

Генетика, 2021, T. 57, № 1, стр. 82-94
Множественный хромосомный полиморфизм, хромосомной расы “эворон” эворонской полевки (Rodentia, Arvicolinae)
И. В. Картавцева 1, *, И. Н. Шереметьева 1, М. В. Павленко 1
1 Федеральный научный центр биоразнообразия наземной биоты Восточной Азии Дальневосточного отделения Российской академии наук
690022 Владивосток, Россия
* E-mail: Kartavtseva@biosoil.ru
Поступила в редакцию 26.02.2020
После доработки 11.05.2020
Принята к публикации 08.07.2020
Аннотация
На основании анализа собственных и литературных данных полевок берегов о. Эворон (n = 8) и р. Амгунь (n = 8), в Эворон-Чукчагирской низменности, а также особей лабораторного разведения, приведены новые данные о кариотипической изменчивости (2n = 38–41, NF = 54–59), хромосомных преобразованиях и их сочетаниях в кариотипе. Нумерация хромосом в кариотипе эворонской полевки с использованием методов дифференциального (GTG–) окрашивания позволила обозначить пары хромосом, вовлеченные в различные хромосомные перестройки, а для пар, образованных в результате слияния хромосом, дать номера парам их образовавшим. Тандемные слияния теломера-теломера (TTel) двуплечих хромосом образуют крупный субметацентрик Mev1/4М; слияние теломера-центромера (TCen) в результате инактивации центромеры одной из пар акроцентриков образуют либо акроцентрик (Mev17/18A), либо метацентрик (Mev17/18M), а робертсоновское слияние этих же пар – образует метацентрик средних размеров (Mev17/18M). Также показана изменчивость положения центромеры в двух парах аутосом (Mev8 и Mev14). Выявленные перестройки позволили описать двенадцать вариантов кариотипа: два с 2n = 41, шесть с 2n = 40, три с 2n = 39 и один с 2n = 38. Предложено отнести полевок Эворон-Чукчагирской низменности, имеющих слияние двуплечих хромосом Mev1 и Mev4, к хромосомной расе “эворон”, остальных – к расе “арги”. Получение плодовитого потомства от особей, имеющих в кариотипе слияние TTel и высокий процент этой перестройки у особей из природных выборок, свидетельствует об отсутствии ее вредного влияния на жизнеспособность полевок.
Восточноазиатские серые полевки рода Alexandromys (=Microtus) (Arvicolini Gray, 1821) характеризуются слабой морфологической дифференциацией [1–4] и высокой скоростью процессов видообразования, сопровождающихся хромосомными перестройками [2, 5, 6]. Кариологические характеристики (2n и NF) видов этого рода являются хорошими дифференцирующими признаками [2, 7–10]. Род включает 12 видов, хромосомные числа которых варьируют от 30 до 52. Для трех видов отмечен стабильный кариотип: A. kikuchii Kuroda, 1920 (2n = 30, NFa = 48) [11], A. sachalinensis Vasin, 1955 (2n = 50, NFa = 60) [12] и A. gromovi Vorontsov et al., 1988 (2n = 44, NFа = 58) [13]; причем первые два вида являются эндемиками островов. Для трех видов отмечены редкие случаи изменчивости числа хромосом посредством перестроек робертсоновского типа: A. oeconomus Vasin, 1955 (2n = 30–32, NFа = 58) [14, 15] A. montebelli Milne–Edwards, 1872 (2n = 30–31, NFa = 54) [16], и A. mongolicus Radde, 1861 (2n = 49–50, NFа = 56) [2]. Для четырех видов описана изменчивость положения центромеры (или инверсии) при стабильном числе хромосом: A. fortis Büchner, 1889 (2n = 52, NFa = 62–64) [17], A. middendorffii Poljakov, 1881 (2n = 50, NFa = 54–56) [2], A. limnophilus Büchner, 1889 (2n = 38, NFа = 56–58) [18], A. mujanensis Orlov et Kovalskaya, 1978 (2n = = 38, NFa = 46–49) [19]. И только для двух видов обнаружена изменчивость числа и морфологии хромосом, сопряженная с множественными хромосомными перестройками: A. evoronensis Kovalskaya et Sokolov, 1980 (2n = 38–40, NF = 52–58) [2, 9] и A. maximowiczii Schrenk, 1859 (2n = 36–44, NFa = = 50–60) [2, 10]. По молекулярно-генетическим данным показано, что полевки группы “maximowiczii” A. mujanensis и A. evoronensis близки к A. maximowiczii, и в одной из последних работ было предложено рассматривать первые два вида в составе A. maximowiczii [20]. Несмотря на то, что мы соавторы этой статьи, считаем, что ранг этих таксонов все еще остается спорным, и они нуждаются в дополнительных исследованиях, особенно гибридологических, поэтому в настоящей работе рассматриваем A. evoronensis как самостоятельный вид.
Внутрипопуляционный множественный хромосомный полиморфизм последних двух видов исследован слабо [2, 7, 9, 10, 21]. Для A. evoronensis при описании вида, основанном на стерильности F1 при ссаживании их с A. maximowiczii и кариологических особенностях [9], были выявлены вариации диплоидного числа и плеч хромосом эворонской полевки (2n = 38–40, NF = 52–58). В этой работе авторы выявили шесть структурных перестроек: два центрических слияния, однотеломерно-центромерное слияние и три перицентрических инверсии, половые хромосомы описаны как акроцентрические. Вслед за этой работой другими авторами [22] при использовании дифференциального (G–) окрашивания хромосом пяти особей из природной популяции, расположенной недалеко от terra typica, были отмечены иные хромосомные перестройки при той же изменчивости числа хромосом (38–40), половые хромосомы описаны как акроцентрические. Различие числа хромосом объясняли тандемным слиянием двух двуплечих пар хромосом. Позже, на основе изучения кариотипов ранее исследованных полевок [22] и их лабораторного потомства (2n = 38–40, NF = 53–56) [2], были выявлены четыре типа перестроек: тандемное слияние двуплечих пар, одно робертсоновское слияние и изменение положения центромеры в двух парах аутосом. Изменение положения центромеры связывали как с различными типами слияния хромосом, так и перицентрическими инверсиями. Морфология половых хромосом была уточнена: Х-двуплечий, а Y-одноплечий элементы. В работе приведены фото шести раскладок хромосом с различными диплоидными числами и сочетаниями хромосомных перестроек (две раскладки с 2n = 40, две с 2n = 39 и одна с 2n = 38). Варианты кариотипа не были описаны, номера двуплечих пар хромосом (D), вовлеченные в тандемное слияние определены как D1 и D6, акроцентрические пары хромосом, участвующие в слиянии хромосом с образованием двуплечей хромосомы (D4) не определены.
Ранее считали, что данный вид обитает только в Эворон-Чукчагирской низменности, расположенной в Хабаровском крае России [23] соответственно и кариотипы исследованы из долины оз. Эворон этой низменности [2, 9, 24, 25]. Недавно нами были обнаружены две изолированные популяции эворонской полевки – одна в Верхнебуреинской впадине Хабаровского края [25], другая в северо-восточной части Амурской обл. [26], видовая диагностика которых была проведена по данным мтДНК. Полевки этих популяций в кариотипах имели 37 и 36 хромосом соответственно, что не входит в размах изменчивости числа хромосом, описанного для вида ранее (38–40) [2, 9]. На основании этих данных, нами было высказано предположение, что эворонская полевка может иметь две хромосомные расы [27].
Чтобы охарактеризовать кариотипическую изменчивость в географически изолированных популяциях вида с числами хромосом 36 и 37 (что и планируется сделать в следующей работе), необходимо сначала описать кариологические особенности вариантов кариотипа полевок из Эворон-Чукчагирской низменности, выявив пары хромосом, вовлеченные в хромосомные перестройки. Поэтому целью настоящего исследования было исследовать кариотипы полевок в Эворон-Чукчагирской низменности, дать номера парам хромосом кариотипа с максимальным числом одноплечих хромосом, на основе собственных и литературных данных описать варианты кариотипа и хромосомные перестройки, характеризующие эти варианты.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом хромосомного исследования послужили 27 полевок: десять особей эворонской полевки, отловленных авторами в трех природных локальных популяциях Эворон-Чукчагирской равнины (рис. 1), и семнадцать – лабораторного разведения. Исследованным полевкам присваивали тройной номер: номер хромосомного препарата/номер образца тканей/номер сиквенса контрольного региона в Генбанке NCBI. Последний номер приведен с целью привязки кариологических данных настоящей работы кранее опубликованным молекулярно-генетическим данным (если таковые имеются) [25, 26, 28].
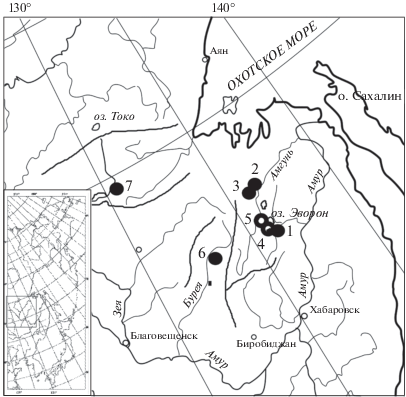
Точка № 1 – самец 2050/81–06/HM135862 – отловлен в сентябре 2006 г. в долине проточного оз. Эворон, на осоково-вейниковом закочкаренном лугу близ развилки дорог на пос. Харпичан и пос. Эворон (51°19′ с.ш., 136°35′ в.д.). Точка № 2 – три самца: 2355/50–08/HM135861, 2369/87–08 и 2370/88–08/НМ135859 – отловлены на разнотравном лугу в июле 2008 г., 4 км на север от пос. им. Полины Осипенко, близ слияния рек Нимелен и Амгунь (52°26′ с.ш., 136°34′ в.д.). Точка № 3 – шесть особей: самец 0060/58–16, самка 0061/94–16, самка 0062/93–16, самка 0063/92–16, самец 0064/91–16 и самка 0066/95–16 – отловлены в июле 2016 г., дамба заросшая травой и ивой, 1 км восточнее пос. им. Полины Осипенко, долина р. Амгунь (52°25.42′ с.ш., 136°29.67′ в.д.). Семнадцать полевок лабораторного разведения получены от четырех природных животных, отловленных в точке № 3: самцов 0060/58–16 и 0064/91–16, самок 0062/93–16 и 0061/94–16. Этим полевкам присваивали двойные номера. В разведении эворонских полевок использованы апробированные методы [2, 29].
Хромосомные препараты, ткани для молекулярного анализа и черепа хранятся в коллекции ФНЦ Биоразнообразия ДВО РАН.
Также с целью выявления вариантов кариотипов эворонской полевки использованы ранее опубликованные данные хромосом из двух точек (рис. 1): точка № 4 (terra typica) – берег р. Девятка, единственной реки, вытекающей из оз. Эворон [9]; точка № 5 – долина р. Одон, впадающей в оз. Эворон [2, 22]. Ранее в двух работах [2, 9] максимальное число плеч хромосом было определено как 58. Однако по данным опубликованной авторами раскладки хромосом при первоописании вида (полевка из точки 4) [9] оно равно 59, поэтому мы считаем это число верным и учитываем в нашей работе.
Суспензии хромосом для полевок точек № 1 и 3 приготовлены из клеток костного мозга бедренной кости по стандартной методике [30], для полевок точки № 2 использован метод кратковременной культуры клеток костного мозга [31]. Гомологию пар хромосом выявляли при помощи GTG-метода [32], окрашивание на структурный гетерохроматин проводили с использованием метода, предложенного Самнером [33], без предварительной обработки препаратов соляной кислотой. Окрашивание ядрышкового организатора (ЯОР) проведено без предварительной обработки муравьиной кислотой [34].
Исходным кариотипом для присвоения номеров пар хромосом послужил кариотип с максимальным числом 2n (42), включающий 26 акроцентрических (A) и 14 двуплечих (М) аутосом (табл. 1). Парам хромосом, образованным в результате слияния хромосом, присваивали дробный номер, состоящий из номеров пар слившихся хромосом и указанием ее морфологии (А или М). Для выявления вариантов кариотипа эворонской полевки, опубликованного ранее из долины оз. Эворон [2, 9, 35 ], мы применили новую нумерацию пар хромосом. Ранее парам хромосом кариотипа с 2n = 40 (16 двуплечих и 22 одноплечих аутосом) были даны номера раздельно двуплечим (D) и одноплечим (A) хромосомам [2, 9], затем для ранее опубликованного кариотипа [2, 9] была дана сквозная нумерация пар [35]. Поскольку для двуплечей хромосомы, образованной в результате слияния акроцентрических пар хромосом, в последней работе был дан порядковый номер, не указывающий на слияние, а пары, участвовавшие в перестройке, не имели номера, мы приводим иную нумерацию хромосом. А именно: кариотипу с максимальным диплоидным числом хромосом (41), имеющему 26 одноплечих аутосом, даем сквозную нумерацию. Для определения числа хромосом анализировали не менее 20 хромосомных пластинок и не менее трех раскладок для каждой особи. Метафазные пластинки анализировали с помощью микроскопа Axio Imager 1 с цифровой камерой AxioCamHR и программного обеспечения AXIOVISION 4.7 (Германия) центра коллективного пользования ФНЦ Биоразнообразия.
Таблица 1.
Хромосомные характеристики вариантов кариотипа и схема изменчивых пар аутосом эворонской полевки Alexandromys evoronensis Эворон-Чукчагирской низменности
| 2n | NF | Число аутосом |
Номера пар аутосом и их морфология | Число перестроек | Источник | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M | A | 1, 4 | 1/4 | 17, 18 | 17/18 | 8 | 13 | всего | ГС | |||
| 42* | 58 | 14 | 26 | xxxx | v vvv | v v | v v | 0 | 0 | Нет данных | ||
| 41a | 56 | 13 | 26 | x x | X | v vvv | v v | v v | 1 | 1 | Наши данные | |
| b | 57 | 14 | 25 | x x | X | v vvv | vx | v v | 2 | 2 | » | |
| 40a | 57 | 15 | 23 | xxxx | vx | v v | v v | 1 | 1 | [2] + наши данные | ||
| b | 56 | 14 | 24 | xxxx | xx | v v | v v | 1 | 0 | [2] | ||
| c | 59 | 17 | 21 | xxxx | vx | xx | v v | 2 | 1 | [9] | ||
| d | 56 | 14 | 24 | x x | X | v v | x | xx | v v | 3 | 1 | [2] |
| e | 54 | 12 | 26 | XX | v vvv | v v | v v | 1 | 0 | Наши данные | ||
| f | 55 | 13 | 25 | XX | v vvv | vx | v v | 2 | 1 | » | ||
| 39a | 55 | 14 | 23 | XX | v v | x | vx | vx | 3 | 2 | [2] | |
| b | 56 | 15 | 22 | x x | X | xx | v v | v v | 2 | 1 | [9] | |
| c | 55 | 14 | 23 | x x | X | vx | xx | v v | 3 | 2 | [2] | |
| 38a | 55 | 15 | 21 | XX | xx | v v | vx | 3 | 1 | [2] | ||
РЕЗУЛЬТАТЫ
Исследование хромосомных наборов A. evoronensis двадцати семи особей (10 природных и 17 лабораторного разведения) полевок Эворон-Чукчагирской низменности выявило новое максимальное диплоидное число (41), ранее оно было равно 40, и число акроцентрических пар аутосом (26). Хромосомные числа исследованных полевок точек № 1, 2 и 3 варьировали от 40 до 41 (рис. 2).
Рис. 2.
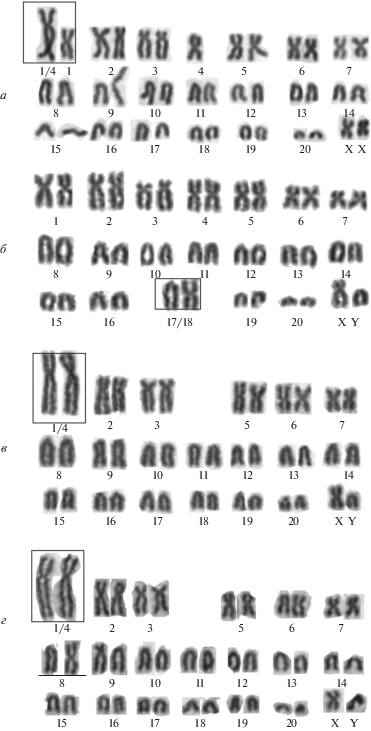
Варианты кариотипа эворонской полевки Alexandromys evoronenesis. а – 2n = 41a, NF = 56 (точка № 2, самка 0063/ 92–16) 13 двуплечих (M), 26 одноплечих (A) аутосом и один крупный субметацентрик (Mev1/4), образованный в результате тандемного слияния пар Mev1 и Mev4; б – 2n = 40a, NF = 57 (точка 1, самец 2050/81–06/HM135862), 15 M и 23 A. Одна пара хромосом гетероморфная – Mev17/18A и Mev17/18M; в – 2n = 40e, NF = 54 (точка № 3, самец 4183, лабораторное разведение), 12 М и 26 А аутосом, тандемное слияние пар Mev1 и Mev4 в гомозиготном состоянии (Mev1/4M и Mev1/4M); г – 2n = 40f, NF = 55 (точка № 3, самец 4527, лабораторное разведение), 13 М, 25 А, тандемное слияние пар Mev1 и Mev4 в гомозиготном состоянии (Mev1/4, Mev1/4) и гетероморфная пара Mev8М и Mev8А.
В результате анализа дифференциально-окрашенных хромосом (рис. 3) и изменчивости числа и морфологии хромосом собственных и литературных данных для точек № 4 [9] и 5 [2] было показано, что диплоидные числа вида варьируют от 38 до 41. С учетом различных хромосомных перестроек выявлено 12 вариантов кариотипа (табл. 1). Дифференциальное окрашивание хромосом эворонской полевки позволило дать номера парам хромосом кариотипа и парам, вступающим в хромосомные перестройки, при формировании двенадцати вариантов кариотипа (рис. 3).
Рис. 3.
Дифферциально окрашенные хромосомы эворонской полевки Alexandromys evoronenesis из двух точек. а – точка № 3, кариотип 2n = 40e, NF = 54 (самец № 4183, лабораторное разведение), 12 М и 26 А аутосом: GTG-окрашивание; б – C-окрашивание, стрелками указаны теломерные С-блоки; в – NOR-окрашивание, ядрышкообразующие районы в прицентромерных районах двух пар аутосом (17 и 20); г, д – локалитет № 1, кариотип 2n = 40a, NF = 57 (самец 2050/81-06/HM135862), 15 M и 23 A. Пара 17/18 гетероморфная (подчеркнута): акроцентрическая хромосома образована путем тандемного слияния (центромера–теломера), метацентрическая – путем центромерного слияния (центромера–центромера). GTG-окрашивание (г), C-окрашивание (д).
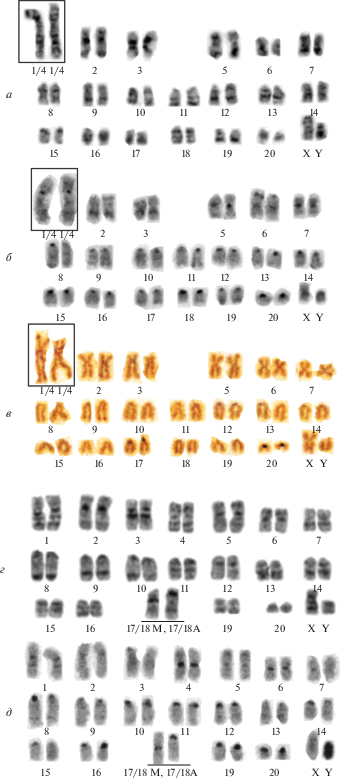
Вариант 2n = 41a, NF = 56 (рис. 2,а) имеет 13 двуплечих (M) и 26 одноплечих (A) аутосом. Среди двуплечих аутосом присутствует один крупный субметацентрик. Дифференциальное окрашивание позволило определить метацентрические пары хромосом Mev1 и Mev4, которые в результате тандемного слияния теломера–теломера (TTel) образовали крупный субметацентрик (Mev1/4M) (рис. 4,а), где активирован центромерный район хромосомы Mev4. В этом варианте кариотипа все три хромосомы (Mev1, Mev4 и Mev1/4M) находятся в гетерозиготном состоянии. Такой вариант имели самцы точек № 2 (2369/87–08; 2370/88-08; 2355/50–08) и особи точки № 3 (самец 0060/58–16; самки 0063/92–16 и 0066/95–16); встречен у особей точки № 2 с частотой 1.0 и точки № 3 – с частотой 0.67. Для популяции берега р. Амгунь (точки № 2 и 3) частота равна 0.75. Окрашивание на структурный гетерохроматин выявило прицентромерный гетерохроматин во всех парах хромосом. В Х-хромосоме присутствует слабо окрашенный С-блок в прицентромерном районе длинного плеча. Y-хромосома полностью гетерохроматиновая. Ядрышкообразующие районы локализованы в прицентромерных районах двух пар аутосом – Mev17A и Mev20A (рис. 2,в).
Рис. 4.
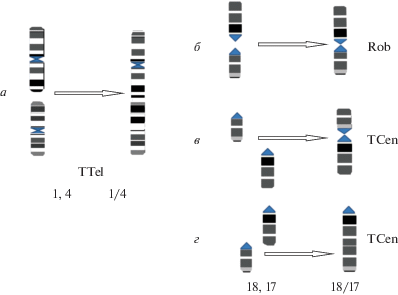
Схемы хромосомных перестроек эворонской полевки Alexandromys evoronenesis. а – тандемное слияние теломера–теломера (TTel) образовали крупный субметацентрик (Mev1/4M). Слияния акроцентрических хромосом Mev17A и Mev18A; б – робертсоновское слияние с образованием метацентрика Mev17/18M; В – тандемное слияние – центромера–теломера (TCen) с образованием метацентрика Mev17/18M; г – тандемное слияние центромера–теломера (TCen) с образованием акроцентрика Mev17/18A.
Вариант 2n = 41b, NF = 56 (табл. 1): 14 двуплечих и 25 одноплечих аутосом. От кариотипа 2n = 41a отличается одной парой аутосом (Mev8), находящейся в гетерозиготном состоянии по морфологии гомологов, представленных акроцентриком (А) и метацентриком (М). Этот вариант обнаружен у особей лабораторного разведения. Метацентрический вариант хромосомы появился в результате ссаживания полевок вариантов кариотипов 40f и 41a.
Варианты с 2n = 40. По данным собственных и литературных данных обнаружено шесть вариантов кариотипа, имеющего 40 хромосом (табл. 1), различающихся сочетаниями хромосомных перестроек таких как тандемное слияние (TTel) метацентрических пар Mev1M и Mev4M с образованием крупного субметацентрика (Mev1/4M); робертсоновское слияние акроцентрических пар Mev17A и Mev18A c образованием метацентрика Mev17/18M (рис. 4,б) и тандемное слияние – центромера-теломера (TCen) акроцентрических пар Mev17A и Mev18A c образованием акроцентрика Mev17/18A, где активирована центромера хромосомы Mev18A (рис. 4,г). В нашем исследовании в кариотипе полевок с 2n = 40 (точки № 2 и 3) были обнаружены два варианта слияния акроцентрических хромосом (рис. 4). Третий вариант TCen с образованием метацентрика Mev17/18M, описанный в работе М.Н. Мейер с соавт. [2], в точке № 5 нами не обнаружен.
Вариант 2n = 40a, NF = 57 (рис. 2,б, табл. 1): 15 двуплечихи 23 одноплечих аутосом. Одна пара хромосом гетероморфная – Mev17/18A и Mev17/18M. При дифференциальном окрашивании хромосом (GTG) виден различный рисунок окрашивания гетероморфных хромосом (рис. 3,г). На рис. 4 приведена схема слияния акроцентрических хромосом (рис. 4,б, г). Окрашивание на структурный гетерохроматин показало наличие прицентромерного гетерохроматина во всех парах аутосом, включая гетероморфную пару. Акроцентрические хромосомы Mev17А и Mev18A, участвовавшие в слиянии, имели гетерохроматиновые блоки в центромерных районах. У метацентрической хромосомы Mev17/18М гетерохроматин выявлен в центромерном районе хромосомы, тогда как при тандемном слиянии гетерохроматиновый блок отмечен только в центромерном районе акроцентрической хромосомы Mev17/18A (рис. 3,д). Такой вариант кариотипа обнаружен у особей точек № 1 (2050/81–06/HM135862) и № 3 лабораторного разведения (4437/297–18). К такому варианту можно отнести ранее обнаруженный кариотип полевки из точки № 5 [2].
Вариант 2n = 40b, NF = 56 (табл. 1): 14 М и 24 А аутосом. Отличается от варианта 2n = 40a тем, что пара Mev17/18M находится в гомозиготном состоянии. Такой вариант имеет одна из полевок точки № 5 [2].
Вариант 2n = 40с, NF = 59 (табл. 1): 17 М и 21 А аутосом. Отличается от 2n = 40a и 2n = 40b тем, что имеет гетероморфную пару: Mev17/18A и Mev17/18M. Пара Mev8M представлена двумя метацентрическими хромосомами. Этот вариант имеет максимальное число плеч хромосом (59) и обнаружен в точке № 4 [9].
Вариант 2n = 40d, NF = 56 (табл. 1): 14 М и 24 А аутосом. Кариотип имеет гетерозиготное состояние хромосом (Mev1M, Mev4M, Mev1/4M, Mev17A, Mev18A, Mev17/18М), где присутствуют два типа слияния одновременно: TTel (Mev1/4M) и робертсоновское (Mev17/18М). Такой вариант имела она из полевок точки № 5 [2].
Вариант 2n = 40e, NF = 54 (рис. 2,в, 3,а, б, в): 12 М и 26 А аутосом. Кариотип имеет TTel-слияние (Mev1/4M) в гомозиготном состоянии. Такой вариант обнаружен нами в точке № 3 (самка 0062/93–16).
Вариант 2n = 40f, NF = 55 (рис. 2,г): 13 М, 25 А. Кариотип имеет TTel-слияние (Mev1/4M) в гомозиготном состоянии и гетероморфную пару – Mev8М, А. У одного из гомологов пары Mev9 имеется хорошо выраженное короткое плечо, и такую хромосому можно назвать субтелоцентрической. Однако в настоящей работе мы относим эту хромосому к акроцентрической и не учитываем при подсчете числа плеч аутосом. Такой вариант обнаружен нами в точке № 3 (самец 0064/91–16).
Вариант 2n = 39a, NF = 55 (табл. 1): 14 М и 23 А аутосом. Кариотип имеет гетерозиготное состояние трех хромосом: Mev17/18M, Mev17А и Mev18А. Две пары аутосом по характеру их морфологии находятся в гетероморфном состоянии (Mev8М, Mev8А и Mev13М, Mev13А). Такой вариант обнаружен нами в точке № 5 [2].
Вариант 2n = 39b, NF = 56 (табл. 1): 15 М и 22 А аутосом. Кариотип имеет TTel-слияние (Mev1/4M) в гетерозиготном состоянии (Mev1М, Mev4M, Mev1/4M) и слияние акроцентрических хромосом Mev17/18М в гомозиготном состоянии. Такой вариант обнаружен в точке № 4 [9].
Вариант 2n = 39с, NF = 55 (табл. 1): 14 М и 23 А аутосом. Кариотип имеет TTel (Mev1/4M), робертсоновское (Mev17/18М) и ТCen (Mev17/18A) слияния в гетерозиготном состоянии (Mev1М, Mev4М, Mev1/4M, Mev17/18М, Mev17/18A). Такой вариант обнаружен в точке № 5 [2].
Вариант 2n = 38a, NF = 55 (табл. 1): 15 М и 21 А аутосом. Кариотип характеризуется слияниями TTel (Mev1/4) и робертсоновским (Mev17/18М), образованные пары находятся вгомозиготном состоянии. Пара Mev13 гетероморфная – Mev13M, A. Такой вариант обнаружен нами в точке № 5 [2].
Окрашивание на структурный гетерохроматин (г/х). Небольшое количество г/х присутствует в прицентромерных районах всех аутосом. Степень его окрашивания от слабого до яркого в различных метафазах варьировало. Также для полевок из точек № 2 и 3 (долина р. Амгунь) отмечены С‑блоки в теломерном районе короткого плеча пары хромосом Mev2. Однако в кариотипе 40а полевки из точки № 1 (рис. 3,д) (долина оз. Эворон) пара аутосом Mev2 таких блоков не имела.
Кариотипы полевок долины р. Амгунь. В точке № 2 у двух особей был кариотип 41а. В точке № 3 диплоидные числа варьировали от 40 до 41. Так, один самец (0060/58–16) и две самки (0063/92–16 и 0066/95–16) имели кариотип 41а, один самец (0064/91–16) – кариотип 40f и одна самка – 40e (0062/93–16). Еще у одной самки, давшей потомство (0061/94–16), кариотип не определен, но в кариотипе потомства (2n = 40a) присутствовала робертсоновская перестройка (Mev17/18M) и тандемное слияние TCen(Mev17/18A). При разведении полевок, нам не удалось сохранить особей с таким вариантом кариотипа. При ссаживании полевок с вариантами кариотипа 2n = 40f и 2n = 40e, получена линия со стабильным числом хромосом (40), но при этом с изменчивой морфологией пары Mev8 (М, А). При ссаживании самки с кариотипом 2n = 40е с самцами вариантов карио-типа 2n = 40f и 2n = 41а получена линия с числами хромосом 40 и 41. Наиболее жизнеспособными оказались особи вариантов кариотипа с тандемным слиянием двуплечих пар аутосом (TTel) и образованием крупного субметацентрика Mev1/4M как в гомозиготном (2n = 40e, 2n = 40f), так и гетерозиготном состоянии (2n = 41а и 2n = 41b). На основании того, что почти все отловленные нами полевки в точках № 2 и 3, имели слияние TTel, а при лабораторном разведении получено потомство с такой перестройкой, можно заключить, что слияние TTel не является вредным в исследуемой популяции. Что касается второго тандемного слияния (TCen) акроцентрических пар с образованием акроцентрика Mev17/18А, то жизнеспособность полевок с такой перестройкой возможно снижена. Нами в виварных условиях (как было сказано выше) были получены полевки с такой перестройкой, но получить от них потомство не удалось.
Кариотипы полевок долины оз. Эворон. Числа хромосом двадцати семи полевок из природы и семи особей лабораторного разведения из точек № 1, 4 и 5 варьируют от 38 до 40 (табл. 2). Имеющиеся в нашем распоряжении собственные и литературные [2, 9] данные только для восьми особей, включая одну полевку лабораторного разведения, позволили описать восемь вариантов кариотипа: один для 2n = 38, три – 2n = 39 и четыре – 2n = 40. Кариотипы с 2n = 41 не обнаружены. Изменчивость кариотипа сопряжена с такими хромосомными перестройками, как TTel, TCen, центрическим слиянием и смещением центромеры.
Таблица 2.
Варианты кариотипа эворонской полевки Alexandromys evoronensis двух долин в Эворон-Чукчагирской низменности
| Долины | Локалитет (число особей) | 2n | |||
|---|---|---|---|---|---|
| 38 | 39 | 40 | 41 | ||
| Оз. Эворон | 1 (1) | – | – | a | – |
| 4 (20*) | + | b | c | – | |
| 5 (5**) | + | a, c | a, b, c | – | |
| Лабораторные 5 (7**) | ? | ? | d | – | |
| Р. Амгунь | 2 (2) | – | – | – | a |
| 3 (6) | – | – | e, f | a, | |
| Лабораторные 3 (17) | – | – | a, e, f | a, b | |
Примечание. + – вариант обнаружен, ? – нет данных, жирным выделены варианты, появившиеся в результате виварного разведения; “–” – вариант не обнаружен; * – исследованы пределы изменчивости 2n и NF, варианты кариотипа определены по двум опубликованным раскладкам [9]; ** – варианты кариотипа определены по шести опубликованным раскладкам [2].
Слияние теломера–теломера. Тандемное слияние (TTel) двуплечих хромосом с образованием крупного субметацентрика Mev1/4 присутствует в девяти вариантах кариотипа из двенадцати обнаруженных, причем в пяти из них – в гетерозиготном состоянии. Из девяти кариотипированных полевок природной популяции берега р. Амгунь все особи имели слияние TTel (Mev1/4), причем, шесть особей имели гетерозиготное состояние по этой перестройке с частотой 0.67. При лабораторном разведении полевок преобладали особи с перестройкой TTel в гомозиготном состоянии. В природных популяциях точек № 4 и 5, были обнаружены особи с такой перестройкой в кариотипах вариантов 2n = 40d, 2n = 39a, b, c, 2n = 38 (табл. 2). Если в точке № 4 частота данных хромосомных перестроек неизвестна, то в точке № 5 (из пяти полевок) TTel выявили в кариотипе двух особей в гомозиготном состоянии и у одной – в гетерозиготном. В точке № 1 одна особь не имела такой перестройки. Таким образом, хромосомные числа полевок Эворон-Чукчагирской низменности варьировали от 38 до 41. Частота теломерного слияния (TTel) в кариотипах здесь равна 0.87, в гетерозиготном состоянии– 0.47. Единственный самец, отловленный нами в окрестностях оз. Эворон, не имел такой перестройки.
ОБСУЖДЕНИЕ
Обнаруженные в настоящей работе числа хромосом эворонской полевки Эворон-Чукчагирской равнины варьируют от 38 до 41, числа плеч – от 54 до 59 (табл. 1). Полученные данные позволяют предполагать, что максимальное число хромосом равно 42. Ранее для данного вида были приведены иные характеристики кариотипа: 2n = 38–40, NF = = 54–58 и без описания вариантов кариотипа и хромосомных перестроек, характеризующих эти варианты [2]. Впервые в настоящей работе выявлен кариотип, имеющий максимальное из обнаруженных для вида число хромосом (41) и одноплечих аутосом (26). Изменчивость числа хромосом обусловлена слияниями четырех пар аутосом – Mev1M, Mev4M, Mev17A и Mev18A. Тандемное слияние пар Mev1M и Mev4M (TTel), с образованием Mev1/4M, приводит к уменьшению числа хромосом и плеч аутосом. Слияние (TCen) акроцентрических пар Mev17A и Mev18A приводит к образованию Mev17/18M, Mev17/18A. Эти два варианта слияний обнаружены у полевки из популяции № 1 (рис. 2,б, рис. 3,г, д). Третий вариант тандемного слияния, с образованием двуплечего элемента, обнаружен в популяции № 5 [2], где активирована центромера хромосомы Mev17. Нами такой вариант не обнаружен, но поскольку имеется описание этой перестройки, мы приводим ее схему (рис. 4,г).
Изменчивость морфологии аутосом (и как следствие изменение числа плеч) происходит также и за счет изменения положения центромеры или инверсии в данных парах: Mev8A и Mev14A. Впервые изменчивость морфологии данных пар аутосом была обнаружена для полевок берегов оз. Эворон [2]. Авторами было предложено два сценария перестройки этих пар аутосом – перицентрическая инверсия и тандемное слияние акроцентрических хромосом, аналогичные таковым в паре Mev17/18. Возможно, то, что мы называем смещением центромеры, является инактивацией одной из центромер слившихся акроцентрических пар аутосом. Таким образом, число вариантов кариотипа эворонской полевки, с учетом всех перестроек хромосом, должно быть значительно больше, чем описано нами.
Исследование кариотипов эворонской полевки из локальных популяций долин оз. Эворон и р. Амгунь позволяет говорить, что они имеют сходный характер перестроек хромосом, но различные числа хромосом (табл. 2). Эти популяции имеют два варианта тандемных слияний и одно робертсоновское.
Несмотря на то, что мы не выявили низкие числа хромосом (38 и 39) в долине р. Амгунь (точках № 2, 3), наличие особей с двумя тандемными и робертсоновским слияниями в этих популяциях позволяет предположить, что при увеличении числа исследованных полевок, мы можем ожидать появления здесь кариотипов с этими числами. Однако по распределению гетерохроматина во второй паре аутосом (Mev2) мы наблюдаем различия между полевками берегов оз. Эворон (точки № 1, 5) и р. Амгунь (точки № 2, 3). Возможно отличия касаются и аутосомы Mev9, в которой появляется короткое плечо. Из-за исследования небольшого объема выборок мы не можем говорить, что обнаруженные различия дифференцируют популяции. О слабой дифференциации полевок берегов р. Амгунь и оз. Эворон свидетельствуют и полученные нами ранее данные контрольного региона мтДНК [26]. Так, было выявлено только десять фиксированных замен между гаплотипами особей (HM135858–HM135861), отловленных в точке № 2, и гаплотипом особи (номер HM135857) точки № 4.
Нами впервые сделано окрашивания ЯОР для эворонской полевки. Для видов рода Alexandromys ЯОР исследованы только для A. fortis (номера пар хромосом несущих ЯОР-блоки не определяли), у которого ЯОР локализованы в теломерном районе метацентрической пары и центромерном районе двух пар аутосом [36]. Наличие ЯОР-блоков в прицентромерном районе небольшой пары и самой мелкой акроцентрической пары набора эворонской полевки возможно соответствуют таковым для A. fortis.
Ранее для кариотипов полевок из долины оз. Эворон (точка № 5) были исследованы С-окрашенные хромосомы [2]. В работе сказано: “Аутосомный гетерохроматин у эворонской полевки очень слабо выражен. В основном – это центромерные блоки, далеко не всегда выявляющиеся у всех пар. На Х-хромосоме кроме центромерного блока присутствует еще околоцентромерный блок. Y-хромосома целиком состоит из гетерохроматина” [2, стр. 201]. Полевки берегов р. Амгунь имеют прицентромерный гетерохроматин во всех парах аутосом (рис. 3,б). Следовательно можно было сделать вывод, что полевки долин р. Амгунь и оз. Эворон различаются по локализации гетерохроматина. Однако у исследованной нами полевки с берегов Эворон (рис. 3,д) мы отмечаем наличие прицентромерных блоков во всех парах аутосом, а также различное гетерохроматиновое окрашивание гетероморфных хромосом Mev17/18M и Mev17/18А. Также отчетливо виден околоцентромерный блок в длинном плече третьей пары аутосом. Поэтому считаем, что наши данные скорее дополняют информацию о характере окрашивания на структурный гетерохроматин в хромосомах этого вида, а не служат отличительными особенностями кариотипов полевки р. Амгунь и оз. Эворон. Следует отметить, что дифференциальное окрашивание хромосом полевок, приготовленных из костного мозга взрослых животных, имеет проблемы, поскольку цитоплазма клеток костного мозга у таких особей имеет много жировых включений, что мешает получению качественных препаратов.
Авторами, описавшими этот вид [9], было высказано предположение, что теоретически диплоидное число у эворонской полевки может варьировать от 36 до 42. Однако на сегодняшний день данные по известным для вида перестройкам в Эворон-Чукчагирской популяции не позволяют допустить наличия здесь кариотипов с 2n = 36–37. Тем не менее обнаруженные нами ранее особи из Верхнебуреинской впадины Хабаровского края и Верхнезейской равнины Амурской обл. имели в кариотипе 37 и 36 хромосом соответственно [25, 26] свидетельствуют о том, что на сегодняшний день эти числа выходят за пределы обнаруженной изменчивости вида. Возможно для эворонской полевки можно предположить наличие двух хромосомных рас: “эворон” с 2n = 38–42 и “арги” c 2n = 36, 37. В настоящее время характер хромосомного полиморфизма расы “арги” не исследован, однако большое число пар, вступающих в перестройки и “легкость”, с которой происходят слияния хромосом у полевок Эворон-Чукчагирской популяции, позволяет предположить обнаружения у этого вида перестроек с участием как аналогичных, так и других пар аутосом.
Исследования характера тандемных перестроек у животных из природных популяций представляют большой интерес, так как показано, что транслокации и тандемные слияния лежали в основе преобразования кариотипов многих групп млекопитающих [37–39]. Наиболее яркими примерами служат тандемные слияния хромосом, сопровождавшие видообразования оленей рода Muntiacus [40, 41] и хлопковых крыс рода Sigmodon [42, 43].
Для серых полевок рода Microtus (в который до недавнего времени входили восточноазиатские полевки в ранге подрода Alexantromys) показана быстрая дивергенция кариотипа [44–46]. Бóльшая часть хромосомных перестроек приходилась на робертсоновские слияния, но и тандемные слияния также не были редкими событиями. Однако под тандемными слияниями как правило подразумевались слияния двуплечего элемента с одноплечим (TCen) [44, 45, 47].
Известно, что во внутри- и межпопуляционный полиморфизм млекопитающих, особенно у эволюционно молодых видов, могут быть вовлечены все известные хромосомные перестройки, однако самым “вредным” из них являются тандемные слияния [39, 48]. Нам известны тандемные слияния в гетерозиготном состоянии для нескольких видов млекопитающих. Так у летучих мышей Uroderma bilobatum [49] перестройка выявлена в гетерозиготном состоянии у 9% особей, у южно-американского грызуна туко-туко Ctenomys talarum – 11% [50], и у одной особи из восьми исследованных в Европе мышовок Sicista subtilis [51]. Однако исследование мейоза Ctenomys talarum допускает, что изменчивость хромосом связана не с тандемным, а с робертсоновским слиянием хромосом [52]. Тандемное слияние двуплечих хромосом с образованием крупного двуплечего элемента показано также для трех видов млекопитающих: песцов Anopex lagopus [53] и двух видов полевок Alexandromys maximowiczii [2, 10, 13], A. evoronensis [2], однако частота такой перестройки для этих видов не была определена. Наше исследование выявило гетерозиготное состояние тандемного слияния (TTel) с частотой 0.47 для особей природной популяции A. evoronensis в Эворон-Чукчагирской низменности.
Принято считать, что теломеры представляют собой нуклеопротеиновые структуры, расположенные на концах линейных эукариотических хромосом и защищающие их от деградации, слияния и активности системы репарации ДНК [54–57]. Обнаружение тандемного слияния хромосом (TTel) в гетерозиготном состоянии в природной популяции эворонской полевки представляет интерес для дальнейшего исследования с целью понимания системы преобразования ДНК в теломерных районах во время хромосомных перестроек.
Благодарим Т.В. Васильеву за помощь в кариотипировании полевок лабораторной популяции.
Работа выполнена при финансовой поддержке РФФИ (12-04-00662а, 15-04-03871).
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Поздняков А.А. Морфотипическая изменчивость жевательной поверхности коренных зубов серых полевок группы “maximowiczii” (Rodentia, Arvicolidae, Microtus): опыт количественного статистического анализа // Зоол. журн. 1993. Т. 72. № 11. С. 114–125.
Мейер М.Н., Голенищев Ф.Н., Раджабли С.И., Саблина О.Л. Серые полевки фауны России и сопредельных территорий. СПб.: Зоол. ин-т РАН, 1996. 320 с.
Лисовский А.А., Оболенская Е.В. Структура краниометрического разнообразия серых полевок Microtus подрода Alexandromys // Тр. Зоол. ин-та РАН. 2011. Т. 315. № 4. С. 461–477.
Войта Л.Л., Голенищев Ф.Н., Тиунов М.П. Дальневосточные полевки Alexandromys (Rodentia: Cricetidae) из позднеплейстоценовых и голоценовых отложений пещеры Медвежий клык Приморского края России // Тр. Зоол. ин-та РАН. 2019. Т. 323. № 3. С. 313–346.
Фрисман Л.В., Коробицына К.В., Картавцева И.В. и др. Серые полевки (Microtus Shrank, 1798) Дальнего Востока России: аллозимная и кариологическая дивергенция // Генетика. 2009. Т. 45. № 6. С. 804–812.
Lemskaya N.A., Romanenko S.A., Golenishchev F.N. et al. Chromosomal evolution of Arvicolinae (Cricetidae, Rodentia). III. Karyotype relationships of ten Microtus species // Chromosome Res. 2010. V. 18. P. 459–471. https://doi.org/10.1007/s10577-010-9124-0
Ковальская Ю.М. Хромосомный полиморфизм Microtus maximowiczii Schrenck, 1858 (Rodentia, Cricetidae) // Бюлл. Моск. о-ва испытателей природы. 1977. Т. 82. № 2. С. 38–48.
Орлов В.Н., Булатова Н.Ш. Сравнительная цитогенетика и кариосистематика млекопитающих. М.: Наука, 1983. 406 с.
Ковальская Ю.М., Соколов В.Е. Новый вид полевок (Rodentia, Cricetidae, Microtus) из нижнего Приамурья // Зоол. журн. 1980. Т. 59. № 9. С. 1409−1416.
Kartavtseva I.V., Sheremetyeva I.N., Korobitsina K.V. et al. Chromosomal forms of Microtus maximowiczii (Schrenck, 1858) (Rodentia, Cricetidae): Variability in 2n and NF in different geographic regions // Russ. J. Theriology. 2008. V. 7. № 2. P. 89–97.
Harada M., Ando A., Lin L.-K., Takada S. Karyotypes of Taiwan vole Microtus kikuchii and the Pere David’s vole Eothenomys melanogaster from Taiwan // J. Mammalogical Society of Japan. 1991. V. № 16. P. 41–45.
Мейер М.Н., Волобуев В.Г. Морфологические особенности и хромосомный набор сахалинской полевки – Microtus sachalinensis Vasin, 1955, Rodentia, Cricetidae // Тр. Биолого-почвенного ин-та ДВНЦ АН СССР. 1974. Т. 17. № 120. С. 75–83.
Sheremetyeva I.N., Kartavtseva I.V., Voyta L.L. et al. Morphometric analysis of intraspecific variation in Microtus maximowiczii (Rodentia, Cricetidae) in relation to chromosomal differentiation with reinstatement of Microtus gromovi Vorontsov, Boeskorov, Lyapunova et Revin, 1988, stat. nov. // J. Zool. Syst. and Evol. Res. 2009. V. 47. № 1. P. 42–48.https://doi.org/10.1111/j.1439-0469.2008.00511.x
Fredga K., Bergstrem U. Chromosome polymorphism in the root vole (Microtus oeconomus) // Hereditas. 1970. V. 66. № 1. P. 145–152. https://doi.org/10.1111/j.1601-5223.1970.tb02340.x
Баскевич М.И., Хляп Л.А., Шварц Е.А. Хромосомный полиморфизм в периферической популяции полевки экономки Microtus oeconomus (Rodentia, Arvicolinae) из Валдайского национального парка // Докл. РАН. 2014. Т. 454. № 2. С. 227–231.
Kyoya T., Obara Y., Nakata A. Chromosomal aberrations in Japanese voles in and around an illegal dumpsite at the Aomori-Iwate prefectural boundary // Zool. Sci. 2008. V. 25. № 3. P. 307–312. https://doi.org/10.2108/zsj.25.30
Kartavtseva I.V., Kryukov A.P. Karyotype of Microtus fortis (Rodentia, Cricetidae) from extreme south of Far East Russia // Chromosome Sci. 1998. V. 2. P. 31–34.
Couranta F., Brunet-Lecomtea P., Volobouev V. et al. Karyological and dental identification of Microtus limnophilus in a large focus of alveolar echinococcosis (Gansu, China) // Biol. et Pathol. Animals. 1999. V. 322. P. 473–480. https://doi.org/10.1016/S0764-4469(99)80097-3
Картавцева И.В., Васильева Т.В., Шереметьева И.Н. и др. Генетическая изменчивость трех изолированных популяций муйской полевки Alexandromys mujanensis Orlovet Kovalskaja, 1978 (Rodentia, Arvicolinae) // Генетика. 2019. Т. 55. № 8. С. 920–935. https://doi.org/10.1134/S0016675819080071
Lissovsky A.A., Petrova T.V., Yatsentyuk S.P. et al. Multilocus phylogeny and taxonomy of East Asian voles Alexandromys (Rodentia, Arvicolinae) // Zool. Scripta. 2018. V. 47. № 1. P. 9–20. https://doi.org/10.1111/zsc.12261
Картавцева И.В., Шереметьева И.Н., Романенко С.А., Гладких О.Л. Изменчивость хромосом полевки Максимовича Microtus maximowiczii (Rodentia, Cricetidae, Microtus) // Цитология. 2013. Т. 55. № 4. С. 261–263.
Голенищев Ф.Н., Раджабли С.И. Новый вид серых полевок с берегов озера Эворон // Докл. АН СССР. 1981. Т. 257. № 1. С. 248–250.
Шереметьева И.Н., Картавцева И.В., Войта Л.Л., Тиунов M.П. Новые данные по распространению серых полевок рода Microtus (Rodentia: Cricetidae) на Дальнем Востоке России // Зоол. журн. 2010. Т. 89. № 10. С. 1273–1276.
Картавцева И.В., Шереметьева И.Н., Немкова Г.А., Лазурченко Е.В. Хромосомные исследования полевки Максимовича Microtus maximowiczii Schrenk, 1858 в Норском заповеднике Амурской области и эворонской Microtus evoronensis Kovalsk. et Socolov, 1980 окрестностей озера Эворон Хабаровского края // Териофауна России и сопредельных территорий. М.: Товарищество научных изданий КМК, 2007. 188 с.
Sheremetyeva I.N., Kartavtseva I.V., Vasiljeva T.V., Frisman L.V. Voles of the genus Alexandromys from the Verkhnebureinskaya Depression // Biol. Bull. 2017. V. 44. № 7. P. 813–819.https://doi.org/10.1134/S1062359017070159
Шереметьева И.Н., Картавцева И.В., Васильева Т.В. Обитает ли Эворонская полевка (Alexandromys evoronensis) на северо-востоке Верхнезейской равнины? // Зоол. журн. 2016. № 2. С. 477–484.
Kartavtseva I.V., Sheremetyeva I.N., Pavlenko M.V. et al. A new chromosomal race of the Evoron voles Alexandromys evoronensis of two isolated populations in the Russian Far East // 6th International Conference of Rodent Biology and Management and 16th Rodents et Spatium (3–7 September, 2018, Potsdam, Germany): Book of abstracts. 2018. P. 268.
Haring E., Sheremetyeva I., Kryukov A. Phylogeny of Palearctic vole species (genus Microtus, Rodentia) based on mitochondrial sequences // Mammalian Biol. 2011.V. 76. P. 258−267.https://doi.org/10.1016/j.mambio.2010.04.006
Павленко М.В., Шереметьева И.Н., Васильева Т.В. Особенности содержания и размножения в лабораторных условиях серых полевок рода Alexandromys Ognev, 1914 из популяции неясного таксономического статуса северо-востока Верхнезейской равнины // Вестн. Иркутской гос. с.-х. акад. 2017. Т. 83. С. 120–125.
Ford C.F., Hamerton J.L. A colchicine hypotonic citrate squash preparation for mammalian chromosomes // Stain Technol. 1956. V. 31. P. 247–251.
Графодатский А.А., Раджабли С.И. Хромосомы сельскохозяйственных и лабораторных млекопитающих. Атлас. Новосибирск: Наука, 1988. 128 с.
Seabright M. A rapid banding technique for human chromosomes // Lancet. 1971. V. 11. № 7731. P. 971–972.
Sumner A.T. A simple technique for demonstrating centromeric heterochromatin // Exp. Cell Res. 1972. V. 83. P. 438–442.
Miinke M., Schmiady H. A simple one-step for staining the nucleolus organizer region // Experientia. 1979. V. 35. P. 602–603.
Sablina O.V., Radjabli S.I., Graphodatsky A.S. Selected karyotypes // Atlas of Mammalian Karyotypes / O’Brien S.J., Nash W.G., Menninger J.C. 2006. P. 267.
Шереметьева И.Н., Картавцева И.Н., Фрисман Л.В. Кариологическая и аллозимная изменчивость дальневосточной полевки (Microtus fortis, Buchner, 1889 Cricetidae, Rodentia) Дальнего Востока России // Генетика. 2006. Т. 42. № 6. С. 833–843.
Kulemzina A.I., Trifonov V.A., Perelman P.L. et al. Cross-species chromosome painting in Cetartiodactyla: Reconstructing the karyotype evolution in key phylogenetic lineages // Chrom. Res. 2009 V. 17. P. 419–436. https://doi.org/10.1007/s10577-009-9032-3
Elder F.F.B., Hsu T.C. Tandem fusions in the evolution of mammalian chromosomes // The Cytogenetics of Mammalian Autosomal Rearrangements / Ed. Sandberg A.A. N.Y.: Alan R. Liss. 1988. P. 481–506.
Dobigny G., Britton-Davidian J., Robinson T.J. Chromosomal polymorphism in mammals: An evolutionary perspective // Biol. Rev. 2017. V. 92. P. 1–21. https://doi.org/10.1111/brv.12213
Liming S., Yingying Y., Xingsheng D. Comparative cytogenetic studies on the red muntjac, Chinese muntjac, and their F1 hybrids // Cytogen. and Cell Genet. 1980. V. 26. P. 22–27.
Huang L., Wang J., Nie W., Su W., Yang F. Tandem chromosome fusions in karyotypic evolution of Muntiacus: Evidence from M. feae and M. gongshanensis // Chromosome Res. 2006. V. 14. P. 637–647.
Elder F.F.B. Tandem fusion, centric fusion, and chromosomal evolution in the cotton rats, genus Sigmodon // Cytogen. Cell Genet. 1980. V. 26. P. 199–210.
Swier V.J., Bradley R.D., Rens W. et al. Patterns of chromosomal evolution in Sigmodon, evidence from whole chromosome paints // Cytogenet. Genome Res. 2009. V. 125. P. 54–66.
Modi W.S. Phylogenetic analyses of chromosomal banding patterns among the Nearctic Arvicolidae (Mammalia, Rodentia) // Syst. Zool. 1987. V. 36. № 2. P. 109–136.
Sitnikova N.A., Romanenko S.A., O’Brien P.C.M. et al. Chromosomal evolution of Arvicolinae (Cricetidae, Rodentia). I. The genome homology of tundra vole, field vole, mouse and golden hamster revealed by comparative chromosome painting // Chromosome Res. 2007. V. 15. P. 447–456.
Lemskaya N.A., Romanenko S.A., Golenishchev F.N. et al. Chromosomal evolution of Arvicolinae (Cricetidae, Rodentia). III. Karyotype relationships of ten Microtus species // Chromosome Res. 2010. V. 18. P. 459–471.https://doi.org/10.1007/s10577-010-9124-0
Mazurok N.A., Rubtsova N.V., Isaenko A.A. et al. 2001. Comparative chromosome and mitochondrial DNA analyses and phylogenetic relationships within common voles (Microtus, Arvicolidae) // Chromosome Res. V. 9. P. 107–120.
King M. Chromosomal Speciation Revisited (Again). Species Evolution. The Role of Chromosome Change Cambridge: Cambridge Univ.Press, 1993. 336 p.
Owen J.G., Baker R.K. The Uroderma bilobatum (Chiroptera, Phyllostomidae) cline revisited // J. Mammalogy. 2001. V. 82. № 4. P. 1102–1113. https://doi.org/10.1644/1545-1542(2001)082<1102:TU-BCPC>2.0.CO;2
Massarini A., Mizrahi D., Tiranti S. et al. Extensive chromosomal variation in Ctenomys talarum from the Atlantic coast of Buenos Aires province, Argentina (Rodentia, Octodontidae) // J. Neotropical Mammalogy. 2002. V. 9. P. 199–207.
Kovalskaya Y., Aniskin V., Bogomolov P. et al. Karyotype reorganization in the subtilis group of Birch Mice (Rodentia, Dipodidae, Sicista): Unexpected taxonomic diversity within a limited distribution // Cytogen. and Genome Res. 2011. V. 132. P. 271–288. https://doi.org/10.1159/000322823
Basheva E.A., Torgasheva A.A., Gomez Fernandez M.J. et al. Chromosome synapsis and recombination in simple and complex chromosomal heterozygotes of tuco-tuco (Ctenomys talarum: Rodentia: Ctenomyidae) // Chromosome Res. 2014. V. 22. № 3. P. 351–363. https://doi.org/10.1007/s10577-014-9429-5
Раджабли С.И., Графодатский А.С. Эволюция кариотипа млекопитающих. Структурные перестройки хромосом и гетерохроматин // Цитогенетика гибридов, мутаций и эволюция кариотипа. Новосибирск: Наука, 1977. С. 231–249.
Garcia-Cao M., O’Sullivan R., Peters A.H. et al. Epigenetic regulation of telomere length in mammalian cells by the Suv39h1 and Suv39h2 histone methyltransferases // Nat. Genet. 2004. V. 36. P. 94–99. https://doi.org/10.1038/ng1278
Benetti R., Garcia-Cao M., Blasco M.A. Telomere length regulates the epigenetic status of mammalian telomeres and subtelomeres // Nat. Genet. 2007. V. 39. P. 243–250.https://doi.org/10.1038/ng1952
Palm W., de Lange T. How shelterin protect mammalian telomeres // Annu. Rev. Genet. 2008. V. 42. P. 301–334. https://doi.org/10.1146/annurev.genet.41.110306.130350
Кордюкова М.Ю., Калмыкова А.И. Природа и функции теломерных транскриптов // Биохимия. 2019. Т. 84. № 2. С. 212–222.https://doi.org/10.1134/S0320972519020040
Дополнительные материалы отсутствуют.