrew Sorex araneus in the Ural Mts: a link between Siberia and Scandinavia? // Acta Theriol. 2000. V. 45. P. 19–26.
Поляков А.В., Панов В.В., Ладыгина Т.Ю. и др. Хромосомная эволюция обыкновенной бурозубки Sorex araneus L. в послеледниковое время на Южном Урале и в Сибири // Генетика. 2001. Т. 37. № 4. С. 448–455.
Häusser J., Catzeflis F., Meylan A., Vogel P. Speciation in the Sorex araneus complex (Mammalia, Insectivora) // Acta Zool. Fennica. 1985. V. 170. P. 125–130.
Häusser J. The Sorex of the araneus-arcticus group (Mammalia, Soricidae): do they actually speciate // Special Publ. Carnegie Museum Natural History. 1994. V. 18. P. 295–306.
Wójcik J.M. Chromosome races of the common shrew Sorex araneus in Poland: a model of a karyotype evolution // Acta Theriol. 1993. V. 38. P. 315–338.
Wójcik J.M., Ratkiewicz M., Searle J.B. Evolution of the common shrew Sorex araneus: chromosomal and molecular aspects // Acta Theriol. 2002. V. 47 (Suppl. 1). P. 139–167. https://doi.org/ doi 10.1007/BF03192485
Дополнительные материалы отсутствуют.