

Генетика, 2021, T. 57, № 4, стр. 420-428
Оценка генетического разнообразия кыргызской породы лошадей с использованием микросателлитных маркеров – расширенное геногеографическое исследование
Ж. Т. Исакова 1, *, М. А. Исаев 2, В. Н. Кипень 3, Л. В. Калинкова 4, К. А. Айтбаев 1, М. А. Арзыбаев 2, С. Б. Мукеева 1, Осмонкул кызы Мээрим 1, Н. М. Алдашева 1
1 Институт молекулярной биологии и медицины
720040 Бишкек, Кыргызстан
2 Кыргызский национальный аграрный университет им. К.И. Скрябина
720005 Бишкек, Кыргызстан
3 Институт генетики и цитологии Национальной академии наук Беларуси
220072 Минск, Республика Беларусь
4 Всероссийский научно-исследовательский институт коневодства
391105 п. Дивово, Рязанская обл., Россия
* E-mail: jainagul@mail.ru
Поступила в редакцию 29.04.2020
После доработки 07.08.2020
Принята к публикации 01.10.2020
Аннотация
Современная порода кыргызских лошадей обнаруживает высокий уровень внутрипородной генетической вариабельности. В 17 STR-локусах идентифицировано 154 аллеля, в том числе 59 редких аллелей. Выявлены новые редкие аллели для 15 локусов – AHT4, AHT5, ASB2, ASB17, ASB23, CA425, HMS1, HMS2, HMS3, HMS6, HMS7, HTG4, HTG6, HTG7, HTG10. Наибольшее генетическое разнообразие среди четырех исследованных географически изолированных зон в Таласской, Ошской и Нарынской областях отмечено среди лошадей кыргызской породы из села Таш-Башат (Нарынская обл.): среднее количество выявленных аллелей было максимальным среди исследованных групп 8.294 ± 0.561, количество эффективных аллелей – 5.009 ± 0.416, индекс разнообразия Шеннона (I) – 1.745 ± 0.076 и ожидаемая гетерозиготность (He) – 0.773 ± 0.020.
Кыргызская порода лошадей имеет многосторонний характер хозяйственного использования. Они широко используются в государственных, коллективных, фермерских, крестьянских, а также личных подсобных хозяйствах населения в качестве живой тягловой силы при выполнении разнообразных видов транспортных и сельскохозяйственных работ [1, 2].
Кыргызская лошадь отличается комбинированными продуктивными качествами и хорошей приспособленностью к условиям табунного содержания, что следует рассматривать как положительный фактор при организации мясного и молочного коневодства. Кочевой образ жизни, экстенсивное ведение табунного коневодства препятствовали созданию статной быстроаллюрной лошади. Особи данной породы не характеризуются большой резвостью, но отличается легкостью и хорошей координацией движений, исключительно выносливы и неприхотливы к кормам; они мало восприимчивы к резким переменам погоды, обладают хорошей дистанционностью при работе под седлом.
Перспективным направлением является продуктивное коневодство, которое подразделяется на мясное табунное и молочное. Молочное коневодство предусматривает получение кобыльего молока и производство из него высокоценного пищевого диетического и лечебного продукта – кумыса. Натуральный кумыс, приготовленный из кобыльего молока, является традиционным продуктом питания населения Кыргызстана. В современной медицине кумыс применяется не только для лечения туберкулеза легких, но и желудочно-кишечных, костных и ряда других заболеваний [2].
Племенные коневодческие хозяйства Кыргызстана занимаются разведением племенных лошадей для улучшения массового коневодства (рабоче-пользовательного и продуктивного) и спортивных лошадей для конно-спортивных организаций различного назначения (конноспортивные школы, секции и клубы, пункты верхового и экипажного проката и др.).
В Кыргызстане планируется внедрить новые виды спорта для выхода на международную арену – конные пробеги. Вместе с тем, в силу ряда причин, многогранные резервы коневодства в увеличении производства сельскохозяйственной продукции используются не полностью, а коневодство отстает от мирового уровня. Национальная порода – кыргызская лошадь, пройдя через века, в XX и XXI столетиях оказалась на грани почти полного вырождения, про нее встречаются только отрывочные сведения в научной литературе, а сама же она рассеялась, смешавшись с разными породами. Для сохранения ценных особенностей генофонда кыргызской лошади необходимо проводить генетические исследования, а сохранение и дальнейшее совершенствование породы должно осуществляться под контролем изучения генетической ситуации как в породе в целом, так и в основных племенных хозяйствах, занимающихся разведением чистопородных лошадей.
В настоящее время наиболее эффективными генетическими маркерами для описания генетической структуры популяций разных видов животных, в частности лошадей, являются полиморфные микросателитные локусы ДНК (STR, Short Tandem Repeat), которые имеют кодоминантный характер наследования и служат незаменимым инструментом при исследовании генетических различий не только между животными, но и популяциями одной породы, а также между породами.
Цель исследования – провести расширенное геногеографическое исследование, а также дать оценку генетической структуры и генетического разнообразия кыргызской породы лошадей с использованием 17-ти микросателлитных маркеров.
МАТЕРИАЛЫ И МЕТОДЫ
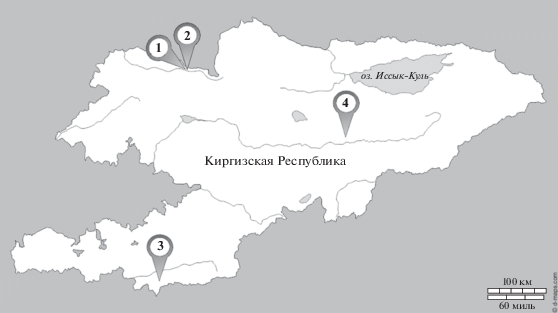
Биологическим материалом для молекулярно-генетического исследования послужили образцы волос с луковицами, взятые у взрослого поголовья кыргызских лошадей (104 животных), разводимых в высокогорных экспериментальных зонах: в селах Копуро-Базар (№ 1) и Талды-Булак (№ 2) (Таласский р-н, Таласская обл.) – по 20 животных, в с. Чон-Алай (Чон-Алайский р-н, Ошская обл.) – 23 животных (№ 3) и в с. Таш-Башат (Нарынский р-н, Нарынская обл.) – 41 животное (№ 4). Места отбора образцов представлены на рис. 1.
Рис. 1.
Места отбора образцов: 1 – с. Копуро-Базар, Таласский р-н, Таласская обл.; 2 – с. Талды-Булак, Таласский р-н, Таласская обл.; 3 – с. Чон-Алай, Чон-Алайский р-н, Ошская обл.; 4 – с. Таш-Башат, Нарынский р-н, Нарынская обл.
Лошадей для молекулярно-генетического исследования отбирали по признакам экстерьера: рост, масть и фенотип. Для кыргызских лошадей характерны сравнительно большая голова, недлинная шея, низкая холка, прямая спина, спущенный, часто короткий круп, широкая и глубокая грудь и сухие ноги. Особо следует отметить высокую прочность копытного рога, позволяющего передвигаться по каменистому грунту даже с тяжелым грузом на спине. Масть кыргызских лошадей в большинстве случаев гнедая или серая, бывают пегие.
Образцы были генотипированы по 17 микросателлитным локусам, рекомендованным Международным обществом генетики животных (ISAG, International Society for Animal Genetics): AHT4, AHT5, ASB2, ASB17, ASB23, CA425, HMS1, HMS2, HMS3, HMS6, HMS7, HTG4, HTG6, HTG7, HTG10, LEX3, VHL20. Для обозначения аллелей применялся международный алфавитный код [3]. Статистическую обработку данных проводили с использованием программ GenALEx v. 6.5 [4], STRUCTURE v. 2.3.4 [5] и POPHELPER v. 1.0.10 [6]. С использованием GenALEx v. 6.5 были рассчитаны следующие показатели: среднее число аллелей на локус (Na), эффективное число аллелей (Ne), уровни ожидаемой (He) и наблюдаемой (Ho) гетерозиготностей, значение информационного индекса Шеннона (I), коэффициент FIS [7]. В программе STRUCTURE v. 2.3.4 по методу J.K. Pritchard с соавт. был рассчитан критерий Q, который характеризует принадлежность каждого отдельного животного к соответствующему кластеру. С использованием веб-приложения POPHELPER v. 1.0.10 произведена графическая интерпретация результатов, полученных в STRUCTURE v. 2.3.4.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Современная порода кыргызских лошадей обнаруживает высокий уровень внутрипородной генетической вариабельности – в 17-ти исследованных нами микросателлитных локусах было идентифицировано 154 аллеля, в том числе 59 редких аллелей (с частотой встречаемости менее 5.0%), что составляет 38.3% от общего количества выявленных аллелей: L/M/N/P для локуса AHT4, I – для AHT5, B/C/I/P – для ASB2, D/H/J/K/L/P/Q/T/W – для ASB17, G/M (21 динуклеотидный повтор)/Q/R – для ASB23, F/G/H/I/K – для CA425, N/Q – для HMS1, J/M/O/P/R – для HMS2, G (19 динуклеотидных повторов)/L/S – для HMS3, N/Q – для HMS6, P/Q – для HMS7, J (29 динуклеотидных повторов)/N/P – для HTG4, M/N/P/R (24 динуклеотидных повтора) – для HTG6, L (16 динуклеотидных повторов) – для HTG7, N/P/Q/S/T – для HTG10, G/I/K/O – для LEX3, J – для VHL20 (табл. 1). Также выявлены 22 новых аллеля (14.3% от общего количества всех аллелей), не обнаруженных в нашем предыдущем исследовании [8]. Из них для 15 локусов – AHT4, AHT5, ASB2, ASB17, ASB23, CA425, HMS1, HMS2, HMS3, HMS6, HMS7, HTG4, HTG6, HTG7, HTG10 – имеются новые редкие аллели с частотой встречаемости менее 5.0% (табл. 1).
Таблица 1.
Частоты распространенности аллелей в 17 микросателлитных локусах ДНК для лошадей кыргызской породы
| Локус | Аллели | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| B | C | D | F | G | H | I | J | K | L | |
| AHT4 | 0.202 | 0.058 | 0.149 | 0.144 | 0.043* | |||||
| AHT5 | 0.005 | 0.188 | 0.260 | 0.091 | ||||||
| ASB2 | 0.019 | 0.010 | 0.010 | 0.159 | ||||||
| ASB17 | 0.005 | 0.038 | 0.096 | 0.038 | 0.091 | 0.019 | 0.043 | 0.034 | ||
| ASB23 | 0.010 | 0.072 | 0.245 | 0.173 | 0.173 | |||||
| CA425 | 0.005 | 0.024 | 0.005 | 0.024 | 0.087 | 0.014 | 0.077 | |||
| HMS1 | 0.063 | 0.346 | 0.067 | 0.053 | ||||||
| HMS2 | 0.293 | 0.130 | 0.014 | 0.173 | 0.269 | |||||
| HMS3 | 0.005# | 0.096 | 0.005 | |||||||
| HMS6 | 0.053 | 0.101 | ||||||||
| HMS7 | 0.111 | 0.053 | 0.341 | |||||||
| HTG4 | 0.005 | 0.163 | 0.115 | |||||||
| HTG6 | 0.188 | 0.058 | 0.154 | |||||||
| HTG7 | 0.284 | 0.005 | ||||||||
| HTG10 | 0.063 | 0.077 | 0.072 | |||||||
| LEX3 | 0.159 | 0.005 | 0.144 | 0.038 | 0.029 | 0.207 | ||||
| VHL20 | 0.135 | 0.024 | 0.067 | |||||||
| Локус | Аллели | |||||||||
| M | N | O | P | Q | R | S | T | U | W | |
| AHT4 | 0.019 | 0.024 | 0.322 | 0.038 | ||||||
| AHT5 | 0.091 | 0.197 | 0.168 | |||||||
| ASB2 | 0.178 | 0.308 | 0.135 | 0.024 | 0.096 | 0.063 | ||||
| ASB17 | 0.063 | 0.163 | 0.077 | 0.010 | 0.034 | 0.207 | 0.053 | 0.024 | 0.005 | |
| ASB23 | 0.005 | 0.005 | 0.043 | 0.149 | 0.014 | 0.125 | ||||
| CA425 | 0.250 | 0.394 | 0.120 | |||||||
| HMS1 | 0.442 | 0.014 | 0.014 | |||||||
| HMS2 | 0.048 | 0.014 | 0.010 | 0.048 | ||||||
| HMS3 | 0.207 | 0.091 | 0.120 | 0.317 | 0.058 | 0.087 | 0.014 | |||
| HMS6 | 0.264 | 0.024 | 0.188 | 0.361 | 0.010 | |||||
| HMS7 | 0.115 | 0.149 | 0.197 | 0.005 | 0.029 | |||||
| HTG4 | 0.591 | 0.034 | 0.053 | 0.038 | ||||||
| HTG6 | 0.034 | 0.005 | 0.529 | 0.029 | 0.005 | |||||
| HTG7 | 0.144 | 0.216 | 0.351 | |||||||
| HTG10 | 0.130 | 0.019 | 0.260 | 0.034 | 0.048 | 0.245 | 0.048 | 0.005 | ||
| LEX3 | 0.207 | 0.077 | 0.048 | 0.087 | ||||||
| VHL20 | 0.221 | 0.111 | 0.120 | 0.101 | 0.091 | 0.130 | ||||
Примечание. * – аллели с частотой распространенности <5.0% (редкий аллель); # – новые аллели, не представленные в https://strbase.nist.gov/horseSTRs.htm.
Значение индекса Шеннона для данной породы составляет 1.749 ± 0.066, что указывает на среднюю сложность структуры сообщества исследованных кыргызских лошадей [2].
Для оценки генетической подразделенности исследуемых групп лошадей кыргызской породы в программе STRUCTURE v. 2.3.4 по методу J.K. Pritchard был проведен расчет критерия Q, который характеризует принадлежность каждого отдельного животного к соответствующей группе. Значение Q, равное 75% или выше, подтверждает членство особи в своем кластере. На рис. 2 графически представлены (с использованием веб-приложения POPHELPER v. 1.0.10 [http://pophelper.com/]) результаты анализа, проведенного в STRUCTURE v. 2.3.4 (проведена автоматическая сортировка на основании принадлежности конкретного образца к мажорному кластеру).
Рис. 2.
Результаты анализа генетической структуры изучаемых четырех выборок кыргызской породы лошадей для наиболее вероятного числа кластеров (K) от 3 до 10 (ось X – ID животного, ось Y – доля членства в соответствующем кластере).
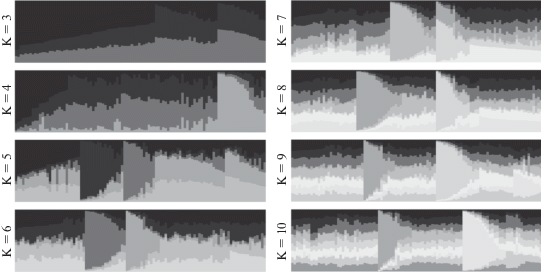
В исследовании был использован генетический материал лошадей из четырех географически изолированных высокогорных зон (рис. 1). Уже для K = 5 выделяется три четких кластера, и данная зависимость сохраняется вплоть до значения K = 10. Для всех образцов за пределами данных кластеров наблюдается общая однородность структуры, вклад каждого субкластера становится равноценным. На рис. 3 представлены результаты кластеризации для всех 104 образцов (расположены по порядку, без сортировки по значению Q) в пределах географических зон (K = 4).
Рис. 3.
Результаты анализа генетической структуры изучаемых четырех выборок кыргызской породы лошадей для K = 4 (четыре выборки): № 1 – с. Копуро-Базар, Таласский р-н, Таласская обл.; № 2 – с. Талды-Булак, Таласский р-н, Таласская обл.; № 3 – с. Чон-Алай, Чон-Алайский р-н, Ошская обл.; № 4 – с. Таш-Башат, Нарынский р-н, Нарынская обл. (ось X – ID животного, ось Y – доля членства в соответствующем кластере).
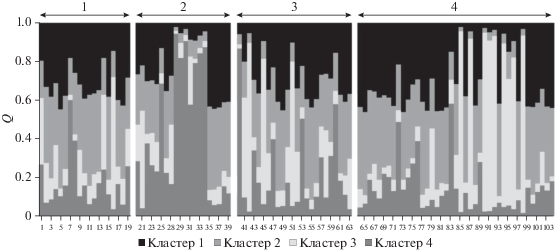
Видно, что для каждой группы (№ 1–4 на рис. 3) имеются особенности: в пределах групп лошадей из с. Талды-Булак (Таласский р-н, Таласская обл.) – № 2 – имеются особи, относящиеся к одному субкластеру (цвет кластера соответствует Cluster 2 на рис. 3); некоторые особи кыргызской породы лошадей из с. Чон-Алай (Чон-Алайский р‑н, Ошская обл.) – № 3, и с. Таш-Башат (Нарынский р-н, Нарынская обл.) – № 4 – также могут быть сгруппированы в один субкластер (цвет кластера соответствует Cluster 3 на рис. 3). Данный факт может быть следствием того, что изучаемые в рамках данного исследования субпопуляции лошадей имеют общих предков (например производителей из других табунов), возможно влияние иных факторов.
На основании анализа генетических дистанций FST, рассчитанных по алгоритму AMOVA для 16 STR-локусов (кроме LEX3), был построен график главных компонент (PCA, principal component analysis), отражающий взаимное сходство или различие исследованных групп лошадей (рис. 4).

Видно, что группа особей из с. Талды-Булак, № 2 (Таласский р-н, Таласская обл.) расположена наиболее удаленно от других исследуемых групп лошадей. Также, несмотря на географическую близость расположения сел Копуро-Базар, № 1 (Таласский р-н, Таласская обл.) и Талды-Булак, № 2 (Таласский р-н, Таласская обл.), наблюдается существенное различие по взаимному расположению их координат на графике главных компонент, на долю которых (первая и вторая компоненты) приходится 97.07% вариации признака. Причина подобных несовпадений в большей степени обусловлена различиями в частоте распространенности аллелей в исследованных STR-локусах, а также наличием в исследованных группах лошадей как редких (частота распространенности менее 5%), так и приватных (встречаются только в одной из исследованных групп) аллелей – табл. 2 и 3 соответственно.
Таблица 2.
Суммарные частоты распространенности редких аллелей в исследованных группах лошадей кыргызской породы (в %)
| Локус | Группа | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| AHT4 | 7.50 | 5.00 | 10.87 | 7.32 |
| AHT5 | – | – | – | 6.10 |
| ASB2 | 2.50 | 2.50 | 4.35 | 7.32 |
| ASB17 | 10.00 | 5.00 | 30.44 | 20.73 |
| ASB23 | 5.00 | – | 2.17 | 1.22 |
| HMS1 | – | 2.50 | 21.74 | 10.98 |
| HMS2 | 2.50 | – | 6.52 | 15.85 |
| HMS3 | 2.50 | – | 17.39 | 2.44 |
| HMS6 | 5.00 | – | 4.35 | 9.76 |
| HTG4 | 2.50 | – | 6.52 | 3.66 |
| HTG6 | – | 2.50 | 2.17 | 1.22 |
| HTG7 | 2.50 | 2.50 | 2.17 | 13.42 |
| HTG10 | 5.00 | 2.50 | 4.35 | 3.66 |
| VHL20 | 2.50 | – | – | – |
| Сумма* | 55.00 | 30.00 | 126.09 | 124.40 |
Таблица 3.
Частоты распространенности приватных аллелей в исследованных группах лошадей кыргызской породы
| Группа | STR-локус | Аллель | Частота, % |
|---|---|---|---|
| 1 | ASB23 | G | 5.00 |
| ASB23 | Q | 2.50 | |
| HTG6 | N | 2.50 | |
| HTG7 | L | 2.50 | |
| 2 | ASB17 | W | 2.50 |
| HMS2 | P | 5.00 | |
| 3 | ASB17 | D | 2.20 |
| CA425 | H | 2.20 | |
| HMS6 | Q | 4.30 | |
| HTG6 | R | 2.20 | |
| 4 | AHT5 | I | 1.20 |
| ASB23 | M | 1.20 | |
| CA425 | F | 1.20 | |
| HMS1 | Q | 3.70 | |
| HMS3 | G | 1.20 | |
| HMS3 | L | 1.20 | |
| HMS7 | P | 1.20 | |
| HMS7 | Q | 7.30 | |
| HTG4 | J | 1.20 | |
| HTG10 | T | 1.20 |
Наибольший накопленный суммарный процент редких аллелей определен для лошадей из с. Чон-Алай (Чон-Алайский р-н, Ошская обл.) – № 4. Минимальное значение данного параметра выявлено для особей из с. Талды-Булак (Таласский р-н, Таласская обл.) – № 2. STR-локус ASB17 отличался наибольшим аллельным разнообразием среди всех исследованных групп. Также имелись и межгрупповые особенности: для локуса HMS2 среди особей из группы № 3 (с. Чон-Алай, Чон-Алайский р-н, Ошская обл.) на долю редких аллелей пришлось 17.39%; для локусов ABS17, HMS1, HTG4 и HTG10 среди особей из группы № 4 (с. Таш-Башат, Нарынский р-н, Нарынская обл.) на долю редких аллелей пришлось 20.73, 15.85, 13.42 и 12.20% соответственно. Для данных групп – № 3 и 4 – имелись также особенности и в количестве выявленных приватных аллелей. Среди лошадей из с. Чон-Алай они определены для четырех локусов – ASB17, CA425, HMS6 и HTG6 – причем для локуса HMS6 аллель Q выявлен в 4.30%. Среди лошадей из с. Таш-Башат приватные аллели выявлены уже для восьми локусов – AHT5, ASB23, CA425, HMS1, HMS3, HMS7, HTG4 и HTG10, – аллель Q для локуса HMS7 был выявлен в 7.30% случаев.
Из 16 STR-локусов – AHT4, AHT5, ASB2, ASB17, ASB23, HMS1, HMS2, HMS3, HMS6, HTG4, HTG6, HTG7, HTG10 и VHL20 – расположенных на аутосомах, наибольшим потенциалом по дифференциации исследуемых групп лошадей кыргызской породы обладают те, для которых рассчитанные значения FST являются максимальными – табл. 4.
Таблица 4.
Дифференцирующий потенциал 16 STR-локусов (результаты анализа locus-by-locus AMOVA)
| Локус | FST* | p-уровень | Локус | FST | p-уровень |
|---|---|---|---|---|---|
| AHT4 | 0.044 | <0.001 | HMS3 | 0.020 | <0.001 |
| AHT5 | 0.020 | <0.001 | HMS6 | 0.014 | <0.001 |
| ASB2 | 0.031 | <0.001 | HMS7 | 0.032 | <0.001 |
| ASB17 | 0.029 | <0.001 | HTG4 | 0.022 | <0.001 |
| ASB23 | 0.063 | <0.001 | HTG6 | 0.028 | <0.001 |
| CA425 | 0.032 | <0.001 | HTG7 | 0.010 | <0.001 |
| HMS1 | 0.031 | <0.001 | HTG10 | 0.026 | <0.001 |
| HMS2 | 0.035 | <0.001 | VHL20 | 0.021 | <0.001 |
Набольшие рассчитанные значения FST показаны для STR-локусов AHT4, ASB23 и HMS7, в целом значения FST для всех локусов невысокие и не превышают 0.1. Информация об аллельном разнообразии и частотах распространенности конкретных аллелей для перечисленных выше STR-локусов представлена в табл. 5.
Таблица 5.
Частоты распространенности аллелей для трех STR-локусов с наибольшим дифференцирующим потенциалом (по результатам FST)
| Локус/Аллели | Группа | |||
|---|---|---|---|---|
| AHT4 | 1 | 2 | 3 | 4 |
| H | 12.50 | 37.50 | 10.87 | 20.73 |
| I | 2.50 | 5.00 | 10.87 | 4.88 |
| J | 22.50 | 2.50 | 19.57 | 14.63 |
| K | 17.50 | 2.50 | 21.74 | 14.63 |
| L | 2.50 | 5.00 | 2.17 | 6.10 |
| M | 5.00 | – | 4.35 | – |
| N | 2.50 | – | 4.35 | 2.44 |
| O | 35.00 | 47.50 | 19.57 | 30.49 |
| P | – | – | 6.52 | 6.10 |
| ASB23 | 1 | 2 | 3 | 4 |
| G | 5.00 | – | 0.00 | 0.00 |
| I | 5.00 | – | 8.70 | 10.98 |
| J | 35.00 | 5.00 | 32.61 | 24.39 |
| K | 27.50 | 7.50 | 19.57 | 15.85 |
| L | 7.50 | 30.00 | 19.57 | 14.63 |
| M | – | – | – | 1.22 |
| Q | 2.50 | – | – | – |
| R | 2.50 | 5.00 | 2.17 | 6.10 |
| S | 5.00 | 42.50 | 6.52 | 10.98 |
| T | – | – | – | – |
| U | 10.00 | 10.00 | 10.87 | 15.85 |
| HMS7 | 1 | 2 | 3 | 4 |
| J | 17.50 | 2.50 | 17.39 | 8.54 |
| K | 7.50 | 5.00 | – | 7.32 |
| L | 37.50 | 22.50 | 32.61 | 39.02 |
| M | 15.00 | 25.00 | 2.17 | 8.54 |
| N | 5.00 | 15.00 | 21.74 | 15.85 |
| O | 17.50 | 30.00 | 26.09 | 12.20 |
| P | – | – | – | 1.22 |
| Q | – | – | – | 7.32 |
Таким образом, в различных группах превалируют конкретные аллели: в группе № 2 для локуса AHT4 в мажорном состоянии находятся аллели H и O, в то время как для остальных групп наибольшую распространенность получил только аллель O; для локуса ASB23 в группе № 2 суммарная частота распространенности аллелей L (30.0%) и S (42.5%) составила 72.5%, в то время как для других групп аллель S встречался с частотой менее 20.0%, для групп № 1, 3, 4 превалировал аллель J; для локуса HMS7 в группе № 2 мажорным (частота – более 30%) является аллель O, для групп же № 1, 3, 4 – аллель L. В целом для каждой группы имеются индивидуальные различия в профиле распределения частот распространенности аллелей по всем исследуемым STR-локусам.
Для анализа внутрипородной генетической подразделенности кыргызских лошадей, разводимых в четырех географически изолированных высокогорных зонах, необходимо также оценить значения показателей Na, Ne, Ho, He, I и коэффициента FIS – табл. 6.
Таблица 6.
Генетическая характеристика четырех субпопуляций лошадей кыргызской породы по результатам генотипирования 17 STR-локусов
| Группа | ||||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| Na | 7.059 ± 0.481 | 6.059 ± 0.441 | 7.471 ± 0.543 | 8.294 ± 0.561 |
| Ne | 4.556 ± 0.321 | 3.898 ± 0.318 | 4.837 ± 0.362 | 5.009 ± 0.416 |
| I | 1.643 ± 0.073 | 1.475 ± 0.083 | 1.695 ± 0.067 | 1.745 ± 0.076 |
| Ho# | 0.791 ± 0.023 | 0.759 ± 0.038 | 0.804 ± 0.020 | 0.765 ± 0.023 |
| He# | 0.756 ± 0.020 | 0.705 ± 0.029 | 0.773 ± 0.016 | 0.773 ± 0.020 |
| FIS# | –0.052 ± 0.031 | –0.075 ± 0.030 | –0.041 ± 0.016 | 0.009 ± 0.021 |
При анализе показателей Na, Ne, Ho, He, I и коэффициента FIS выявлено, что наименьшим генетическим разнообразием среди четырех географически изолированных зон отличаются лошади кыргызской породы из с. Талды-Булак (Таласский р-н, Таласская обл.) – № 2: среднее количество выявленных аллелей Na было минимальным среди исследованных групп – 6.059 ± 0.441, количество эффективных аллелей Ne – 3.898 ± 0.318, индекс разнообразия Шеннона I также оказался наименьшим среди рассчитанных – 1.475 ± 0.083, и ожидаемая гетерозиготность He также оказалась минимальной – 0.705 ± 0.029.
Наибольшее генетическое разнообразие было обнаружено среди лошадей кыргызской породы из с. Таш-Башат (Нарынский р-н, Нарынская обл.), № 4: Na – 8.294 ± 0.561, I – 1.745 ± 0.076.
ЗАКЛЮЧЕНИЕ
Современная порода кыргызских лошадей обнаруживает высокий уровень внутрипородной генетической вариабельности. В 17 STR-локусах идентифицировано 154 аллеля, в том числе 59 редких аллелей. Выявлены новые аллели, не обнаруженныe в нашем предыдущем исследовании [8], из них для 15 локусов – AHT4, AHT5, ASB2, ASB17, ASB23, CA425, HMS1, HMS2, HMS3, HMS6, HMS7, HTG4, HTG6, HTG7, HTG10, – имеются новые редкие.
При анализе показателей Na, Ne, Ho, He, I и коэффициента FIS выявлено, что наибольшее генетическое разнообразие среди четырех географически изолированных зон – с. Копуро-Базар (Таласский р-н, Таласская обл.), с. Талды-Булак (Таласский р-н, Таласская обл.), с. Чон-Алай (Чон-Алайский р-н, Ошская обл.) и с. Таш-Башат (Нарынский р-н, Нарынская обл.) – отмечено среди лошадей кыргызской породы из с. Таш-Башат: среднее количество выявленных аллелей Na было максимальным среди исследованных групп 8.294 ± ± 0.561, количество эффективных аллелей Ne – 5.009 ± 0.416, индекс разнообразия Шеннона I – 1.745 ± 0.076 и ожидаемая гетерозиготность He – 0.773 ± 0.020.
Исследование было поддержано Федерацией органического движения “БИО-KG” Кыргызстана (договор № 9 от 6 ноября 2019 г).
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Абрамзон С.М. Киргизы и их этногенетические и историко-культурные связи. Л.: Наука, 1971. 478 с.
Рипар Ж. Кыргызская лошадь: сохранение и использование, скачки на выносливость и экотуризм. Практическое указание / Под ред. Рипар Ж., Уатто С., Перез С., перевод Илиязова А. Бишкек: 2007. 32 с.
Van De Goor L.H.P. A proposal for standardization in forensic equine DNA typing: Allele nomenclature for 17 equine-specific STR loci // Anim. Genet. 2010. V. 41(2). P. 122–127. https://doi.org/10.1111/j.1365-2052.2009.01975.x
Peakall R., Peter E. Smouse. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research – an update // Bioinformatics. 2012. V. 28. P. 2537–2539. https://doi.org/10.1093/bioinformatics/bts460
Pritchard J.K., Stephens M., Donnelly P. Inference of population structure using multilocus genotype data // Genetics. 2000. V. 155(2). P. 945–959.
Hammer Q., Harper A.T., Ryan P.D. Past: Paleontological Statistics Software Package for Education and Data Analysis // Palaeontologia Electronica. 2001. V. 4. P. 1–9.
Excoffier L. Analysis of molecular variance inferred from metric distances among dna haplotypes: Application to human mitochondrial dna restriction data // Genetics. 1992. V. 131. P. 479–491.
Исакова Ж.Т., Токтосунов Б.И., Кипень В.Н. и др. Генетический портрет кыргызской лошади // Коневодство и конный спорт. 2018. № 1. С. 21–23.
Дополнительные материалы отсутствуют.