

Генетика, 2019, T. 55, № 5, стр. 551-556
Реконструкция митохондриального генома древней лошади из городища Ашна-Пандо (Среднее Поволжье)
Е. И. Антонова 1, *, А. В. Соловьев 1, Л. А. Вязов 2, Ю. А. Семыкин 3, А. В. Мищенко 4
1 Ульяновский государственный педагогический университет им. И.Н. Ульянова, Научно-исследовательский центр фундаментальных и прикладных проблем биоэкологии и биотехнологии
432071 Ульяновск, Россия
2 Казанский федеральный университет, Институт международных отношений, истории и востоковедения,
кафедра истории Татарстана, археологии и этнологии
420008 Казань, Россия
3 Ульяновский государственный педагогический университет им. И.Н. Ульянова,
кафедра истории
432071 Ульяновск, Россия
4 Ульяновский государственный педагогический университет им. И.Н. Ульянова,
кафедра географии и экологии
432071 Ульяновск, Россия
* E-mail: antonov_67@mail.ru
Поступила в редакцию 12.06.2018
После доработки 12.07.2018
Принята к публикации 18.10.2018
Аннотация
Проведена реконструкция митохондриального генома лошади, известной по костным останкам из раскопок городища Ашна-Пандо (Среднее Поволжье, Ульяновская обл., бассейн р. Сура). Установлено, что по фрагменту контрольного района Д-петли данная особь принадлежала к гаплотипу В1, распространенному среди как современных, так и древних лошадей Европы и Азии, тогда как по полному митохондриальному геному изученная особь относится к гаплогруппе М, характерной для различных современных пород, в том числе ахалтекинской. Полученные данные позволяют предполагать “степное” происхождение исследованной лошади. Представляется наиболее вероятным, что проникновение лошадей южного происхождения относительно далеко в глубь зоны широколиственных лесов относится ко второй–третьей четверти I тыс. до н.э. и связано с контактами оседлого населения региона с кочевниками скифского круга.
В последнее время спектр методик, применяемых для реконструкции историко-культурных процессов в древности и средневековье, неуклонно расширяется. Одной из исследовательских процедур, прочно вошедших в состав комплексного археологического исследования, стал молекулярно-генетический анализ. Реконструкция ДНК древних и средневековых домашних животных позволяет получить впечатляющие результаты, характеризующие как животноводство само по себе, так и направления культурных и экономических связей и контактов древнего населения. В настоящей статье представлены результаты анализа костных остатков лошади, одомашненной более 5 тыс. лет назад [1, 2] и оказывавшей определяющее воздействие на развитие экономики и военного дела вплоть до недавнего времени. Исследованный образец происходит с территории Среднего Поволжья, где ранее не проводились аналогичные исследования, что делает полученные результаты важными для пополнения базы сравнительных материалов.
Городище Ашна-Пандо расположено в Среднем Поволжье, на территории современной Ульяновской области, в среднем течении р. Суры. Памятник был описан и исследован П.Д. Степановым в 1949 г. Согласно результатам этих исследований он осваивался древним населением трижды: в эпоху бронзы (II тыс. до н.э.), раннего железного века (I тыс. до н.э.) и в эпоху Великого переселения народов (середина I тыс. н.э.). Полученные в ходе исследования материалы включают в себя, помимо артефактов, значительную коллекцию костей животных. Среди них – кости сельскохозяйственных (крупный и мелкий рогатый скот, лошадь, свинья) и различных диких видов (лось, медведь, бобр, лисица, косуля, кабан и др.; видовые и возрастные определения проведены в 2014–2015 гг. ст. н. сотр. Института археологии Академии наук Республики Татарстан Г.Ш. Асылгараевой). Все остеологические материалы памятника, за исключением отдельных костей человека и орудий, изготовленных из кости, представляют собой кухонные остатки. Остеологическая коллекция памятника имеет самостоятельное значение в качестве источника по реконструкции хозяйственной деятельности его обитателей в разные эпохи.
Комплекс находок свидетельствует о том, что территория памятника в эпоху бронзы была освоена населением балановской культуры, существовавшей в 13–12 вв. до н.э. [3]. Анализ археозоологических коллекций балановских памятников свидетельствует, что население вело животноводческое хозяйство, основанное на разведении четырех сельскохозяйственных видов, дополнявшееся охотой и, возможно, примитивным земледелием.
Второй этап освоения, приходящийся на ранний железный век, представлен существенно большим числом находок. Керамический комплекс весьма своеобразен. Наиболее близкие в культурном отношении материалы были выявлены на городище Ош-Пандо П.Д. Степановым в 1946–1948 гг. [4]. Вероятно, население в культурном отношении было близким к носителям городецкой культуры и вело животноводческое хозяйство при минимальной роли охоты и земледелия [5]. На основании аналогий псалию из клыка кабана, имеющихся в материалах юхновской культуры [6], можно предположить датировку древностей раннего железного века с городища Ашна-Пандо в пределах второй–третьей четверти I тыс. до н.э.
Поздний этап освоения городища Ашна-Пандо относится к эпохе раннего средневековья. В это время территория Среднего Посурья осваивается населением именьковской культуры IV–VII вв. н.э. [7].
Особенности вещевого материала именьковского комплекса городища Ашна-Пандо позволяют отнести его к позднему периоду развития именьковских древностей и датировать VI–VII вв. н.э. Хозяйство именьковского населения носило комплексный характер. Оно базировалось на сочетании пашенного земледелия с животноводством, охотой и рыболовством.
В ходе раскопок не было выявлено комплексов, период функционирования которых можно было бы однозначно связать с той или иной эпохой функционирования памятника. Единственным основанием для хронологической атрибуции той или иной группы находок являются общие стратиграфические наблюдения. Так, основная масса материала эпохи бронзы и раннего железного века сосредоточена на краю площадки памятника, где глубина культурных наслоений доходит до 1.5 м, в то время как раннесредневековые кухонные остатки присутствуют на всей площадке городища.
Образец, взятый для проведения генетического анализа, представляет собой верхнечелюстной зуб лошади возрастом 8–9 лет. Происходит из участка раскопа, расположенного на восточном краю площадки городища (траншея А, кв. 11), с уровня 9-го штыка (глубина 1.3 м от поверхности), что соответствует нижней части культурного слоя на склоне городища, в которой могут присутствовать как раннесредневековые, так и более ранние материалы. Наряду с образцом, взятым на анализ, из данного участка происходят четыре кости крупного рогатого скота (фрагменты ребра, лучевой кости, нижней челюсти и фаланга 1), три кости лошади (фрагменты нижней челюсти, бедренной и тазовой костей), таранная кость свиньи и трубчатая кость птицы. Археозоологический контекст и стратиграфические основания позволяют с большой долей вероятности отнести отобранный для анализа образец к комплексу раннего железного века и датировать его второй–третьей четвертью I тыс. до н.э.
МАТЕРИАЛЫ И МЕТОДЫ
Выделение древней мтДНК
Выделение древней ДНК проводили по методу, описанному ранее в статье [8]. Затем получали библиотеки для секвенирования с использованием TruSeq® DNA Sample Preparation Kit (Illumina, США), по протоколу производителя. Перед секвенированием проводили процедуру обогащения библиотек путем гибридизации с биотинилированной мтДНК современной лошади, иммобилизованной на Dynabeads® Streptavidin магнитных частицах (Life Technologies, США) по методу, описанному в статье [9].
Иммобилизация мтДНК современной лошади на Dynabeads® М-280 Streptavidin. Митохондриальную ДНК лошади нарабатывали в ПЦР в виде четырех фрагментов (около 4 тыс. пн каждый) с праймерами: 1F-gaggagcctgttccataatcg, 1R-ggttagggggaggagtagg, 2F-tcccatccacaaacaacataaa, 2R-gaggcttggagaagggtgaag, 3F-tgaccacccacaggtatcca, 3R-acgttggtggagtgttctagtt, 4F-ggcagcatttttgccggatt, 4R-gatggtggggtttatcgggg.
Эквимолярную смесь фрагментов биотинилировали с помощью Nick Translation Bionik Labeling System Kit (Invitrogen, США), очищали на колонках MinElute, денатурировали 5 мин при 95°С и иммобилизовали на магнитных частицах (предварительно промытых 3 × 2SSC) в растворе 2SSC 30 мин при комнатной температуре. Замеряли оптическую плотность в исходном растворе и в трех отмывках по 1 мл 2SSC и определяли количество иммобилизованной ДНК, обычно составляющее 1.2–1.7 мкг на 50 мкл исходной суспензии Dynabeads.
Гибридизация. Библиотеки, содержащие обычно около 900–1500 нг ДНК в 30 мкл 10 мМ трис-НСl буфера, денатурировали 5 мин при 95°С, добавляли 10 мкл Dynabeads® М-280 Streptavidin – магнитных частиц, несущих на себе 0.24–0.34 мкг биотинилированной мтДНК, добавляли 0.1 V 20SSC и проводили гибридизацию при 65°С 48–72 ч. Затем частицы промывали 3 × 100 мкл 2SSC при 65°С, 2 × 0.2SSC при комнатной температуре и элюировали в 30 мкл воды. Амплификацию обогащенных фрагментов древней ДНК из библиотек для секвенирования проводили в конечном объеме 50 мкл, содержащем Phusion HF Buffer, 0.25 мM 4dNTP, 1 мкМ праймеров для адаптеров фрагментов библиотек, 1 е.а. Phusion ДНК-полимеразы.
После квантификации полученных библиотек с использованием ПЦР-РВ в присутствии интеркалирующего красителя SYBR Green I проведено секвенирование на платформе MiSeq, ILLUMINA (использовали Reagent Kits v2, 500 циклов).
Редактирование и выравнивание полученных последовательностей, филогенетические реконструкции проводили с использованием программы Geneious 8.0.5.
Биоинформационная обработка данных
Множественное выравнивание нуклеотидных последовательностей проводили в программе MAFFT. В выравнивании полных митохондриальных геномов мы выделили пять функциональных групп последовательностей. Модели замен для каждой группы определили с помощью программы PartitionFinder: для вторых кодонов белок-кодирующих генов использовали модель HKY+G, для РНК-кодирующих генов, первых и третьих кодонов белок-кодирующих генов – HKY+G+I, а для гипервариабельных районов – GTR+G+I. Для построения филогенетического дерева использовали Байесовский метод (программа MrBayes): 2 000 000 генераций Марковской цепи, частота взятия образцов – 1000, первые 25% деревьев отбрасываются. Филогенетическая сеть построена после выравнивания последовательностей фрагмента 246 пн из гипервариабельного района I митохондриального генома лошадей на основе медианного (median joining) графа в программе NETWORK 4.612.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Из 2 784 180 полученных ридов 696 картированы на митохондриальный геном лошади. Сборка фрагментов позволила реконструировать 94.7% митохондриального генома лошади с покрытием 7.9 раз, средняя длина ридов составляет 188.3 пн (от 40 до 251 пн) (идентификационный номер в базе данных GenBank: MH605334).
Для анализа данных мы использовали две различные классификации: Сизлака с соавт. [10] по фрагменту контрольного района митохондриального генома, который охватывает большое число как современных, так и древних лошадей, но является менее точной, поскольку основывается на коротком фрагменте длиной 246 пн; вторая классификация Ачилли с соавт. [11] по последовательности полного митохондриального генома различных современных пород домашних лошадей является более точной, но недостаток данных по древним лошадям не позволяет изучить изменение популяций на различных территориях в течение времени.
Относительно начала 15 000 пн в референсной последовательности (X79547) были обнаружены замены 495C, 602T, 617C, 659C, 720A, характерные для гаплотипа B1 по классификации Сизлака с соавт. [10].
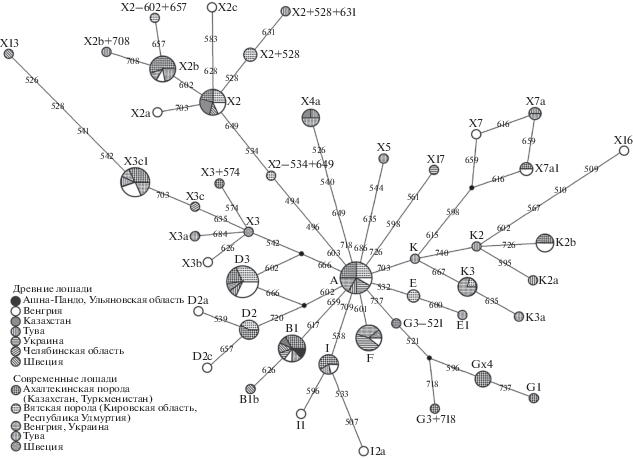
На основе выравнивания последовательностей фрагмента 246 пн из гипервариабельного района I митохондриального генома исследуемой лошади, древних и современных лошадей [10] мы построили филогенетическую сеть (рис. 1). Гаплотип В1 широко распространен в древних популяциях лошадей Казахстана, Швеции, Венгрии, Украины, среди современных лошадей ахалтекинской породы (Туркменистан, Казахстан) и лошадей Тувы.
Рис. 1.
Филогенетическая сеть гаплотипов мтДНК современных и древних лошадей Кировской обл., Челябинской обл., Тувы, Удмуртии, Казахстана, Украины, Венгрии, Швеции. Названия вершин сети соответствуют названиям гаплотипов по [10]. Размер кругов пропорционален числу особей, относящихся к гаплотипу. Разными заливками и штриховками обозначены популяции лошадей из разных районов. Цифры на узлах сети соответствуют позиции однонуклеотидной замены от начала 15 000 пн в референсной последовательности (X79547).
Согласно опубликованным данным, гаплогруппа B состоит из древних и современных лошадей Аляски, Европы (от Уральских гор и Каспийского моря до Пиренеев, включая Армению), Сибири, Казахстана и Малой Азии. Она включает в себя шесть гаплотипов: B, B1, B1a, B1b, B2, B3. Лошади гаплотипа B1 обитали в Европе и Малой Азии во времена до и после одомашнивания. Гаплотип В во времена позднего плейстоцена (126–11.7 тыс. лет назад) был распространен на Пиренейском полуострове, лошади гаплотипа В2 встречались на Аляске, а гаплотип В3 был распространен в Северной и Восточной Сибири. Тогда же гаплотип В1а встречался в Европе, гаплотип В1 – в Западной, Южной Сибири и Казахстане, в Европе (за исключением Пиренейского полуострова), в Малой Азии и Армении. В эпоху палеометалла (4000–3000 лет до н.э.) в Европе, Малой Азии и Армении был особенно распространен гаплотип В1, тогда как на Пиренейском полуострове доминировал гаплотип B вплоть до бронзового века (3500–1200 лет до н.э.). В раннем железном веке (с 1200 г. до н.э. до 340 г. н.э.) гаплотип В1 был распространен в тех же районах, что и в позднем плейстоцене, распространение данного гаплотипа дополнено в настоящей статье находкой с городища Ашна-Пандо (рис. 2). Гаплотип B1 в Европе (не включая Пиренейского полуострова), в Малой Азии и Армении встречается вплоть до средневековья.
Рис. 2.
Распространение гаплогруппы B. Гаплотипы: 1 – B2, AF326672 (Аляска, поздний плейстоцен); 2 – B3, DQ007574 (о. Большой Ляховский, поздний плейстоцен); 3 – B1, FJ204351 (Книгроте, 15 000–14 000 лет до н.э.); 4 – B, FJ204382 (пещера Фоске, 5210–4910 лет до н.э.); 5 – B1, FJ204360 (Маяки, 3250–3100 лет до н.э.); 6 – B, DQ683528 (Порталон, 2200–1970 лет до н.э.); 7 – B, FJ204391 (Вальядолид, 800 лет до н.э.–6 лет н.э.); 8 – B1, FJ204339 (Тува, 619–608 лет до н.э.); 9 – B1, AJ876889 (курган Берел, 300 лет до н.э.); 10 – B1, EU093038 (Среднедунайская низменность, 900 лет н.э.); 11 – B1, AF326676 (Южная Швеция, 200–500 лет н.э.); 12 – B1b, AF326679 (Южная Швеция, 200–500 лет н.э.); 13 – B1, Ашна-Пандо (Ульяновск, 600–300 лет до н.э.). Пунктирным овалом объединены находки древних лошадей гаплогруппы B1, наиболее близкие по времени существования к найденным останкам лошади из Ашна-Пандо.
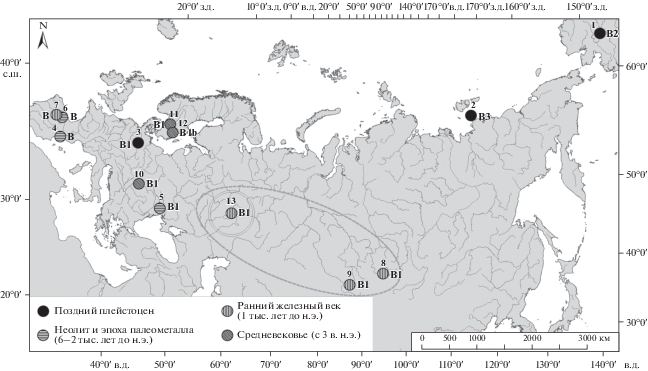
В железном веке в Европе широко распространился гаплотип В1b, а на Пиренейском полуострове все так же доминировал гаплотип B. Гаплотипы B1a, B1b, B2, B3 встречаются только среди древних лошадей. Современные лошади древних пород, принадлежащие к гаплогруппе В, редко встречаются в Китае, Марокко, Испании, чаще в Монголии, Греции, Турции, Болгарии, Румынии, Ливии, Египте, они наиболее распространены в Северной Европе [10].
По полному митохондриальному геному проанализированной кости лошади из городища Ашна-Пандо и последовательностям лошадей [11] было построено филогенетическое дерево (рис. 3).
Рис. 3.
Филогенетическое дерево современных лошадей из разных районов мира и древней лошади из захоронения в Ульяновской обл. A–R – названия митохондриальных гаплогрупп по классификации Ачилли и др. [11]. Цифры на ветвях дерева обозначают апостериорную вероятность топологии, полученную Байесовским методом. Образец “UL” соответствует изучаемому экземпляру лошади (идентификационный номер в GenBank: MH605334); образцы 60–64 соответствуют номерам анализируемых образцов в статье Ачилли и др. [11] (идентификационные номера в GenBank: JN398435, JN398436, JN398439, JN398437, JN398438 соответственно). В качестве аутгруппы взят осел (Equus asinus L.)
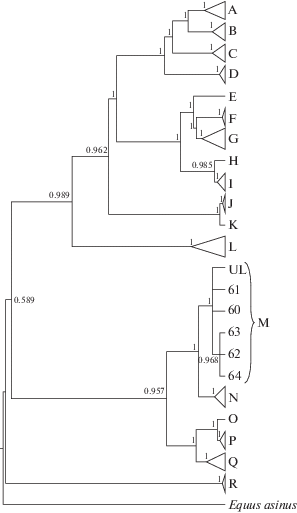
Исследованная лошадь принадлежит к митохондриальной гаплогруппе M по классификации Ачилли с соавт. [11], включающей следующие породы лошадей: ахалтекинская (Центральная Азия), Каспийский пони (Средний Восток), Клейдесдаль (шотландская хладнокровная лошадь) (Северная Европа), Маремано (Южная Европа), Фризская лошадь (Центральная Европа). Близкородственная к ней гаплогруппа N включает: шайр (Северная Европа), сэддлбред (Северная Америка), эксмурский пони (Северная Европа), андалузская (Южная Европа), неопределенная иранская порода (Ближний Восток). Следует отметить, что животные данной гаплогруппы не обнаруживались среди древних лошадей Южной Сибири [12].
Недостаток сравнительного материала не позволяет напрямую использовать полученные данные для культурно-исторических реконструкций. Наиболее вероятным представляется, что источником распространения лошадей данной гаплогруппы на территории лесной зоны Среднего Поволжья были степные территории, занятые в период, синхронный слою раннего железного века на городище Ашна-Пандо, кочевниками скифского и савроматского круга, через посредство которых в степной зоне и за ее пределами могли распространяться не только многие элементы материальной культуры, но и новые породы лошадей. На вероятность проникновения в лесную зону привозных породистых лошадей указывает, в частности, находка упомянутого выше псалия из клыка кабана [4], являющегося частью снаряжения верхового коня и бесполезного при хозяйственном использовании лошади.
Настоящим исследованием было установлено, что фрагмент кости лошади, полученный в результате раскопок городища Ашна-Пандо в бассейне р. Суры (Среднее Поволжье, Ульяновская обл., Россия) и на основании археологического контекста датированный второй–третьей четвертью I тыс. до н.э., принадлежит гаплотипу B1 (по короткому фрагменту, классификация Сислака с соавт. [10]), а по почти полному митохондриальному геному – представляет из себя новый гаплотип гаплогруппы М (по классификации Ачилли с соавт. [11]). Дальнейшие исследования позволят уточнить хронологическую интерпретацию и обогатить сравнительную базу, что необходимо для культурно-исторических реконструкций.
Коллектив авторов выражает благодарность А.С. Графодатскому, Н.В. Воробьевой, А.С. Дружковой, М.А. Куслий, В.А. Трифонову (Институт молекулярной и клеточной биологии СО РАН, Новосибирский государственный исследовательский университет, Новосибирск) за всестороннюю помощь при проведении исследований и интерпретации полученных результатов.
Исследование проведено при поддержке гранта РФФИ № 18-44-730006.
Список литературы
Lippold S., Matzke N.J., Reissmann M., Hofreiter M. Whole mitochondrial genome sequencing of domestic horses reveals incorporation of extensive wild horse diversity during domestication // BMC Evol. Biol. 2011. V. 11: 328. doi https://doi.org/10.1186/1471-2148-11-328
Wallner B., Vogl C., Shukla P. et al. Identification of genetic variation on the horse y chromosome and the tracing of male founder lineages in modern breeds // PLoS One. 2013. V. 8: e60015. doi https://doi.org/10.1371/journal.pone.0060015
Бадер О.Н., Халиков А.Х. Памятники балановской культуры // Свод археологических источников. М.: Наука, 1976. 168 с.
Степанов П.Д. Ош-Пандо Саранск: Мордовское кн. изд-во, 1967. 110 с.
Сарапулкина Т.В. Хозяйственно-культурный тип городецкого населения Верхнего Дона // Изв. Самарского научн. центра РАН. 2010. Т. 12. № 2. С. 278–281.
Каравайко Д.В., Горбаненко С.А. Господарство носіїв юхнівської культури. Київ: Наук. думка, 2012. 304 с.
Матвеева Г.И. Среднее Поволжье в IV–VII вв. н.э.: Именьковская культура. Самара: Самарский ун-т, 2004. С. 74–76.
Druzhkova A.S., Thalmann O., Trifonov V.A. et al. Ancient DNA analysis affirms the canid from Altai as a primitive dog // PLoS One. 2013. V. 8(3): e57754. doi https://doi.org/10.1371/journal.pone.0057754
Maricic T., Whitten M., Pääbo S. Multiplexed DNA sequence capture of mitochondrial genomes using PCR products // PLoS One. 2010. Vol. 5 (11): e14004. doi https://doi.org/10.1371/journal.pone.0014004
Cieslak M., Pruvost M., Benecke N. et al. Origin and history of mitochondrial DNA lineages in domestic horses // PLoS One. 2010. V. 5 (12): e15311. doi https://doi.org/10.1371/journal.pone.0015311
Achilli A., Olivieri A., Soares P. et al. Mitochondrial genomes from modern horses reveal the major haplogroups that underwent domestication // Proc. Natl Acad. Sci. USA. 2012. V. 109 (7). P. 2449–2454.
Куслий М.А., Дружкова А.С., Попова К.О. и др. Генотипирование и определение масти древних лошадей Бурятии // Цитология. 2016. Т. 58. С. 304–308.
Дополнительные материалы отсутствуют.