

Генетика, 2020, T. 56, № 7, стр. 826-833
Генофонд хакасов и шорцев по маркерам Y-хромосомы: общие компоненты и генетическая структура родов
В. Н. Харьков 1, *, Л. М. Новикова 1, О. В. Штыгашева 2, Ф. А. Лузина 3, И. Ю. Хитринская 1, В. Г. Волков 1, В. А. Степанов 1
1 Научно-исследовательский институт медицинской генетики, Томский национальный исследовательский медицинский центр Российской академии наук
634050 Томск, Россия
2 Хакасский государственный университет им. Н.Ф. Катанова
655017 Абакан, Россия
3 Научно-исследовательский институт комплексных проблем гигиены и профессиональных заболеваний Сибирского отделения Российской академии наук
654041 Новокузнецк, Россия
* E-mail: vladimir.kharkov@medgenetics.ru
Поступила в редакцию 13.07.2019
После доработки 12.12.2019
Принята к публикации 17.12.2019
Аннотация
Исследована генетическая структура хакасских и шорских популяций и родов (сеоков) по маркерам Y-хромосомы. Результаты анализа частот гаплогрупп и YSTR-гаплотипов свидетельствуют, что хакасские и шорские сеоки являются родственными объединениями, в большинстве случаем имеющими одного родоначальника по мужской линии. Показано, что генофонд хакасов и шорцев, а точнее часть, маркируемая гаплогруппами Y-хромосомы, структурирована, прежде всего, по родовому принципу. Для подавляющего большинства образцов показана тесная генетическая близость представителей одного сеока. Отдельные гаплогруппы Y-хромосомы в составе хакасского и шорского генофондов демонстрируют их генетическую близость с тувинцами, кетами, алтайцами, телеутами. Это позволяет говорить о единстве обобщенного генофонда коренного южносибирского населения и генетической преемственности популяций, проживающих на данной территории. Выявлены значительные различия между субэтническими группами, вошедшими в состав современных хакасов и шорцев, при этом оба народа формировались на базе общих генетических компонент и демонстрируют генетическое родство по различным родам.
Хакасы и шорцы – родственные тюркоязычные народы, составляющие большинство коренного населения Республики Хакасия и Кемеровской области. Хакасы подразделяются на четыре этнографические группы: сагайцы, качинцы, кызыльцы, койбалы [1]. Шорцы представлены двумя группами – южной горнотаежной и северной лесостепной. По данным Всероссийской переписи населения 2010 г., численность хакасов составила 72 959 человек, а шорцев – 12 888 человек [2]. Как и другие народы Южной Сибири, хакасы и шорцы характеризуются сложным этногенезом. Генофонд коренного населения этого региона складывался за счет длительного и многоэтапного смешения большого числа локальных генофондов различных племен европеоидного и монголоидного происхождения. Смешение многочисленных тюркских, монгольских, енисейских, уральских групп на основе генетического субстрата древних европеоидных племен и таежных монголоидов сформировало в результате пеструю картину генетического разнообразия коренного населения [3].
Ранее по маркерам Y-хромосомы уже были охарактеризованы генетическая структура и субэтническая подразделенность хакасов [4] и шорцев [5]. Показано, что хакасский и шорский генофонды структурированы в соответствии с территориально-субэтническим принципом. Был выявлен сложный и многокомпонентный состав их генофонда, который содержит в себе практически все основные компоненты, характерные для других коренных этносов Южной Сибири, при этом обладая уникальным соотношением частот гаплогрупп и структуры YSTR-гаплотипов. Для хакасов были показаны сильные различия между качинцами и сагайцами по составу гаплогрупп, уровню генетического разнообразия и спектру гаплотипов, показан высокий уровень генетической дифференциации исследованных выборок. Cубэтнические группы лесостепных и горно-таежных шорцев также достоверно различаются между собой.
Но ограниченный набор известных информативных SNP-маркеров и относительно небольшое число прогенотипированных YSTR на тот момент не позволяли точно определить близость генетических компонент, маркируемых различными гаплогруппами. В последние годы обнаружено огромное количество информативных SNP на Y-хромосоме и существенно расширена база YSTR-гаплотипов, что дает возможность анализировать генофонд этих популяций на совершенно другом уровне.
Особенностью этнического состава большинства южносибирских народов является наличие родов (сеоков), где счет родства ведется по мужской линии. Такая родовая структура характерна для шорцев, хакасов, северных и южных алтайцев, телеутов [1]. Это приводит нас к необходимости рассмотрения организации генофонда коренного народонаселения Южной Сибири помимо этнического и субэтнического уровня также и на уровне родов. В настоящее время точность определения гаплотипов и значительная детализация сублиний внутри гаплогрупп позволяют проверить, является ли сеок кровнородственным объединением, имеющим одного родоначальника по мужской линии или осознание такой общности является лишь условностью, и сеок является общностью людей, проживающих на одной территории, но не связанных генетическим родством по отцовским генеалогическим линиям. Установление родовой принадлежности у хакасов и шорцев в большинстве случаев можно однозначно сделать по фамилиям, так как практически все они приписаны этнографами к конкретному сеоку [6]. Даже если современные жители не относят себя к тому или иному роду, не помнят принадлежности своих предков к конкретному сеоку, более того в настоящее время осознают себя сагайцами, хотя многие поколения их прародителей считали себя бельтирами или бирюсинцами, фамилия человека служит указателем на его происхождение. Особенностью хакасской и шорской родовой системы является почти полное отсутствие однофамильцев. Это связано с разнообразием используемых имен, от которых и произошли фамилии. Родоначальники этих фамилий жили в основном в XVIII в. Каждая такая фамилия входит в состав конкретного сеока: харга, хасха, пилтир, хый, хобый, хыргыс и т.д.
Цель исследования – с помощью подробного анализа генетической структуры по маркерам Y‑хромосомы выявить общие компоненты генофонда хакасов и шорцев и установить межэтническое генетическое родство отдельных родоплеменных групп в составе этих этносов.
МАТЕРИАЛЫ И МЕТОДЫ
Материал исследования составили образцы ДНК неродственных между собой мужчин из различных локальных популяций хакасов (N = 292) и шорцев (N = 108). Забор первичного биологического материала (венозной крови) у доноров проводили с соблюдением процедуры письменного информированного согласия на проведение исследования. На каждого донора составлялась анкета с краткой родословной, указанием этнической принадлежности и мест рождения предков. В исследование включены только образцы ДНК доноров-мужчин, по результатам анкетирования отрицавших факт метисации по отцовской линии с представителями других этносов минимум в трех поколениях. Индивида относили к данной этнической группе на основании его собственной этнической идентификации, его родителей, места рождения и принадлежности к определенному сеоку. Выборка мужчин хакасов представляет три популяционные группы жителей территориально разобщенных районов: Аскизского (села Усть-Есь, Есино (улус Полтаков), Усть-Чуль и Кызлас, N = 160), Таштыпского (деревни Матур, Анчуль, Большая Сея и Бутрахты, N = 81) и Ширинского (села Малый Спирин и Топанов, N = 51). Материал получен в ходе совместных научно-практических медицинских экспедиций в 2007–2008 и 2016 гг. Среди всех мужчин, включенных в настоящее исследование, оказались представлены следующие сеоки (в алфавитном порядке): кичин, пилтыр, пурут, сагай, сайын, сарыг, сойыт, сор, соххы, табан, таяс, таяш, тиилек, том, том-сагай, туран, халар, хан, харга, хасха, хахпына, хобый, хызыл хая, хый, хыргыс, чити пуур, чыстар, ызыр.
Выборка шорцев представляет население пос. Усть-Кабырза (N = 45) в Таштагольском районе Кемеровской области, пос. Ортон (N = 32) Междуреченского городского округа, пос. Чувашка (N = 28) и г. Мыски (N = 3) Мысковского городского округа. Среди всех мужчин, включенных в данное исследование, оказались представлены следующие сеоки (в алфавитном порядке): ак-шор, кара-шор, карга, кобый, кызай, кый (кой), сары-шор, таеш, тарткын, чедибер, челей, шор-кызай.
Для изучения состава и структуры гаплогрупп Y-хромосомы в исследование были включены две системы генетических маркеров: диаллельных локусов, представленных SNP, и полиаллельных высоковариабельных микросателлитов (YSTR). С помощью 97 SNP-маркеров определяли принадлежность образцов к той или иной гаплогруппе. Классификация гаплогрупп дана в соответствии с предложенной Консорциумом по исследованию Y-хромосомы [7], с последующими изменениями [8, 9]. Анализ STR-гаплотипов внутри гаплогрупп проводили с использованием 45 микросателлитных маркеров нерекомбинирующей части Y-хромосомы (YSTR) (DYS19, 385a, 385b, 388, 389I, 389II, 390, 391, 392, 393, 426, 434, 435, 436, 437, 438, 439, 442, 444, 445, 448, 449, 456, 458, 460, 461, 481, 504, 505, 518, 525, 531, 533, 537, 552, 570, 576, 635, 643, YCAIIa, YCAIIb, GATA H4.1, Y-GATA-A10, GGAAT1B07). STR-маркеры генотипировали с помощью капиллярного электрофореза на генетическом анализаторе ABI Prism 3730. Генотипирование SNP-маркеров проводили с помощью ПЦР и последующего анализа фрагментов ДНК с помощью ПДРФ. STR-маркеры генотипировали с помощью капиллярного электрофореза на генетическом анализаторе ABI Prism 3730 и программного обеспечения GeneMapper. Экспериментальные исследования проведены на базе Центра коллективного пользования научно-исследовательским оборудованием “Медицинская геномика” (НИИ медицинской генетики Томского НИМЦ). Построение медианных сетей гаплотипов Y-хромосомы проводили с использованием программы Network v. 5.0.1.1 (Fluxus Technology Ltd.; www.fluxus-engineering.com) по методу медианных сетей Бандельта [10]. Оценку возраста генерации наблюдаемого разнообразия гаплотипов в гаплогруппах проводили методом ASD [11], на основании средних квадратичных отличий в числе повторов между всеми маркерами.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Генотипирование расширенного набора новых информативных SNP и YSTR позволило гораздо более точно охарактеризовать структуру генофонда хакасов и шорцев, выявить наличие отличающихся сублиний внутри различных гаплогрупп и оценить степень их генетической близости между представителями отдельных родов, субэтносов и народов. В данной работе мы сфокусировали внимание не на стандартном описании генетического разнообразия и дифференциации, а на анализе генетического родства между хакасами и шорцами на уровне родов и этнографических групп. В.Я. Бутанаев выделяет шесть таких групп, вошедших в состав хакасов и шорцев: качинцы, койбалы, сагайцы, бельтиры, бирюсинцы, кызыльцы [12]. В XIX в. основное деление происходило по принадлежности к административным родам. Перечисленные выше сеоки, к которым принадлежат протестированные люди, входили в состав различных административных родов. Ниже будет описана генетическая структура именно этих существовавших в прошлом этнографических групп, которые по различным данным вошли в состав современных субэтносов хакасов и шорцев. Рассмотрим подробнее полученные результаты.
Бельтиры
Эта группа хакасов является генетически гетерогенной. Cеоки пилтыр и чыстар представлены в основном образцами гаплогруппы R1a-Y62155. В других исследованных субэтнических выборках хакасов эта гаплогруппа отсутствует. У шорцев она также не выявлена, но с небольшой частотой встречается у тувинцев, которые очень близки к хакасам по гаплотипам (собственные неопубликованные данные). Единственный образец сеока кичин принадлежит сублинии R1a-Y43109. Все обследованные мужчины сеока табан относятся к гаплогруппе N1a2b2a2-VL67; практически все образцы сеока хахпына – к гаплогруппе Q1b1a3b-YP1102.
Это полностью соответствует данным Л.П. Потапова о том, что название “бельтиры” представляло собой, по-видимому, собирательное географическое наименование, означающее в переводе на русский язык “устьинцы” (т.е. живущие в устьях рек) [13]. В XVIII в. они представляли собой тюркоязычную группу, довольно смешанную в этническом отношении. Согласно распространенным легендам, бельтиры образовались преимущественно из выходцев из Тувы. Данные по генетике бельтир, таким образом, в целом подтверждают этнографические.
Образцы сеока хахпына очень близки по YSTR-гаплотипам с образцами Q1b1a3b-YP1102, найденными у тувинцев (собственные неопубликованные данные). Предполагается, что хахпына связаны с народом Капкапас, который в XIII в. населял истоки р. Енисей [1]; также имеется гипотеза, что в древности капкапасы были кетоязычными, так как хахпын по-кетски значит “кукушка” [13]. Гаплогруппа Q1b1a3b-YP1102 (ранее Q1a3) на территории Южной Сибири с максимальной частотой выявлена именно у кетов, селькупов и тувинцев, маркируя именно кетский генетический компонент [14, 15]. Все эти данные свидетельствуют о сложном, многокомпонентном происхождении бельтир, объединивших кыргызские и тувинские элементы.
Бирюсинцы
Разделяются на три группы сеоков по принадлежности к различным гаплогруппам. Отличительной особенностью сеоков хый и хобый является доминирование по частоте гаплогруппы N1a1a2-B499, что не наблюдается больше не в одном другом хакасском сеоке. Таким образом, эти два сеока объединяют в себе отдельный генетический компонент, не характерный для других хакасов. У шорцев к этой гаплогруппе относятся все мужчины сеоков кый и кобый. Родовое название кобый есть во всех известных списках шорцев. Сеок кобый в середине XIX в. составлял отдельную Кивийскую или Кобыйскую волость. При этом гаплотипы N1a1a2-B499 на медианной сети формируют два звездообразных кластера, отражающих достаточно сильный эффект основателя по данной гаплогруппе у бирюсинцев.
Имеются этнографические данные о переселении ясачных Кивинской волости в верховья Абакана к енисейским кыргызам [16]. После включения в 1822 г. этой территории в Сагайскую степную думу Минусинского округа постепенно происходило растворение абаканских кобыйцев в среде сагайцев-хакасов [6]. В настоящее время жители Таштыпского района из сеоков хый и хобый осознают себя частью хакасского народа [16].
Антрополог В.П. Алексеев, обследовавший несколько черепов из старых могил абаканских кобыйцев, пришел к выводу, что тюркизация последних могла происходить только путем культурных, а не брачных контактов, так как кобыйцы сохранили в чистоте уральский расовый тип, характерный для самодийских и уральских народов [17]. Связать наличие гаплогруппы N1a1a2-B499 в генофонде хакасов и шорцев с каким-то одним конкретным предковым субстратным элементом однозначно невозможно, но наиболее вероятно, что она маркирует именно самодийский этнический компонент.
Гаплогруппа N1a1a2-B499 имеет очень ограниченный ареал, и до сих пор обнаружена только у хакасов, шорцев и тувинцев [9] (ранее была описана нами как кластер II N1c1) [15]. Она очень рано отделилась от основного ствола N1a1 и как и многие другие редкие Y-хромосомные линии сохранилась с достаточно высокой частотой лишь в относительно изолированных горных популяциях. Вероятно, что исходным местом ее распространения была территория Тувы, откуда она распространилась в Горную Шорию и затем в Хакасию.
На медианной сети гаплотипов гаплогруппы N1a1a2-B499 (рис. 1) отчетливо видно, что, несмотря на наличие двух различных кластеров, между шорцами и хакасами наблюдается тесное генетическое родство. Общий возраст для всего массива гаплотипов, рассчитанный на основании генеалогического темпа мутирования в 0.0021 на локус на поколение, составил 2160 лет (SD = 564 года), для кластера доминирующего по частоте у шорцев 580 лет (SD = 146 лет). Вероятно, общий возраст несколько завышен, поскольку между двумя кластерами гаплотипов наблюдаются различия в несколько мутационных шагов по нескольким STR, что приводит к высоким значениям дисперсии.
Рис. 1.
Медианная сеть YSTR-гаплотипов гаплогруппы N1a1a2-B499. Белым цветом обозначены хакасы, серым – шорцы.
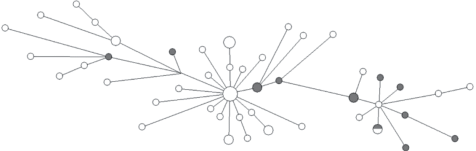
Представители бирюсинских хакасских сеоков туран Дальнекаргинского рода и хызыл хая Кызыльского рода относятся к сублинии R1a-Y43850. У шорцев к этой гаплогруппе, за единичными исключениями, относятся все мужчины сеоков тарткын, шор-кызай и кара-шор. Распространение различных сублиний гаплогруппы R1a на территории Кузнецкого Алатау и Минусинской котловины связано вероятнее всего с тюрками и енисейскими кыргызами. Сеок кызай отмечен всеми исследователями конца XIX–начала XX в. и обитал в это время в среднем течении Мрассу. Из исторических документов следует, что еще в XVIII в. часть сеока кызай переселилась в “Киргизскую землицу” в верховьях Абакана [18]. К настоящему времени эти абаканские потомки сеока кызай (хызыл-хая) полностью слились с сагайцами и считают себя частью хакасского этноса [16].
Мужчины из хакасских сеоков таяс и халар Изушерского рода, кичин и таяш Мрасско-Изушерского рода, сеоков сор Карачерского рода и большинство образцов шорского сеока челей относятся к сублинии R1a-Y43109. Три мужчины-хакаса с различными фамилиями из сеока харга и все шорцы сеока карга принадлежат к сублинии R1a-YP1509. Сеоки сайын и харга Ближнекаргинского рода практически полностью состоят их носителей гаплогруппы N1a2b2a2-VL67. К этой же линии принадлежат девять мужчин шорского сеока челей.
Наличие одинаковых гаплогрупп и близость гаплотипов показывают, что административные рода, на которые делились бирюсинцы в XIX в., представляют собой объединение родственных друг другу сеоков. Наиболее близкими между собой оказались: Кивинский и Кийский рода (N1a1a2-B499), Изушерский, Мрасско-Изушерский и Карачерский рода (R1a-Y43109), сеоки харга и сайын Ближнекаргинского рода бирюсинцев (N1a2b2a2-VL67), которые согласно этнографическим и историческим данным относятся к шорским родам. Уже в названии родов Изушерский, Мрасско-Изушерский и Карачерский содержится указание на их связь с шорцами (окончание -чер, -шер). Этнографы относят эпонимы шор и сайын к самодийским по происхождению [16]. Гаплогруппа N1a2b2a2-VL67 в генофондах хакасов и шорцев маркирует, судя по всему, именно самодийский субстрат.
Бирюсинцы, таким образом, также являются генетически гетерогенной группой, в составе которой присутствуют четыре различных компонента, маркирующих самодийский и тюркский субстрат. Доказано, что большинство бирюсинских хакасских и шорских сеоков с общими или близкими названиями имеют общее происхождение, фактически составляя общие патрилинейные группы, административно и территориально разделенные в настоящее время и вошедшие в состав современных шорцев и хакасов.
Сагайцы
Представители хакасского сеока туран относятся к сублинии R1a-Y43850. Сеок чити-пуур относится к сублинии R1a-Y43109. Мужчины сеоков сагай, сарыг, ичеге относятся к гаплогруппе N1a2b2a2-VL67. В состав сагайцев и бельтыров в основном вошли остатки бывших кыргызских аймаков Алтырского улуса. Кроме того, к ним примкнули небольшие группы переселенцев с территорий современных Алтая, Тувы, Шории и даже Джунгарии [1].
Полученные результаты также хорошо согласуются с данными антропологических исследований о большей доле европеоидной примеси именно у сагайцев, по сравнению с качинцами. Наличие гаплогрупп R1a-Y43850 и R1a-Y43109 у хакасов обусловлено европеоидным субстратом, на основе которого формировался данный этнос. В археологическом отношении, на наиболее позднем этапе развития, носителям этого компонента соответствует тагарская культура, представленная на территории Хакасии многочисленными курганами.
Интересно, что азиатский вариант гаплогруппы R1a, широко распространенный в популяциях Южной Сибири, оказался у хакасов и шорцев представлен несколькими различными сублиниями, которые разделились очень давно. При этом не в одной из этих субгаплогрупп не наблюдается звездообразной филогении гаплотипов, пониженного разнообразия и следов эффекта основателя. Судя по всему, это свидетельствует о значительном размере эффективной численности популяций древних европеоидов и тюрков, которые привнесли эти компоненты в генофонд современных хакасов и шорцев.
Качинцы
Этот хакасский субэтнос ранее подразделялся на четыре рода: Кубанов (сеок соххы), Мунгатов (сеок ызыр), Татаров (сеок пурут) и Шалошин (сеоки пурут и хасха). Яркой отличительной особенностью качинцев является то, что все сеоки полностью состоят из носителей гаплогруппы N1a2b2a2-VL67 с довольно специфическим спектром YSTR-гаплотипов (рис. 2). Общий возраст для всего массива гаплотипов составил 1400 лет (SD = 464 года), для кластера сеока хасха – 487 лет (SD = 153 года), для кластера сеока ызыр – 501 год (SD = 203 года), для кластера сеока соххы – 585 лет (SD = 215 лет). Таким образом, примерно совпадает не только возраст начала экспансии численности различных сеоков качинцев, но и бирюсинцев.
Рис. 2.
Медианная сеть YSTR-гаплотипов гаплогруппы N1a2b2a2-VL67 у качинцев. Белым цветом обозначен сеок соххы, серым – ызыр, черным – хасха, в клетку – пурут.
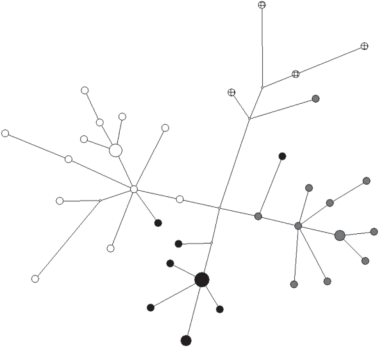
Результаты сравнительного анализа убедительно свидетельствуют о близком родстве между всеми четырьмя качинскими сеоками. За единичными исключениями, каждый сеок имеет свой специфичный кластер гаплотипов, равноудаленный от других. При этом все сеоки значительно генетически удалены от сагайцев, бельтиров, бирюсинцев и шорцев. Несмотря на то что N1a2b2a2-VL67 является самой распространенной линией у хакасов и, несомненно, представляет для них общий генетический субстрат, она очень сильно различается по своей внутренней гаплотипической структуре у разных субэтносов. Это свидетельствует о значительной генетической дифференциации между ними. Скорее всего, это объясняется отсутствием сколько-нибудь значительного обмена генами на протяжении последних нескольких сотен лет, и можно утверждать, что основным фактором популяционной динамики, сыгравшим роль в этом процессе, была генетическая изоляция качинцев от их более южных соседей.
Название качинцы, согласно свидетельству Г.Ф. Миллера, появилось в XVII в. и связано с названием сеока хасха [19]. Русская форма качи затем была перенята хакасским населением как хааш (хаас), но уже в значении общего обозначения скотоводов Качинской степной думы. Историческое название “хааш” характерно для этнических образований оленеводов Восточных Саян. Например, “хааш” – один из основных сеоков тофаларов, “хааш” – тувинское название бурятских сойотов, “хааш” – дархатское обозначение тувинцев – тоджинцев, “хара хааш” – тувинское наименование тофаларов. По всей видимости, термин хааш (хаас) восходит к самодийскому хас – мужчина [20]. В.Я. Бутанаев [19] замечает, что истинный ареал этого этнонима подтверждает правоту многих ученых, полагающих, что он связан с самодийцами, но добавляет, что самодийцы не имеют этногенетических связей ни с тюркоязычными качинцами, ни с территорией Хакасии. Но данные генетики убедительно свидетельствуют об обратном. Все качинские сеоки по мужской линии имеют прямую генетическую связь с народами самодийской языковой группы. Таким образом, предполагаемая этнографами связь этнических групп, название которых восходит к общесамодийскому обозначению человека – кас, каш, хааш, подтверждается данными генетики. Гаплогруппа N1a2b2a2-VL67 наиболее распространена именно у современных самодийских народов: ненцев, энцев, нганасан, а также у тувинцев, для которых самодийский компонент является частью их генофонда [9, 15].
Качинцы, таким образом, выделяются по сравнению с другими субэтническими группами хакасов наибольшей долей самодийского по происхождению компонента в их генофонде. Это хорошо согласуется с антропологическими отличиями качинцев от других хакасских групп и преобладанием у них комплекса признаков, характерного для южносибирского типа [21].
Полученные результаты не противоречат общепринятым версиям этногенеза хакасов и шорцев, но заставляют по-новому взглянуть на этот процесс. По данным антропологов и этнографов считается, что хакасы и шорцы представляют собой тюркизированных потомков угорских, самодийских и кетоязычных аборигенов северной, таежной части Алтае-Саянского нагорья. При этом, отличаясь меньшей монголоидностью основных антропологических признаков, шорцы обнаруживают общность антропологического типа с хантами и манси [21–23]. Отдельные элементы в составе хакасского и шорского генофондов по гаплогруппам Y‑хромосомы демонстрируют их генетическую близость с тувинцами, кетами, алтайцами, телеутами. Это позволяет говорить о единстве обобщенного генофонда коренного южносибирского населения и генетической преемственности популяций, проживающих на данной территории. Выявлены сильные различия между субэтническими группами, вошедшими в состав современных хакасов и шорцев по составу гаплогрупп, маркирующих различные генетические компоненты в составе их генофонда.
Получено подтверждение того, что хакасский и шорский этносы действительно формировались путем территориально-административного разделения генетически монофилетичных родственных родовых групп (по мужской линии). Результаты проведенного анализа доказывают, что хакасские и шорские сеоки являются, прежде всего, объединением родственников по отцовской линии. Конечно, не для всех сеоков приписанные к ним образцы принадлежали к одной гаплогруппе и одному кластеру гаплотипов, но это явилось скорее исключением из общего правила. Для подавляющего большинства образцов показана тесная генетическая близость представителей одного сеока. Полученные результаты позволяют совершенно по-новому взглянуть на историю формирования хакасов и шорцев, с одной стороны, на самом современном уровне генотипирования Y-хромосомных маркеров, с другой стороны, полностью учитывая родовую структуру народа, анализируя межэтнические взаимосвязи и родовые различия. Именно этот уровень организации генофонда этноса как единой системы является наиболее точно характеризующим его популяционную структуру. Полученные результаты генетического анализа позволяют нам сделать вывод, что разделение на сеоки действительно обусловлено генетическим родством по мужской линии.
Работа выполнена в рамках Научно-технической программы Союзного государства “Разработка инновационных геногеографических и геномных технологий идентификации личности и индивидуальных особенностей человека на основе изучения генофондов регионов Союзного государства” (“ДНК-идентификация”), государственный контракт № 011-17 от 26.09.2017, а также поддержана грантом РФФИ № 18-29-13045 “Популяционная геномика и транскриптомика человека: поиск сигналов не-нейтральной эволюции”.
Все процедуры, выполненные в исследовании с участием людей, соответствуют этическим стандартам институционального и/или национального комитета по исследовательской этике и Хельсинкской декларации 1964 г. и ее последующим изменениям или сопоставимым нормам этики.
От каждого из включенных в исследование участников было получено информированное добровольное согласие.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Тюркские народы Сибири / Под ред. Функ Д.А., Томилов Н.А. Ин-т этнологии и антропологии им. Н.Н. Миклухо-Маклая РАН; Омский филиал Института археологии и этнографии СО РАН. М.: Наука, 2006. 678 с.
http://www.perepis-2010.ru/.
Генофонд населения Сибири: Сборник научных статей / Под. ред. Коненкова В.И., Пузырева В.П., Воеводы М.И. Новосибирск: Ин-т археологии и этнографии СО РАН, 2003. С. 174–176.
Харьков В.Н., Хамина К.В., Медведева О.Ф. и др. Разнообразие генофонда хакасов: внутриэтническая дифференциация и структура гаплогрупп Y-хромосомы // Мол. биология. 2011. Т. 45. № 3. С. 446–458.
Харьков В.Н., Новикова Л.М., Лузина Ф.А и др. Анализ генофонда и родоплеменной структуры шорцев по маркерам Y-хромосомы // Мед. генетика. 2016. Т. 15. № 5. С. 48–51. doi.org/10.1234/XXXX-XXXX-2016-5-48-51
Бутанаев В.Я. Происхождение хакасов по данным этнонимики // Проблемы археологии и этнографии. Л., 1983. Вып. 2. С. 68–73.
The Y-Chromosome Consortium. A nomenclature system for the tree of human Y-chromosmal binary haplogroups // Genome Res. 2002. V. 12. P. 339–348. https://doi.org/10.1101/gr.217602
http://www.isogg.org/.
Ilumäe A.-M. Reidla M., Chukhryaeva M. et al. Human Y-chromosome haplogroup N: a non-trivial time-resolved phylogeography that cuts across language families // Am. J. Hum. Genet. 2016. V. 99. P. 163–173. https://doi.org/10.1016/j.ajhg.2016.05.025
Bandelt H.-J., Forster P., Rohl A. Median-joining networks for inferring intraspecific phylogenies // Mol. Biol. Evol. 1999. V. 16. P. 37–48. https://doi.org/10.1093/oxfordjournals.molbev.a026036
Sengupta S., Zhivotovsky L.A., King R. et al. Polarity and temporality of high resolution Y-chromosome distributions in India identify both indigenous and exogenous expansions and reveal minor genetic influence of central Asian pastoralists // Am. J. Hum. Genet. 2006. V. 78. P. 202–221. https://doi.org/10.1086/499411
Бутанаев В.Я. Происхождение хакасских родов и фамилий. Абакан, 1994. 93 с.
Потапов Л.П. Происхождение и формирование хакасской народности. Абакан: Хакасское кн. изд-во, 1957. 307 с.
Историческая этнография: традиции и современность / Проблемы археологии и этнографии. Вып. II. Л.: Изд-во ЛГУ, 1983. 184 с.
Харьков В.Н., Хамина К.В., Медведева О.Ф. и др. Структура генофонда тувинцев по маркерам Y‑хромосомы // Генетика. 2013. Т. 49. № 12. С. 1416–1425. https://doi.org/10.7868/S0016675813120035
Кимеев В.М. Проблема этногенеза шорцев // Вестник Кемеровского гос. ун-та. 2014. № 3-3(59). С. 54–66.
Алексеев В.П. Антропологические данные и проблемы происхождения шорцев // Уч. записки Хак-НИИЯЛИ. Абакан, 1965. Вып. XI. С. 86–100.
Долгих Б. О. Родовой и племенной состав народов Сибири в XVII в. // ТИЭ АН СССР. Нов. сер. М.; Л., 1960. Т. 55. С. 104–118.
Бутанаев В.Я. Вопрос о самоназвании хакасов // Этнограф. обозрение. 1992. № 2. С. 63–69.
Вайнштейн С.И. Очерк этногенеза тувинцев // Уч. записки ТувНИИЯЛИ Кызыл, 1957. Вып. 5. С. 198.
Алексеев В.П., Гохман И.И. Антропология азиатской части СССР. М., 1984. 208 с.
Окладников А.П. Древнее население Сибири и его культура: Сборник “Народы Сибири”. М.; Л.: Изд-во АН СССР, 1956. С. 21–107.
Антропо-экологические исследования в Туве / Под ред. Алексеевой Т.И., Урысон М.И. М.: Наука, 1984. 225 с.
Дополнительные материалы отсутствуют.