

Генетика, 2020, T. 56, № 10, стр. 1184-1188
В-хромосомы восточноазиатских мышей (Apodemus peninsulae Thomas, 1907 (Rodentia, Muridae)) на восточных склонах Цинхай-Тибетского плато (КНР)
Ю. М. Борисов 1, И. А. Жигарев 2, *, Б. И. Шефтель 1, **
1 Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
119071 Москва, Россия
2 Московский педагогический государственный университет
129164 Москва, Россия
* E-mail: i.zhigarev@gmail.com
** E-mail: borissheftel@yahoo.com
Поступила в редакцию 10.11.2019
После доработки 12.12.2019
Принята к публикации 14.01.2020
Аннотация
Впервые изучены хромосомные наборы восьми особей Apodemus peninsulae, отловленных в провинции Ганьсу (заповедник Лианьхуашань: Lianhuashan Natural Reserve) в 2012 г. Хромосомные наборы мышей содержали от 55 до 62 хромосом и включали 48 А-хромосом и 7–14 добавочных В-хромосом разной морфологии.
Добавочные хромосомы (В-хромосомы) к настоящему времени открыты у многих видов не только животных, но и растений и грибов. У млекопитающих они отмечены у 55 видов. Большая их часть принадлежит к отряду грызунов (Rodentia). Например, из 20 видов мышей рода Apodemus дополнительные В-хромосомы выявлены у шести [1]. Одним из них является восточноазиатская, или корейская мышь (A. peninsulae). К настоящему времени хорошо изучен кариотип представителей этого вида из многих районов на севере ареала, в основном в пределах России, а также Японии, Кореи и Монголии. Практически у всех исследованных к настоящему времени A. peninsulae в индивидуальных кариотипах отмечены добавочные хромосомы, исключения редки [2]. Они имеют разные формы и количество: их число может доходить до 30 (в результате эти особи имеют в кариотипе 78 хромосом). Известна единственная популяция, обитающая на о-ве Сахалин, зверьки которой не имеют в кариотипе B-хромосом [3], однако в другой островной популяции – на о-ве Хоккайдо они имеются [4]. Наибольшее число В-хромосом выявлено у мышей, обитающих в Сибири [2, 3, 5–7].
Цитогенетики имеют особый интерес к этому виду вследствие его кариологической уникальности, поскольку он характеризуется некоторыми редкими особенностями. К ним следует отнести наличие самого высокого и разнообразного среди млекопитающих числа В-хромосом (оно варьирует от 0 до 30); резкое морфологическое различие между А-хромосомами (одноплечими) и В-хромосомами (чаще двуплечими); наличие в системе В-хромосом этого вида разнообразных размерно-морфологических вариантов (по размерам – от точечных до крупных, по морфологии – от микрохромосом, с неясным положением центромеры до двуплечих макро-В-хромосом); существование трех типов изменчивости по В-хромосомам: межпопуляционной, межиндивидуальной и внутрииндивидуальной (мозаицизм). Эти положения обусловливают перспективность использования восточноазиатских мышей в качестве модельного объекта при исследовании феномена В-хромосом в кариотипах млекопитающих. Это поможет внести ясность в понимание природы, роли и происхождения этих загадочных элементов геномов эукариот [2, 8, 9 и др.].
Восточноазиатская мышь распространена в Сибири от Алтая до Приморья, на северо-востоке Казахстана, в Северной Монголии, в Корее, на островах Сахалин и Хоккайдо, в Северо-Восточном и Центральном Китае до Цинхай-Тибетского плато на западе и провинции Юньнань на юге (рис. 1). Особый интерес представляют мыши, обитающие на восточных склонах Цинхай-Тибетского плато (КНР). Здесь сохранилось большое количество узкоареальных эндемичных видов мелких млекопитающих, населяющих уникальные реликтовые местообитания [10]. Примером подобных местообитаний служат хвойные леса и сопутствующие им растительные формации восточных склонов, исчезнувшие за пределами данного изолята. Такие растительные сообщества занимают юго-запад провинции Ганьсу, запад провинции Сычуань и северо-запад провинции Юньнань. Важная особенность этих горных хвойных лесов состоит в том, что они расположены между 30° и 35° с.ш. и изолированы степями и пустынями от схожих с ними северных таежных лесов. Существует точка зрения, что таежные экосистемы сформировались в горах Центральной Азии и лишь потом проникли на север, где заняли огромные пространства в Евразии и Северной Америке [11]. Поэтому мы предположили, что здесь могут существовать восточноазиатские мыши с уникальными сочетаниями классов В-хромосом.
Рис. 1.
Ареал A. peninsulae и пункты отлова мышей на территории Китайской Народной Республики: 1 – в провинции Ганьсу; 2 – в провинции Шэньси; 3 – в провинции Шаньдун.
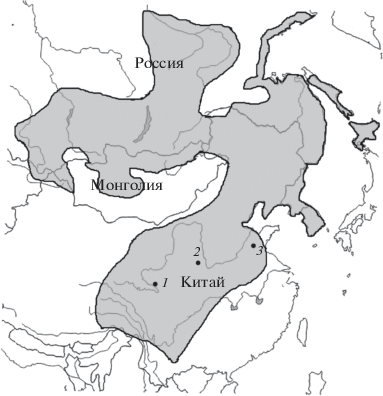
Данные о числе и морфотипах В-хромосом восточноазиатских мышей с территории Китая практически отсутствуют. Известна единственная работа [12], в которой приведены кариотипические материалы по мышам из двух провинций Центрального Китая (рис. 1, пункты 2 и 3).
Цель настоящей работы – анализ кариотипических особенностей и, в первую очередь, системы добавочных В-хромосом A. peninsulae, обитающих на восточных склонах Цинхай-Тибетского плато (Китай).
МАТЕРИАЛЫ И МЕТОДЫ
Отлов мелких млекопитающих проводили в окрестностях природного резервата Лианьхуашань в 2012 г. (Lianhuashan Natural Reserve), провинции Ганьсу (КНР) (рис. 1, пункт 1). Отловы были проведены в различных биотопах на трех высотных уровнях: 1) на высоте 2400 м над ур. м. на горных склонах c ксероморфными кустарниками, чередующимися с небольшими участками возделываемых полей; 2) в хвойных лесах и на пойменном лугу в долине небольшой речки на высоте 2850 м над ур. м.; 3) в зарослях кустарников на скальных склонах альпийского пояса на высоте 3100 м над ур. м. Подробно природные условия резервата описаны в монографии [13]. Всего кариотипировано восемь особей A. peninsulae. Восточноазиатские мыши ловились преимущественно в антропогенно-трансформированных биотопах.
Препараты изготовляли из клеток селезенки с применением технических приемов предложенных Е.Ю. Крысановым с коллегами [14].
Для характеристики кариотипа каждого животного использовали анализ более чем 20 метафазных пластинок. В настоящей работе нами применялась система классификации В-хромосом и формула их цифрового кодирования по Ю.М. Борисову [5, 6, 15]. В формуле цифрового кодирования выделено пять классов В-хромосом по их размерам и морфологическим характеристикам. Согласно этой формуле первая цифра – это В-хромосомы I класса: двуплечие, крупных размеров, равные по размерам 1–8 парам А-хромосом; вторая цифра – это В-хромосомы II класса: двуплечие, средних размеров, равные 9–16 парам А-хромосом; третья – это В-хромосомы III класса: двуплечие, мелкие, равные 17–23 парам А-хромосом; четвертая цифра – это составляющие IV класс В-акроцентрики, мелкие, равные 17–23 парам А-хромосом; наконец, пятая цифра – это В-хромосомы V класса, или микро В-хромосомы, с неясным положением центромеры, в несколько раз мельче самых мелких А-хромосом набора.
Для оценки общего количества хроматина добавочных хромосом мы использовали индекс условной массы В-хромосом кариотипа особи (индекс mВ), предложенный Г.В. Рослик и И.В. Картавцевой [16].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
У всех восьми особей A. peninsulae был определен основной стабильный набор А-хромосом данного вида. Он представлен 23 парами акроцентрических аутосом и двумя половыми хромосомами: крупной акроцентрической Х-хромосомой и мелкой акроцентрической Y-хромосомой (рис. 2). Вариабельная часть кариотипа у этих мышей представлена 7–14 дополнительными В-хромосомами. Таким образом, кариотипы исследованных мышей имеют от 55 до 62 хромосом. Спектр изменчивости вариантов системы В-хромосом восьми мышей A. peninsulae, отловленных в провинции Ганьсу (заповедник Лианьхуашань), показан с помощью формул цифрового кодирования в табл. 1.
Рис. 2.
Основной А-набор (48 акроцентрических хромосом) (а) и В-хромосомы: одна средняя метацентрическая (б), одна мелкая метацентрическая (в) и девять микро В-хромосом (г) самца A. peninsulae (Китай, юг провинции Ганьсу, заповедник Лианьхуашань в 2012 г.).

Таблица 1.
Популяционные варианты системы В-хромосом A. peninsulae в Китае (юг провинции Ганьсу, заповедник Лианьхуашань) в 2012 г.
| № | Пол | Система В-хромосом | Количество хромосом | Формула цифрового кодирования Bs | |||||
|---|---|---|---|---|---|---|---|---|---|
| (2n = 48)+Bs | Bs | X | X | x | v | · | |||
| 1 | ♀ | x … … | 55 | 7 | 0 | 0 | 1 | 0 | 6 |
| 2 | ♀ | x … … | 55 | 7 | 0 | 0 | 1 | 0 | 6 |
| 3 | ♂ | … … … | 57 | 9 | 0 | 0 | 0 | 0 | 9 |
| 4 | ♂ | x … … … | 58 | 10 | 0 | 0 | 1 | 0 | 9 |
| 5 | ♂ | X x … … … | 59 | 11 | 0 | 1 | 1 | 0 | 9 |
| 6 | ♂ | x … … … .. | 60 | 12 | 0 | 0 | 1 | 0 | 11 |
| 7 | ♀ | X X x … … … | 60 | 12 | 0 | 2 | 1 | 0 | 9 |
| 8 | ♂ | X X x … … … .. | 62 | 14 | 0 | 2 | 1 | 0 | 11 |
| ♀: 3, ♂: 5 | Сумма | 82 | 0 | 5 | 7 | 0 | 70 | ||
| Доля и ошибка доли, % | 100 | 0 | 6.1 ± 2.7 | 8.5 ± 3.1 | 0 | 85.4 ± 3.9 | |||
| Популяционный индекс xB, с ошибкой | 10.25 ± 0.88 | 0 | 0.63 ± 0.32 | 0.88 ± 0.13 | 0 | 8.75 ± 0.67 | |||
| Коэффициент вариации, % | 24 | 0 | 147 | 40 | 0 | 22 | |||
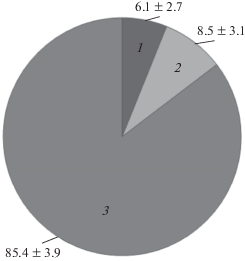
В кариотипах всех исследуемых зверьков имелись точечные микрохромосомы, их число варьировало у разных особей от 6 до 11. Практически все особи (87.5%) имели по одной мелкой метацентрической хромосоме. При этом у исследуемых мышей отсутствовали крупные метацентрики (для сравнения – в северных частях ареала они присутствуют у 28% особей [2]), а также акроцентрики (в северных частях ареала они присутствуют у 12% особей). Таким образом основная изменчивость у исследуемых особей касалась вариации количества средних метацентрических (изменялись от 0 до 2) и точечных (изменялись от 6 до 11) добавочных хромосом. Долевое соотношение добавочных хромосом отображено на рис. 3. Среднее число В-хромосом – популяционный индекс (хВ) у восьми особей составил 10.25 ± 0.88. В целом эти особенности соответствуют характеру изменчивости в северных (сибирских) популяциях.
Рис. 3.
Доли классов (в %) В-хромосом A. peninsulae в Китае (юг провинции Ганьсу, заповедник Лианьхуашань) в 2012 г. 1 – средние метацентрики; 2 – мелкие метацентрики; 3 – микро В-хромосомы.
Для оценки суммарной “массы” добавочных хромосом в кариотипе A. peninsulae можно использовать относительный индекс условной массы В-хромосом (индекс mB). Он позволяет сравнить разные выборки по объему добавочного хроматина в клетках мышей разных популяций. Этот индекс показывает умеренные значения для мышей окрестностей заповедника Лианьхуашань (12.4 ± 1.5). Подобные показания характерны для многих районов Сибири и Забайкалья [2]. Наименьшее значение индекс mB приобретает в Приморье (4.1 ± 0.3), а наибольшее – в Кемеровской области и на юге Красноярского края (индексы mB > 18).
Кроме нашего материала, с территории Китая известны кариотипы 12 особей A. peninsulae из двух районов в центральной части страны [12]: семь особей из провинции Шаньдун (Tai, Shandong Prov. (рис. 1, пункт 3)) и пять особей из провинции Шэньси (Qinling, Shaanxi Prov. (рис. 1, пункт 2)). У зверьков из провинции Шэньси число В-хромосом варьировало от 0 до 1. Все они были макро В-хромосомы, микро В-хромосомы у них не выявлены. У зверьков из провинции Шаньдун число В-хромосом варьировало от 5 до 14. Одна из них была метацентрическая макро В‑хромосома, а число микро В-хромосом изменялось от 4 до 13.
Таким образом, мы установили, что, несмотря на реликтовые местообитания восточных склонов Цинхай-Тибетского плато, восточноазиатские мыши, обитающие в этих местах, не имеют уникальных сочетаний различных классов В-хромосом. Такое положение, по всей видимости, объясняется тем, что данный вид не является коренным компонентом реликтовых экосистем, а проникает сюда лишь по нарушенным местообитаниям.
В целом по соотношениям различных классов хромосом, особенно по большому количеству микро В-хромосом, кариотипы зверьков из данного региона больше всего напоминают кариотипы зверьков с южного берега оз. Байкал (Бурятия) [17]. Напротив, эти кариотипы существенно отличаются от кариотипов зверьков, собранных в соседней китайской провинции Шэньси.
Важно заключить, что добавочные хромосомы в кариотипах представителей A. peninsulae, по крайней мере континентальных популяций, присутствуют практически всегда и являются скорее обязательным, чем нежелательным компонентом. Однако вопрос адаптивности этого цитогенетического феномена остается открытым, что требует дальнейших исследований изменчивости добавочных хромосом в популяциях A. peninsulae.
Благодарим А.Ю. Александрову за помощь в приготовлении хромосомных препаратов восточноазиатских мышей. Благодарим научный, административный и технический персонал Национального резервата Лианхуашань за постоянную помощь в проведении исследований.
Работа выполнена при финансовой поддержке РФФИ-ГФЕН (11-04-91188).
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Vujosevic M., Blagojević J. B chromosomes in populations of mammals // Cytogenet. Genome Res. 2004. V. 106. № 2–4. P. 247–256. https://doi.org/10.1159/000079253
Borisov Y.M., Zhigarev I.A. B Chromosome system in the Korean field mouse Apodemus peninsulae Thomas, 1907 (Rodentia, Muridae) // Genes. 2018. V. 9. № 472. https://doi.org/10.3390/genes9100472
Kartavtseva I.V., Roslik G.V. A complex B chromosome system in the Korean field mouse, Apodemus peninsulae // Cytogenet. Genome Res. 2004. V. 106. № 2–4. P. 271–278. https://doi.org/10.1159/000079298
Hayata J. Chromosomal polymorphism caused by supernumerary chromosomes in field mouse, Apodemus giliacus // Chromosoma. 1973. V. 42. № 4. P. 403–414.
Борисов Ю.М. Географическая изменчивость вариантов системы добавочных хромосом у континентальных форм Apodemus peninsulae (Rodentia. Muridae) // Изв. СО АН СССР. Сер. биол. наук. 1980. № 15(330). С. 61–69.
Борисов Ю.М. Система В-хромосом восточноазиатской мыши как интегрирующий и дифференцирующий признак популяций // Докл. АН СССР. 1986. Т. 288. № 3. С. 720–724.
Борисов Ю.М., Афанасьев А.Г., Лебедев Т.Т., Бочкарев М.Н. Множество микро-В-хромосом в сибирской популяции мышей Apodemus peninsulae (2n = = 48 + 12 – 30 B-хромосом) // Генетика. 2010. Т. 46. № 6. С. 798–804.
Houben A., Jones N., Martins C., Trifonov V. Evolution, composition and regulation of supernumerary B chromosomes // Genes. 2019. V. 10. № 161.
Trifonov V.A., Perelman P.L., Kawada S.-I. et al. Complex structure of B-chromosomes in two mammalian species: Apodemus peninsulae (Rodentia) and Nyctereutes procyonoides (Carnivora) // Chromosome Res. 2002. V. 10. № 2. P. 109–116.
Шефтель Б.И., Банникова А.А., Фанг Ю. и др. Заметки по фауне, систематике и экологии мелких млекопитающих юга провинции Ганьсу (Китайская Народная Республика) // Зоол. журн. 2017. Т. 96. № 2. С. 232–248.
Толмачев А.И. К истории возникновения и развития темнохвойной тайги. М.; Л.: Изд-во АН СССР, 1954. 155 с.
Wang J., Zhao X., Qi H. et al. Karyotypes and B chromosomes of Apodemus peninsulae (Rodentia, Mammalia) // Acta Theriologica Sinica. 2000. V. 20. № 4. P. 289–296.
Sun Y.-H., Fang Y., Klaus S. et al. Nature of the Lianhuashan Natural Reserve. Liaoning Sci. and Technology Publ. House, 2008. 100 p.
Крысанов Е.Ю., Демидова Т.Б., Шефтель Б.И. Простой метод приготовления препаратов хромосом мелких млекопитающих // Зоол. журн. 2009. Т. 88. № 2. С. 234–238.
Борисов Ю.М. Цитогенетическая дифференциация популяций восточноазиатской мыши в Восточной Сибири // Генетика. 1990. Т. 26. № 10. С. 1828–1839.
Рослик Г.В., Картавцева И.В. Морфотипы B-хромосом Apodemus peninsulae (Rodentia) Дальнего Востока России // Цитология. 2012. Т. 54. № 1. С. 66–77.
Борисов Ю.М., Шефтель Б.И., Сафронова Л.Д., Александров Д.Ю. Устойчивость популяционных систем B-хромосом восточноазиатской мыши Apodemus peninsulae Прибайкалья и Северной Монголии // Генетика. 2012. Т. 48. № 10. С. 1190–1199.
Дополнительные материалы отсутствуют.