

Генетика, 2019, T. 55, № 8, стр. 978-982
Регрессионная модель времени цветения староместных сортов нута
К. Н. Козлов 1, *, М. Г. Самсонова 1, С. В. Нуждин 1, 2
1 Санкт-Петербургский политехнический университет им. Петра Великого,
научно-исследовательская лаборатория Математическая биология и биоинформатика,
Институт прикладной математики и механики
195251 Санкт-Петербург, Россия
2 Университет Южной Калифорнии, Программа молекулярной
и вычислительной биологии
24105 Лос-Анжелес, США
* E-mail: kozlov_kn@spbstu.ru
Поступила в редакцию 01.12.2018
После доработки 01.02.2019
Принята к публикации 12.02.2019
Аннотация
Разработаны регрессионные модели времени цветения староместных сортов нута из коллекции ВИР, собранных в Турции и Эфиопии. Предсказанное время цветения хорошо согласуется с опубликованными ранее данными. Различие моделей статистически значимо. Вклад температуры в модель для сортов из Эфиопии слабее, чем для сортов из Турции, 48 и 60% соответственно. Вклад осадков в модель составляет более 80%. Результаты согласуются с особенностями климата в местах сбора.
Зернобобовые культуры, такие как нут, служат основным источником растительного белка. Нут, обладая высокими пищевыми и вкусовыми достоинствами, имеет и целый ряд биологических преимуществ перед другими культурами. Он отличается высокой засухоустойчивостью, жаровыносливостью, устойчивостью к полеганию, осыпанию семян и вредителям, в частности такому злостному, как зерновка.
Время цветения является важным фенологическим признаком нута [1]. Точное его предсказание может помочь селекционерам в выборе родительских линий для создания сортов, оптимальных для конкретных регионов [2, 3].
Математическое моделирование стало одним из важнейших методов решения этой задачи. Стандартными методами моделирования фенологии являются расчет сумм температур выше температурного минимума и регрессионные модели [4–8]. Ранее нами был предложен метод построения нелинейных регрессионных моделей на основе грамматической эволюции, который был применен для моделирования фенологии сои [9].
Целью данного исследования было выявление различий реакции времени цветения сортов нута из коллекции ВИР, собранных в Турции и Эфиопии, на изменение агроклиматических факторов.
Образцы нута. Были рассмотрены агроклиматические данные, полученные в Сирии вблизи Алеппо для староместных сортов нута из коллекции ВИР, собранных в Турции и Эфиопии. Имеющиеся климатические данные представлены в табл. 1.
Таблица 1.
Климатические факторы
| Символ | Описание | Турция | Эфиопия |
|---|---|---|---|
| среднее ± стандартное отклонение | |||
| Tmean | Средняя годовая температура | 130.12 ± 30.59 | 173.30 ± 22.57 |
| ∆Tmean | Средний разброс температуры по месяцам | 115.97 ± 14.94 | 137.23 ± 9.65 |
| ∆T | Годовой разброс температуры | 310.06 ± 38.04 | 185.68 ± 17.73 |
| T sd | Сезонность температуры (100 × стандартное отклонение) | 6899.28 ± 1040.20 | 1172.47 ± 381.10 |
| P ann | Годовые осадки | 582.17 ± 151.76 | 1002.39 ± 255.42 |
| P sd | Сезонность осадков (100 × стандартное отклонение) | 53.60 ± 19.88 | 95.84 ± 21.37 |
Математическая модель времени цветения. Модель времени цветения строится как линейная комбинация N управляющих функций, зависящих от агроклиматических признаков нелинейным образом. Таким образом модель имеет вид (1):
(1)
${{{y}_{{\text{i}}}} = {{{\beta }}_{{\text{0}}}} + \sum\limits_{n = {\text{0}}}^{N - {\text{1}}} {{{{\beta }}_{{{\text{n}} + {\text{1}}}}}} {{F}_{{\text{n}}}}\left( {{{X}_{{\text{i}}}}} \right) + {{{\varepsilon }}_{{\text{i}}}}},$Аналитический вид функции. В предыдущих работах были рассмотрены различные виды зависимости между фенотипом и климатическими факторами [10–14]. Нами предложен более общий подход, который позволяет исследовать широкий круг нелинейных зависимостей между фенотипом и факторами. Аналитический вид управляющей функции определяется автоматически методом грамматической эволюции [15, 16] вместе с нахождением регрессионных коэффициентов методом LASSO [17] и набором предикторов путем стохастической минимизации отклонения решения модели от данных методом разностной эволюции [18]. При этом обучающая выборка составляла 75%, а ошибка рассчитывалась по неиспользованным при обучении данным.
В использованном подходе каждая модель характеризуется вектором из 19 чисел, 15 из которых описывают слова в грамматике, а 4, включая свободный член, являются коэффициентами построенной методом LASSO регрессионной модели. Таким образом, мы можем сравнить модели, сравнивая такие векторы.
Сравнение вкладов в модель различных факторов оценивается как отношение разницы сумм квадратов остатков без учета этого фактора и полной модели к сумме квадратов остатков без учета этого фактора.
Зависимость времени цветения от климатических факторов. В табл. 2 представлены значения коэффициента корреляции Пирсона между временем цветения и климатическими факторами. Статистически значимых зависимостей не обнаружено, все p-значения оказались значительно больше 0.05.
Таблица 2.
Корреляции климатических факторов со временем цветения
| Символ | Описание | Турция | Эфиопия |
|---|---|---|---|
| T mean | Средняя годовая температура | 0.104 (р = 0.462) | −0.236 (р = 0.078) |
| ∆T mean | Средний разброс температуры по месяцам | −0.049 (р = 0.731) | 0.026 (р = 0.850) |
| ∆T | Годовой разброс температуры | −0.061 (р = 0.665) | −0.077 (р = 0.568) |
| T sd | Сезонность температуры (100 × стандартное отклонение) | −0.043 (р = 0.761) | −0.192 (р = 0.151) |
| P ann | Годовые осадки | −0.004 (р = 0.975) | 0.172 (р = 0.201) |
| P sd | Сезонность осадков (100 × стандартное отклонение) | 0.035 (р = 0.708) | −0.039 (р = 0.771) |
Таким образом, зависимость времени цветения от климатических факторов не может иметь простой вид.
Адаптация модели к данным. Была проведена серия численных экспериментов. Поскольку метод минимизации целевой функции является стохастическим, требуется проводить много запусков. Путем перебора было установлено, что лучшими значениями является N = 3 функций длиной L = 15. На последнем этапе было проведено по 20 запусков для данных староместных сортов, собранных в Турции и Эфиопии. На рис. 1 представлен типичный график сходимости процесса минимизации.
Староместные сорта, собранные в Эфиопии. Среди полученных результатов для староместных сортов, собранных в Эфиопии, была выбрана модель (2):
(2)
$\begin{gathered} {{F}_{{\text{E}}}} = {\text{26}}{\text{.687}} + {\text{0}}{\text{.001}}\Delta {{T}^{{{\text{mean}}}}} - \\ - \,\,{\text{0}}{\text{.013}}{{T}^{{{\text{mean}}}}} - {\text{0}}{\text{.001}}{{T}^{{{\text{sd}}}}} + \\ + \,\,\frac{{{\text{162}}{\text{.879}}\left( {{{P}^{{{\text{sd}}}}} - {\text{95}}{\text{.842}}} \right)}}{{\left( {{{T}^{{{\text{sd}}}}} - {\text{1172}}{\text{.474}}} \right)\left( {{{T}^{{{\text{mean}}}}} - {\text{219}}{\text{.798}}} \right)}}, \\ \end{gathered} $Староместные сорта, собранные в Турции. Среди полученных результатов для староместных сортов, собранных в Турции, была выбрана модель (3):
(3)
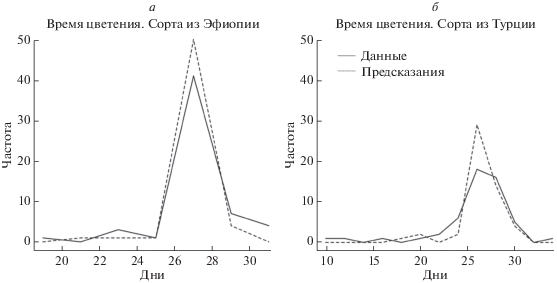
$\begin{gathered} {{F}_{{\text{T}}}} = {\text{29}}{\text{.602}} - {\text{0}}{\text{.004}}{{P}^{{{\text{ann}}}}} + \\ + \,\,\frac{{{\text{20}}{\text{.667}}}}{{{{P}^{{{\text{sd}}}}} - {\text{61}}{\text{.397}}}} + \frac{{{\text{10}}{\text{.449}}}}{{{\Delta }{{T}^{{{\text{mean}}}}} - {\text{137}}{\text{.866}}}}, \\ \end{gathered} $Сравнение моделей. На рис. 2 представлено сравнение полученных предсказаний с данными для сортов из Эфиопии (рис. 2,а) и Турции (рис. 2,б). Коэффициент корреляции Пирсона между векторами, описывающими функции FE и FT, составил 0.13 и оказался статистически незначим – p‑value = 0.38 > 0.05, коэффициент корреляции Спирмана составил 0.15 и незначим, p-value = 0.29 > > 0.05. Вклад температуры составил в модель для Турции 60%, для Эфиопии – 48%. Вклад осадков – 86 и 89%, для Турции и Эфиопии соответственно.
Рис. 2.
Гистограммы времени цветения. Линии соединяют точки, ординаты которых соответствуют количеству измерений, попавших в интервал, центр которого совпадает с абсциссой этой точки. Предсказанное время цветения хорошо согласуется с данными.
С помощью построенных нелинейных регрессионных моделей показано различие зависимостей времени цветения от климатических факторов для староместных сортов, собранных в Эфиопии и Турции.
Особенностью модели для староместных сортов, собранных в Эфиопии, является наличие зависимости от сложной функции одновременно осадков и температуры. Вклад температуры в модель для сортов из Эфиопии слабее, чем для сортов из Турции, что может являться следствием того, что в Эфиопии среднегодовые температуры относительно постоянны и способность реагировать на их изменения была утрачена. Вклад осадков в модель составляет более 80%, хотя для сортов из Эфиопии на 3% сильнее, чем для Турции, что может объясняться чувствительностью сортов к влаге.
Работа выполнена в рамках проекта РНФ 16-16-00007.
Вычисления проводились в СКЦ “Политехнический” ФГАОУ ВО СПбПУ.
Авторы заявляют, что у них нет конфликта интересов.
Настоящая статья не содержит каких-либо исследований с использованием в качестве объекта животных.
Настоящая статья не содержит каких-либо исследований с участием в качестве объекта людей.
Список литературы
Berger J., Milroy S., Turner N. et al. Chickpea evolution has selected for contrasting phenological mechanisms among different habitats // Euphytica. 2011. V. 180. P. 1–15. https://doi.org/10.1007/s10681-011-0391-4
Abbo S., Berge, J., Turner N. Evolution of cultivated chickpea: Four bottlenecks limit diversity and constrain adaptation // Functional Plant Biol. 2003. V. 30. P. 1081–1087. https://doi.org/10.1071/FP03084
Kumar J., Abbo S. Genetics of flowering time in chickpea and its bearing on productivity in the semi-arid environments // Adv. Agron. 2001. V. 72. P. 107–138. https://doi.org/10.1038/35098564
Pedersen P., Boote K.J., Jones J.W., Lauer J.G. Modifying the CROPGRO-Soybean model to improve predictions for the upper midwest // Agronomy J. 2004. V. 96. P. 556–564. https://doi.org/10.2134/agronj2004.0556
Setiyono T.D., Weiss A., Specht J. et al. Understanding and modeling the effect of temperature and daylength on soybean phenology under high-yield conditions // Field Crops Res. 2007. V. 100. P. 257–271. https://doi.org/10.1016/j.fcr.2006.07.011
Major D.J., Johnson D.R., Tanner J.W., Anderson I.C. Effects of daylength and temperature on soybean development // Crop Sci. 1975. V. 15. P. 174–179. https://doi.org/10.2135/cropsci1975.0011183X001500020009x
Soltani A., Hammer G.L., Torabi B. et al. Modeling chickpea growth and development: Phenological development // Field Crops Res. 2006 V. 99. P. 1–13. https://doi.org/10.1016/j.fcr.2006.02.005
Vadez V., Soltani A., Sinclair T.R. Modelling possible benefits of root related traits to enhance terminal drought adaptation of chickpea // Field Crops Res. 2012. V. 137. P. 108–115. https://doi.org/10.1016/j.fcr.2012.07.022
Козлов К.Н., Новикова Л.Ю., Cефеpова И.В., Cамcонова М.Г. Математичеcкая модель влияния климатичеcкиx фактоpов на pазвитие cои // Биофизика. 2018. Т. 63. Вып. 1. С. 175–176.
Hammer G.L., Vaderlip R.L., Gibson G. et al. Genotype-by-environment interaction in grain sorghum. II. Effects of temperature and photoperiod on ontogeny // Crop Sci. 1989. V. 29. P. 376–384. https://doi.org/10.2135/cropsci1989.0011183X002900020029x
Horie T. Crop ontogeny and development // Physiology and Determination of Crop Yield / Eds Boote K.J., Bennett J.M., Sinclair T.R., Paulsen G.M. Madison, 1994. P. 153–180.
Piper E.L., Boote K.J., Jones J.W., Grimm S.S. Comparison of two phenology models for predicting flowering and maturity date of soybean // Crop Sci. 1996. V. 36. P. 1606–1614. https://doi.org/10.2135/cropsci1996.0011183X003600060033x
Yin X., Kropff M.J., Horie T. et al. A model for photothermal responses of flowering in rice. I. Model description and parameterization // Field Crops Res. 1997. V. 51. P. 189–200. https://doi.org/10.1016/S0378-4290(96)03456-9
Robertson M.J., Watkinson A.R., Kirkegaard J.A. et al. Environmental and genotypic control of time to flowering in canola and Indian mustard // Australian J. Agricult. Res. 2002. V. 53. P. 793–809. https://doi.org/10.1071/ AR01182
O’Neill M., Ryan C. Grammatical evolution // IEEE Transactions on Evolutionary Computation. 2001. V. 5. P. 349–358. https://doi.org/10.1109/4235.942529
Noorian F., de Silva A.M., Leong P.H.W. gramEvol: Grammatical Evolution in R // J. Statist. Software. 2016. V. 71. P. 1–26. https://doi.org/10.18637/jss.v071.i01
Tibshirani R. Regression shrinkage and selection via the LASSO // J. Royal Statistical Soc. Ser. B. 1996. V. 58. P. 267–288.
Kozlov K., Samsonov A.M., Samsonova M. A software for parameter optimization with Differential Evolution Entirely Parallel method // Peer J. Computer Sci. 2016. V. 2. P. E74 (2016). https://doi.org/10.7717/peerj-cs.74
Дополнительные материалы отсутствуют.