

Генетика, 2021, T. 57, № 4, стр. 437-444
Генетическая структура популяции голштинизированного черно-пестрого скота на территории Урала
М. В. Модоров 1, *, И. В. Ткаченко 1, А. А. Грин 1, М. Ю. Севостьянов 1, Н. Н. Зезин 1
1 Уральский федеральный аграрный научно-исследовательский центр
Уральского отделения Российской академии наук
620142 Екатеринбург, Россия
* E-mail: mmodorov@gmail.com
Поступила в редакцию 19.05.2020
После доработки 04.12.2020
Принята к публикации 10.12.2020
Аннотация
Проанализирована изменчивость 15 микросателлитных локусов у 15 786 особей крупного рогатого скота черно-пестрой, голштинской и тагильской пород, разводимых на территории Урала. Показано, что современная популяция голштинизированного черно-пестрого скота, содержащегося в племзаводах и племрепродукторах Свердловской, Курганской, Тюменской, Челябинской областей, Пермского края и Республики Удмуртия представляет собой единую популяцию. Генетическая дифференциация телят, рожденных в Свердловской обл. в период 2008–2019 гг., и быков голштинской породы, содержащихся в АО “Уралплемцентр”, статистически значимо не отличается от ноля, что позволяет утверждать о принадлежности современной популяции крупного рогатого скота региона к голштинской породе. Показано статистически значимое снижение параметров генетического разнообразия популяции молочного скота Свердловской обл. в период 2010–2019 гг., что свидетельствует о росте инбридинга в популяции.
Голштинизированный черно-пестрый скот является наиболее массовой породой крупного рогатого скота молочного направления на территории Урала. Численность животных в регионе составляет около 500 тыс. голов [1]. История формирования данной популяции включает несколько этапов миграции генов, а также несколько центров селекции. При формировании местного тагильского скота использовались племенные производители голландской породы, затем в 1932–1936 гг. начались работы по скрещиванию тагильского скота с остфризскими быками-производителями с целью создания уральского черно-пестрого скота, формирование которого завершилось к 1959 г. Последующее совершенствование уральского черно-пестрого скота с 1965 по 1985 гг. осуществлялось путем прилития крови голландского скота, а с 1983 г., когда была принята программа создания нового внутрипородного типа, путем межпородного скрещивания с голштинскими быками-производителями [2]. Сходная ситуация имела место в Челябинской, Пермской и Курганской обл. В то же время в Тюменской и Курганской обл. имеется значительное количество черно-пестрого скота, родственно не связанного с тагило-остфризским. Таким образом, при формировании современного молочного скота Урала произошло смешение как минимум четырех пород: тагильской, голландской, черно-пестрой и голштинской [1, 3].
Современным методическим стандартом изучения структуры популяций крупного рогатого скота является использование SNP-чипов средней и высокой плотности [4–8]. Однако относительно высокая цена тестов ограничивает размер выборок животных, которые могут быть протипированы в рамках научных проектов. Исторически SNP-чипы пришли на смену микросателлитным маркерам, которые успешно применялись для исследования популяционной структуры крупного рогатого скота до конца нулевых годов XXI в. [9–11]. К настоящему времени в рамках работ по определению достоверности происхождения животных на территории Урала нами типировано несколько тысяч особей голштинизированного черно-пестрого скота по 15 микросателлитным локусам. С учетом большого размера полученной выборки и большого охвата стад животных региона не вызывает сомнения, что полученная база данных является хорошей основой для анализа пространственной структуры популяции и изменения этой структуры во времени. Целью данной работы является описание генетической структуры голштинизированного черно-пестрого скота Урала и оценка динамики генофонда популяции. Наибольшее внимание будет уделено популяции Свердловской обл.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для анализа послужили данные об изменчивости 15 микросателлитных локусов. Всего было проанализировано 17 936 особей крупного рогатого скота. После исключения из базы совпадающих генотипов (монозиготных близнецов) и животных с неидентифицированными аллелями (пропусками данных) выборка составила 15 786 образцов. Из них:
1. Голштинизированный черно-пестрый скот, содержащийся в племзаводах и племрепродукторах Свердловской обл. (21 организация из 46 существующих в области) – 13 224 особи.
2. Быки-производители голштинской породы, содержащиеся в АО “Уралплемцентр” (выборка Hol1) – 48 особей.
3. Животные голштинской породы, эмбрионы которых были привезены на Урал из Канады (выборка Hol2) – 6 особей.
4. Голштинизированный черно-пестрый скот, принадлежащий племзаводам и племрепродукторам Курганской, Тюменской, Челябинской обл., Пермского края и Республики Удмуртия (всего 8 организаций) – 1569 особей.
5. Животные тагильской породы Tag – 92 особи. Выборка получена из последнего стада данной породы, Пермский край.
6. В качестве внешнего контроля использовали две выборки животных герефордской породы (Her), Пермский край, 847 особей.
ДНК животных выделяли из крови, законсервированной в цитрате натрия или ЭДТА, с использованием набора “ДНК-Экстран-1” (“Синтол”, Россия). В анализ были включены 15 микросателлитных локусов, локализованных в различных хромосомах: BM1824, INRA23, BM2113, SPS115, ETH10, TGLA122, ETH225, TGLA126, TGLA227, BM1818, ETH3, TGLA53, CSRM60, CSSM66, ILSTS006. ПЦР проводили с использованием набора COrDIS Cattle (“Гордиз”, Россия) согласно протоколу производителя. Длины амплифицированных фрагментов определяли на генетическом анализаторе ABI 3130 (Applied Biosystems, США) в присутствии маркера молекулярной массы S-550 (“Гордиз”). Хроматограммы расшифровывали в программе Gene Mapper v. 4.0.
Для анализа популяционной структуры использовали метод главных координат (PCoA) и байесовскую кластеризацию. PCoA проводили в пакете ade4 v. 1.7-13 [12] R v. 3.5.2 software [13]. При расчете брали классические дистанции Эвклида (или дистанции Роджерса), полученные в пакете adegenet v. 2.1.2 [14]. Данные о годе рождения животных брали из базы Селэкс “Молочный скот”. Байесовскую кластеризацию выполнили в программе STRUCTURE v. 2.3.4 [15]. Анализ позволил выделить два генофонда (K = 2), условно названных “A” и “B”, объединение которых привело к формированию современного генофонда каждого проанализированного стада.
Генетическую дифференциацию выборок (FST) рассчитывали с использованием пакета hierfstat v. 0.04-22 по формуле M. Nei [16]. Для тестирования гипотезы об отличии параметра FST от ноля использовали программу GenAlEx v. 6.1 [17], при расчетах проводили 999 реплик.
Среднее (Na) и эффективное (Ne) число аллелей на локус, а также индекс фиксации (F) рассчитывали в программе GenAlEx v. 6.1 [17]. Аллельное разнообразие (Ar15, а именно число аллелей, которое будет обнаружено в выборке из 15 особей), ожидаемую (He) и наблюдаемую (Ho) гетерозиготности оценивали с использованием пакета hierfstat v. 0.04-22. При анализе связи двух параметров использовали линейную регрессионную модель типа:
Всего построили три модели. В качестве объясняемой переменной (y) брали параметры Ar15, He, Ho. В качестве объясняющей переменной (x), год рождения особей. Связь считали установленной при статистически значимом отличии коэффициента b от ноля (p < 0.05).
РЕЗУЛЬТАТЫ
Индексы, характеризующие генетическую изменчивость голштинизированного черно-пестрого скота Свердловской обл., приведены в табл. 1. В 15 локусах обнаружено 164 аллеля, частота встречаемости 38 из них менее 0.1%. Среднее число аллелей на локус равняется 10.9, эффективное число аллелей на локус – 4.02. Параметры наблюдаемой и ожидаемой гетерозиготностей близки, они составляют 0.73 (Ho) и 0.72 (He).
Таблица 1.
Параметры генетической изменчивости 15 микросателлитных локусов голштинизированной черно-пестрой породы крупного рогатого скота Свердловской обл. (N = 13 224)
| Локус | Ch | Размеры фрагментов (п.н.)* | Na | Ne | Ho | He | F | Редкие аллели** |
|---|---|---|---|---|---|---|---|---|
| BM1824 | 1 | 174, 178–190 | 7 | 3.72 | 0.73 | 0.73 | –0.005 | 174, 184 |
| BM2113 | 2 | 125–143, 147 | 11 | 4.20 | 0.76 | 0.76 | –0.002 | 129, 131, 141, 143, 147 |
| ETH3 | 19 | 103, 109, 113, 117–131, 139 | 12 | 3.40 | 0.71 | 0.71 | –0.005 | 103, 109, 113, 139 |
| ETH10 | 5 | 209, 213–225 | 8 | 4.34 | 0.78 | 0.77 | –0.011 | |
| ETH225 | 9 | 140–152 | 7 | 3.38 | 0.70 | 0.70 | 0.008 | |
| INRA23 | 3 | 198–216 | 10 | 4.15 | 0.76 | 0.76 | –0.007 | |
| SPS115 | 15 | 248–261 | 8 | 2.27 | 0.56 | 0.56 | –0.010 | 250, 261 |
| TGLA53 | 16 | 154, 158–190 | 18 | 7.17 | 0.87 | 0.86 | –0.007 | 172, 180, 188, 190 |
| TGLA122 | 21 | 139–153, 157–165, 169–177, 181–185 | 21 | 6.60 | 0.84 | 0.85 | 0.014 | 157, 159, 165, 169, 175, 177, 185 |
| TGLA126 | 20 | 111–123 | 7 | 2.60 | 0.62 | 0.62 | –0.009 | 111, 113 |
| TGLA227 | 18 | 77–103 | 14 | 6.10 | 0.85 | 0.84 | –0.011 | 85, 95 |
| BM1818 | 23 | 258–272 | 8 | 2.59 | 0.62 | 0.61 | –0.010 | 272 |
| CSRM60 | 10 | 86–106 | 11 | 2.86 | 0.66 | 0.65 | –0.009 | 86, 88, 90, 94 |
| CSSM66 | 14 | 179–199, 207 | 12 | 4.05 | 0.75 | 0.75 | 0.006 | 199 |
| ILSTS006 | 7 | 282–300 | 10 | 3.08 | 0.67 | 0.68 | 0.003 | 282, 284, 298, 300 |
| Среднее | 10.9 | 4.03 | 0.73 | 0.72 | –0.004 | |||
| SE | 1.1 | 0.39 | 0.02 | 0.02 | 0.002 |
Примечаниe. Ch – хромосома, в которой расположен локус, Na – среднее число аллелей на локус, Ne – эффективное число аллелей на локус, Ho – наблюдаемая гетерозиготность, He – ожидаемая гетерозиготность, F – индекс фиксации, * – все локусы представляют собой динуклеотидные повторы, ** – аллели с частотой встречаемости менее 0.1%.
Для сравнения уровня генетического разнообразия популяции голштинизированного черно-пестрого скота Свердловской обл. с другими популяциями голштинской и черно-пестрой пород использовали параметр ожидаемой гетерозиготности (He). Общими микросателлитными маркерами, используемыми в большинстве проанализированных литературных источников, были девять локусов: BM1824, INRA23, BM2113, SPS115, ETH10, TGLA122, ETH225, TGLA126, TGLA227, для которых нами были рассчитаны средние значения параметра He. В выборках голштинизированного скота Китая значения He составили 0.66 [18] и 0.73 [19], в популяции Чехии – 0.75 [20], Ирана – 0.76 [21], Турции – 0.77 [22]. Для черно-пестрой породы, разводимой на территории Республики Беларусь, значение ожидаемой гетерозиготности составляет 0.80 [23]. В популяции голштинизированного черно-пестрого скота Свердловской обл. уровень ожидаемой гетерозиготности для девяти вышеперечисленных локусов равняется 0.73, что меньше большинства значений, отмеченных в других локальных популяциях животных.
Кластеризация всех имеющихся выборок в осях главных координат представлена на рис. 1. Одной выборкой (или стадом) считали животных, принадлежащих одной организации. На рис. 1,а в качестве внешнего контроля для скота молочного направления использовали герефордскую (мясную) породу животных. Первая ось разделила три чистопородные группы (голштинскую, тагильскую и герефордскую). Ожидаемо, что породы молочного направления оказались более близки между собой, уровень генетической дифференциации между выборками Hol1 и Tag составляет FST = 0.03 (p < 0.01), выборками Tag и Her FST = 0.08 (p < 0.01), Hol1 и Her FST = 0.12 (p < 0.01). Голштинизированный черно-пестрый скот уральского региона по первой оси главных координат расположился между голштинской и тагильской породами.
Рис. 1.
Положение выборок крупного рогатого скота по осям главных координат. Кругами показаны животные, содержащиеся на территории Свердловской обл., треугольниками – животные, содержащиеся на территории Курганской, Тюменской обл., Пермского края и Республики Удмуртия. Размер фигуры пропорционален десятичному логарифму размера выборки. Обозначения: Hol1 – быки голштинской породы, содержащиеся в АО “Уралплемцентр”, Hol2 – особи голштинской породы, родиной которых является Канада, Tag – выборка тагильской породы, Her – выборки герефордской породы. Необозначенные выборки представляют голштинизированный черно-пестрый скот. а – ординация мясного и молочного скота (включает выборки герефордской породы), б – ординация выборок молочного скота.
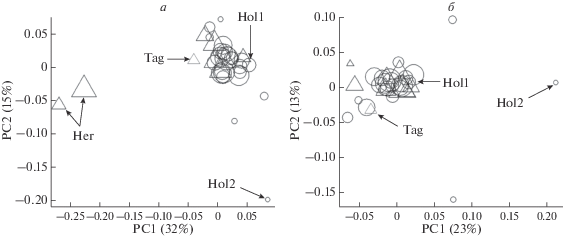
Для более детального анализа целевой группы животных отдельно была проведена кластеризация скота молочного направления, рис. 1,б. По результатам анализа выборки животных можно разделить на три группы. К первой относятся 27 из 29 стад голштинизированного черно-пестрого скота, содержащиеся на территории различных областей региона, быки, принадлежащие АО “Уралплемцентр”, а также выборка тагильской породы. Значения генетической дифференциации в пределах группы составляет FST = 0.004–0.037 (в среднем FST = 0.017), рис. 1,б. Вторая группа представлена животными из двух хозяйств, расположенных на территории Свердловской обл., данные выборки удалены от основной группы животных как по первой оси главных координат, так и по второй. Средний уровень дифференциации животных первой и второй групп составляет FST = 0.04. Вероятной причиной подобных различий является то, что поставщиком спермопродукции в хозяйства второй группы является не АО “Уралплемцентр”, а другие организации. Уровень генетической дифференциации животных второй группы и выборки быков, принадлежащих АО “Уралплемцентр”, статистически значимо отличается от ноля (p < 0.006). Наиболее дифференцированной группой молочного скота является выборка Hol2, т. е. чистокровные голштины, родиной которых является Канада. Уровень генетической дифференциации выборки Hol2 от других стад молочного скота составляет FST = 0.04–0.09 (в среднем FST = 0.06), рис. 1. Генетическая дифференциация выборок Hol1 и Hol2 статистически значимо отличается от ноля (FST = 0.04, p = 0.009) и превышает дифференциацию пары выборок Hol1 и Tag (FST = 0.03, p < 0.01).
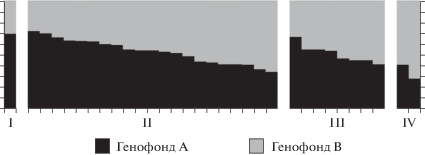
Результаты байесовской кластеризации стад представлены на рис. 2. При K = 2 проанализированные стада различаются по соотношению вклада двух генофондов, условно названных “А” и “В”. Доля генофонда “В” в стадах варьирует от 0.28 до 0.74. Максимальное значение показателя отмечено в выборке голштинской породы (Hol2), а в выборке животных тагильской породы наблюдается одно из самых низких значений данного показателя (доля генофонда “В” составляет 0.31), рис. 2.
Рис. 2.
Результаты байесовской кластеризации крупного рогатого скота (K = 2), учетная единица стадо. Колонки показывают доли генофондов “A” и “B” в следующих группах: I – стадо тагильской породы, II – стада, принадлежащие племзаводам и племрепродукторам Свердловской обл., III – стада, принадлежащие племзаводам и племрепродукторам Курганской, Тюменской, Челябинской обл., Пермского края и Республики Удмуртия, IV – группа быков голштинской породы, содержащихся в АО “Уралплемцентр” (выборка Hol1) и группа животных голштинской породы, привезенная на Урал из Канады (выборка Hol2).
Ординация выборок голштинизированного черно-пестрого скота, содержащегося в племзаводах и племрепродукторах Свердловской обл., в зависимости от года рождения животного, приведена на рис. 3. Полученные данные указывают на постепенное изменение генофонда популяции Свердловской обл. в период 2008–2019 гг. в сторону увеличения сходства с быками голштинской породы, принадлежащими АО “Уралплемцентр”, и снижения сходства с выборкой тагильской породы. Значения параметров генетической дифференциации между выборками голштинизированного черно-пестрого скота, рожденного в различные годы, и голштинами (Hol1) варьируют от FST = 0.002 до 0.029 (табл. 2), однако статистически значимо не отличаются от ноля (p < 0.05).
Рис. 3.
Ординация выборок голштинизированного черно-пестрого скота, содержащегося в племзаводах и племрепродукторах Свердловской обл., в зависимости от года рождения животного. Цифрами 2008–2019 показаны выборки животных разных годов рождения (размеры выборок приведены в табл. 2), Hol1 – выборка быков голштинской породы, содержащихся в АО “Уралплемцентр”, Tag – животные тагильской породы.
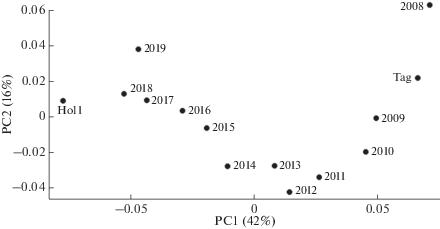
Таблица 2.
Показатели генетического разнообразия и генетической дифференциации выборок голштинизированного черно-пестрого скота, содержащегося в племзаводах и в племенных репродукторах Свердловской обл., в зависимости от года рождения животных
| Год | N | Ho | He | Ar15 | FST : Hol1 – год |
|---|---|---|---|---|---|
| 2008 | 18 | 0.71 | 0.73 | 90.7 | 0.029 |
| 2009 | 60 | 0.71 | 0.74 | 89.9 | 0.028 |
| 2010 | 120 | 0.75 | 0.74 | 93.3 | 0.023 |
| 2011 | 273 | 0.75 | 0.74 | 91.9 | 0.020 |
| 2012 | 486 | 0.75 | 0.74 | 90.7 | 0.020 |
| 2013 | 812 | 0.74 | 0.73 | 88.4 | 0.015 |
| 2014 | 1053 | 0.73 | 0.72 | 88.3 | 0.013 |
| 2015 | 1963 | 0.73 | 0.72 | 86.1 | 0.006 |
| 2016 | 2852 | 0.72 | 0.72 | 84.7 | 0.003 |
| 2017 | 3094 | 0.72 | 0.71 | 82.9 | 0.002 |
| 2018 | 1152 | 0.71 | 0.71 | 82.4 | 0.002 |
| 2019 | 916 | 0.72 | 0.71 | 84.2 | 0.006 |
Примечаниe. N – размер выборки, Ho – наблюдаемая гетерозиготность, He – ожидаемая гетерозиготность, Ar15 – разнообразие аллелей (ожидаемое число аллелей 15 локусов, которое будет обнаружено в выборке размером 15 особей). Генетическая дифференциация рассчитана между выборкой Hol1 и выборками уральского скота разных лет рождения.
Динамика показателей генетического разнообразия в выборках животных, рожденных в период 2008–2019 гг. приведена в табл. 2. Можно выявить два тренда. Во-первых, закономерное, статистически значимое снижение показателей в период 2010–2019 гг.: Ho с 0.75 до 0.72 (p(b) $ \ll $ 0.01), He с 0.74 до 0.71 (p(b) $ \ll $ 0.01), Ar15 с 93.3 до 84.2 (p(b) $ \ll $ 0.01). Во-вторых, рост показателей, характеризующих генетическое разнообразие популяции в период 2008–2010 гг.
ОБСУЖДЕНИЕ
Генетическая дифференциация чистопородного скота
В исследовании проанализированы четыре выборки чистопородных животных, представляющих голштинскую (две выборки), тагильскую и герефордскую породы. Как и ожидалось, наиболее отличной от остальных групп животных оказалась группа животных мясного направления продуктивности (герефордская). Уровень генетической дифференциации между ней и остальными выборками превышает FST > 0.08. Генетическая дифференциация двух выборок животных голштинской породы (FST = 0.04) была выше величины дифференциации быков голштинской породы, принадлежащих АО “Уралплемцентр” и тагильской породы (FST = 0.03). Полученные данные свидетельствуют о высоком генетическом разнообразии в пределах голштинской породы животных, а также высоком генетическом сходстве локальной уральской породы (тагильской) и быков, спермопродукцию которых используют в Свердловской обл. в настоящее время.
Объяснить наблюдаемый уровень дифференциации трех выборок молочных пород можно с использованием данных о регионе происхождения животных. При формировании тагильского скота шла активная миграция генов из популяции Северной Европы, а именно холмогорской и голландской пород, генетически близких к европейской субпопуляции голштинской породы [4, 5]. Значительная часть выборки быков, принадлежащих АО “Уралплемцентр”, также происходит из региона Северной Европы. На контрасте с ними выборка голштинской породы (Hol2) представляет субпопуляцию Северной Америки.
Пространственная структура популяции голштинизированного черно-пестрого скота Урала
Результаты анализа, проведенного с использованием метода главных координат и байесовской кластеризации, свидетельствует о том, что административные границы Уральского региона не являются фактором, влияющим на генетическую дифференциацию стад голштинизированного черно-пестрого скота. Животные, содержащиеся в племзаводах и племрепродукторах Свердловской, Курганской, Тюменской, Челябинской областей, Пермского края и Республики Удмуртия, представляют собой единую популяцию с незначительным уровнем генетической дифференциации. В настоящее время на территории Урала присутствуют, во-первых, стада голштинизированного черно-пестрого скота, генетически сходные со стадом тагильской породы, во-вторых, стада, генетически сходные с группой быков голштинской породы, принадлежащих АО “Уралплемцентр”, в-третьих, стада с “промежуточным” генофондом относительно тагильской и голштинской пород. Стада племенных животных, генетически сходные с североамериканской субпопуляцией голштинской породы, в регионе не обнаружены.
Динамика генофонда популяции голштинизированного черно-пестрого скота Свердловской области в период 2008–2019 гг.
В исследовании были проанализированы генетические характеристики выборок телят, рожденных на территории Свердловской обл. в период 2008–2019 гг. Показана постепенная направленная трансформация генофонда популяции голштинизированного черно-пестрого скота в сторону сходства с генофондом быков голштинской породы, содержащихся в АО “Уралплемцентр”. Генетическая дифференциация выборок телят, рожденных в регионе в 2015–2019 гг. и быков голштинской породы была менее FST < 0.01 и статистически значимо не отличалась от ноля, что позволяет считать телят и быков АО “Уралплемцентр” принадлежащими к одной породе, а именно голштинской. В период 2010–2019 гг. трансформация генофонда уральской популяции сопровождалась снижением параметров, характеризующих ее генетическое разнообразие, что свидетельствует о росте инбридинга в популяции.
Неожиданностью было то, что животные, рожденные в данном регионе в 2008–2009 гг., были генетически сходны со скотом тагильской породы, хотя миграция генов между этими группами прекратилась в 30-е годы прошлого века. Очевидно, что это результат использования для улучшения уральского черно-пестрого скота в период 1958–1983 гг. голландских быков-производителей из Северной Европы, которые были генетически слабо дифференцированны от животных тагильской породы. После 2010 г. заметное влияние на генофонд уральской популяции стали оказывать быки, происходящие из других субпопуляций голштинской породы, в частности, североамериканской. Более подробно этот вопрос будет изучен в последующих исследованиях.
ЗАКЛЮЧЕНИЕ
Современные стада голштинизированного черно-пестрого скота, содержащегося в племзаводах и племрепродукторах Свердловской, Курганской, Тюменской, Челябинской обл., Пермского края и Республики Удмуртия, представляют собой единую популяцию. Величина генетической дифференциации телят, рожденных в Свердловской области в 2015–2019 гг. и быков голштинской породы, содержащихся в АО “Уралплемцентр”, составляет менее FST < 0.01 и статистически значимо не отличается от ноля. С учетом того, что с 80-х годов прошлого столетия для улучшения уральского крупного рогатого скота используются исключительно быки голштинской породы и наблюдаемый уровень генетической дифференциации уральского скота и чистопородных голштинских быков минимален, можно актуализировать вопрос о породной принадлежности современных уральских стад племенного молочного скота.
В период 2010–2019 гг. в популяции черно-пестрого скота Свердловской обл. происходит постепенное снижение параметров генетического разнообразия, что свидетельствует о росте инбридинга в популяции. Этот результат показывает принципиальную возможность использования данных, полученных в ходе определения достоверности происхождения крупного рогатого скота с использованием панели из 15 микросателлитных локусов, для мониторинга уровня инбридинга региональной популяции молочного скота.
Отбор и генотипирование образцов выполнены в ФГБНУ УрФАНИЦ УрО РАН в рамках государственного задания “Разработать селекционно-генетические и теоретические основы сохранения и эффективного использования генофонда крупного рогатого скота в Уральском регионе с применением современных биотехнологий”. Обработка результатов исследования выполнена при финансовой поддержке РФФИ и Свердловской обл. в рамках научного проекта № 20-416-660008.
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Гридина С.Л., Гридин В.Ф., Мымрин В.С. и др. Характеристика племенных и продуктивных качеств черно-пестрого скота в областях и республиках Урала. Екатеринбург: УрО РАН, 2018. 80 с.
Мымрин В.С. Черно-пестрый скот на Урале (состояние и методы совершенствования). Екатеринбург: Изд-во УралГАУ, 2003. 145 с.
Гридина С.Л., Мымрин В.С., Гридин В.Ф. и др. Современное состояние и перспективы развития молочного скотоводства на Урале. Екатеринбург: ООО “Веселый пиксель”, 2018. 150 с.
Sermyagin A.A., Dotsev A.V., Gladyr E.A. et al. Whole-genome SNP analysis elucidates the genetic structure of Russian cattle and its relationship with Eurasian taurine breeds // Genet. Sel. Evol. 2018. V. 50. P. 1–13. https://doi.org/10.1186/s12711-018-0408-8
Yurchenko A., Yudin N., Aitnazarov R. et al. Genome-wide genotyping uncovers genetic profiles and history of the Russian cattle breeds // Heredity. 2018. V. 120. № 2. P. 125–137. https://doi.org/10.1038/s41437-017-0024-3
Decker J.E., McKay S.D., Rolf M.M. et al. Worldwide patterns of ancestry, divergence, and admixture in domesticated cattle // PLoS Genet. 2014. V. 10. № 3. https://doi.org/10.1371/journal.pgen.1004254
Iso-Touru T., Tapio M., Vilkki J. et al. Genetic diversity and genomic signatures of selection among cattle breeds from Siberia, eastern and northern Europe // Anim. Genet. 2016. V. 47. № 6. P. 647–657. https://doi.org/10.1111/age.12473
Зиновьева Н.А., Доцев А.В., Сермягин А.А. и др. Изучение генетического разнообразия и популяционной структуры российских пород крупного рогатого скота с использованием полногеномного анализа SNP // С.-хоз. биология. 2016. Т. 51. № 6. С. 788–800. https://doi.org/10.15389/agrobiology.2016.6.788eng
Li M.H., Kantanen J. Genetic structure of Eurasian cattle (Bos taurus) based on microsatellites: Clarification for their breed classification1 // Anim. Genet. 2010. V. 41. № 2. P. 150–158. https://doi.org/10.1111/j.1365-2052.2009.01980.x
Martin-Burriel I., Rodellar C., Canon J. et al. Genetic diversity, structure, and breed relationships in Iberian cattle // J. Anim. Sci. 2011. V. 89. № 4. P. 893–906. https://doi.org/10.2527/jas.2010-3338
Medugorac I., Medugorac A., Russ I. et al. Genetic diversity of European cattle breeds highlights the conservation value of traditional unselected breeds with high effective population size // Mol. Ecol. 2009. V. 18. № 16. P. 3394–3410. https://doi.org/10.1111/j.1365-294X.2009.04286.x
Dray S., Dufour A. The ade4 package: Implementing the duality diagram for ecologists // J. Stat. Soft. 2007. V. 22. № 4. P. 1–20. https://doi.org/10.18637/jss.v022.i04
R Core Team, R: A language and environment for statistical computing. 2019. URL: http://www.R-project.org/.
Jombart T. Adegenet: A R package for the multivariate analysis of genetic markers // Bioinformatics. 2008. V. 24. № 11. P. 1403–1405. https://doi.org/10.1093/bioinformatics/btn129
Pritchard J.K., Stephens M., Donnelly P. Inference of population structure using multilocus genotype data // Genetics. V. 155. № 2. P. 945–959.
Nei M. Molecular Evolutionary Genetics. Columbia: Columbia Univ. Press, 1987. 512 p.
Peakall R., Smouse P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research // Mol. Ecol. Notes. 2006. V. 6. № 1. P. 288–295. https://doi.org/10.1111/j.1471-8286.2005.01155.x
Tian F., Sun D.X., Zhang, Y. Establishment of paternity testing system using microsatellite markers in Chinese Holstein // J. Genet. Genomics. 2007. V. 35. № 5. P. 279–284. https://doi.org/10.1016/s1673-8527(08)60040-5
Zhang Y., Wang Y.C., Sun D.X., Yu Y. Validation of 17 microsatellite markers for parentage verification and identity test in Chinese Holstein cattle // Asian-Australasian J. Anim. Sci. 2010. V. 23. № 4. P. 425–429. https://doi.org/10.5713/ajas.2010.90424
Rehout V., Hradecka E., Citek J. Evaluation of parentage testing in the Czech population of Holstein cattle // Czech J. Anim. Sci. 2006. V. 51. № 12. P. 503–509.
Hashemi M., Amirinia C., Harkinezhad M.T. et al. Validation of pedigree relationships using a multiplex microsatellite marker assay in Iranian Holstein cattle // Ann. Anim. Sci. 2013. V. 13. № 3. P. 481–493. https://doi.org/10.2478/aoas-2013-0042
Ozkan E., Soysal M.I., Ozder M. et al. Evaluation of parentage testing in the Turkish Holstein population based on 12 microsatellite loci // Livestock Sci. 2009. V. 124. № 1–3. P. 101–106. https://doi.org/10.1016/j.livsci.2009.01.004
Глинская Н.А. STR-полиморфизм генофонда крупного рогатого скота Белорусской черно-пестрой породы и его использование в селекции: Дис. … канд. с.-х. наук. Гродно: Гродненский государственный аграрный университет, 2014. 108 с.
Дополнительные материалы отсутствуют.