

Генетика, 2021, T. 57, № 5, стр. 544-556
Структура и пути формирования полиморфизма гордеинов, контролируемых аллелями гордеин-кодирующих локусов в культурном ячмене (Hordeum vulgare L.)
А. А. Поморцев 1, *, А. В. Рубанович 1, Е. В. Лялина 1
1 Институт общей генетики им. Н.И. Вавилова Российской академии наук
119991 Москва, Россия
* E-mail: pomortsev@vigg.ru
Поступила в редакцию 02.07.2020
После доработки 03.08.2020
Принята к публикации 25.08.2020
Аннотация
Методом электрофореза в крахмальном геле изучен полиморфизм гордеина в 2244 образцах ячменя. Выборка включала 1197 стародавних местных сортов из 22 стран, входящих в основные центры разнообразия ячменя и граничащих с ними, 428 сортов ячменя, возделывавшихся на территории бывшего СССР и Российской Федерации с 1929 по 2019 гг. и 619 сортов из стран Европы, Азии, Америки и Африки. По гордеинам, контролируемым аллелями локуса Hrd A, выявлено 156 вариантов блоков компонентов, локуса Hrd B – 271 вариант и локуса Hrd F – 5 вариантов. Основываясь на данных литературы о молекулярных механизмах формирования полиморфизма гордеинов и исследовании искусственных мутантов по гордеин-кодирующим локусам, среди вариантов гордеинов А и В выделены группы фенотипически схожих блоков компонентов – семейства блоков компонентов. Для HRD А выделено 12 семейств (AI–AXII), для HRD B – 17 семейств (BI–BXVII). Семейства существенно различаются по числу составляющих их вариантов блоков: для HRD A от 5 (AXII) до 60 (AI), для HRD B – от 3 (BXVII) до 41 (BXII). Сделано предположение, что семейства, включающие наибольшее число вариантов гордеинов А и В, являются наиболее древними в культурном ячмене. Показана мозаичность и неравномерность частот в распространении семейств вариантов HRD А и HRD B в исследованных местных популяциях из различных стран мира. Предполагается, что появление собственно семейств HRD A и HRD B обусловлено спонтанной гибридизацией между H. vulgare и H. spontaneum при распространении культурного ячменя. Сделан вывод, что полиморфизм гордеинов в культурном ячмене – результат спонтанной гибридизации культурного ячменя с диким предшественником и накоплением мутаций в генах локусов Hrd A и Hrd B у H. vulgare.
Спирторастворимые белки зерна ячменя – гордеины – характеризуются уникальным разнообразием и по своей информативности не уступают AFLP- и RAPD-маркерам [1]. Наиболее полная информация по наследованию и генетическому контролю этих белков получена при использовании метода электрофореза гордеинов в крахмальном геле. Установлено, что электрофоретические компоненты гордеинов наследуются группами (блоками) и контролируются семью сцепленно наследуемыми локусами, локализованными в коротком плече хромосомы 5 (1Н) ячменя, обозначенными как: Hrd A, Hrd B, Hrd C, Hrd D, Hrd E, Hrd F и Hrd G (рис. 1). Локусы Hrd C, Hrd D, Hrd E, Hrd F и Hrd G контролируют отдельные компоненты гордеина и у большинства сортов не обнаружены. Три локуса – Hrd A, Hrd B и Hrd F – являются полиморфными [2–4]. При электрофорезе в полиакриламидном геле с использованием додецилсульфата натрия выделяют четыре группы белковых полос: А, B, C и D. Группы полипептидов В, C и D контролируются локусами, обозначенным Hor 2, Hor 1 и Hor 3 соответственно [5, 6]. Локус Hor 1 соответствует локусу Hrd A, локус Hor 2 – локусу Hrd B [7]. Группа полипептидов А, которые при электрофорезе в крахмальном геле мигрируют в нижний буфер, по своему аминокислотному составу не являются гордеинами [8], а D-гордеины, не выявляемые при электрофорезе в крахмальном геле, и контролируемые локусом Hor 3, относятся к глютелинам [9]. Позднее P.R. Sewry с соавт. [10, 11], используя электрофорез в полиакриламидном геле в присутствии додецилсулфата натрия, подтвердили информацию о существовании локусов Hrd F и Hrd G, обозначив их как Hor 5 и Hor 4 соответственно.
Рис. 1.
Генетический контроль компонентов гордеина, выявляемых при электрофорезе в крахмальном геле.
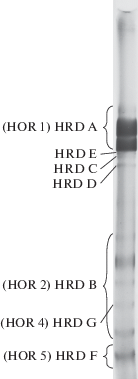
Как видно из рис. 1, блоки компонентов гордеина, контролируемые локусами Hrd A, Hrd B и Hrd F, на электрофореграмме пространственно разделены. Это позволяет как идентифицировать известные варианты блоков компонентов в анализируемых образцах ячменя, так и выделять новые варианты блоков компонентов, контролируемые аллелями указанных локусов при исследовании коллекции. Ранее, в результате электрофоретического анализа гордеинов в крахмальном геле в 619 сортах озимого и ярового ячменя из 27 стран Европы, Азии, Америки и Африки были идентифицированы 29 вариантов блоков компонентов, контролируемых аллелями локуса Hrd A, 34 – локуса Hrd B и 5 – локуса Hrd F [7]. Среди обнаруженных вариантов гордеинов А и В присутствовали группы блоков компонентов, различающиеся между собой по числу, подвижности или интенсивности отдельных белковых полос. Такие группы фенотипически схожих блоков компонентов были названы “семействами блоков компонентов”. Для гордеина А было выделено пять семейств блоков компонентов, для гордеина В – восемь семейств. Варианты гордеина F различались только по электрофоретической подвижности. При этом в отдельных сортах гордеин F отсутствовал (F0). Было высказано предположение, что локусы, контролирующие блоки компонентов, являются сложными и включают несколько генов, а возникновение семейств блоков компонентов гордеинов является следствием различного рода мутаций в генах, входящих в состав сложных локусов.
К настоящему времени информация о полиморфизме годеинов в культурном ячмене существенно увеличилась. Кроме этого, в литературе появились сведения о молекулярной организации гордеин-кодирующих локусов. В настоящей работе приведены новые данные о полиморфизме гордеинов, контролируемых локусами Hrd A и Hrd B, обобщены сведения о молекулярной организации гордеин-кодирующих локусов, позволяющие понять механизмы формирования структуры полиморфизма гордеина.
МАТЕРИАЛЫ И МЕТОДЫ
Разнообразие запасных белков, контролируемых локусами Hrd A, Hrd B и Hrd F в H. vulgare, изучали путем электрофоретического анализа гордеинов в 2244 образцах ячменя. Выборка включала: 1197 стародавних местных сортов из 22 стран, входящих в основные центры разнообразия ячменя и граничащих с ними государств, 228 сортов ярового ячменя, возделывавшихся на территории бывшего СССР с 1929 по 1991 гг., 200 сортов, допущенных к использованию в России с 1992 по 2019 гг. Кроме этого использовали полученные ранее данные по полиморфизму гордеинов в 619 сортах ярового и озимого ячменя из стран Европы, Азии, Америки и Африки [7] (табл. 1). Материал для исследований был получен: из Федерального исследовательского центра Всероссийский институт генетических ресурсов растений им. Н.И. Вавилова (ВИР), National Small Grains Research Collection USDA (Aberdeen, Idaho, США), Research Institute for Bioresources Okayama University (Япония), International Center for Agricultural Research in the Dry Areas – ICARDA (Aleppo, Сирия), кафедры селекции Самаркандского сельскохозяйственного института (Узбекистан), Памирского биологического института (Таджикистан, Хорог), ФГУ “Государственная комиссия Российской Федерации по испытанию и охране селекционных достижений” Министерства сельского хозяйства РФ. Значительная часть образцов местных ячменей была собрана в начале ХХ в. экспедициями Н.И. Вавилова, Е.Г. Черняковской, А.К. Гольбека, В.Е. Писарева и других сотрудников ВИРа, а также экспедициями Д.Р. Харлана (J.R. Harlan). Коллекция местных ячменей из Узбекистана была собрана в 1930-е годы с приусадебных участков декхан селекционерами В.Н. Покровским и Ф.А. Квятковским. Для каждого образца местных ячменей анализировали гордеины в 5–10 линиях, а для селекционных сортов – в 40–100 линиях или индивидуальных зерновках.
Таблица 1.
Число исследованных образцов местных и коммерческих сортов культурного ячменя из различных стан
| № п/п | Страна | Число образцов | № п/п | Страна | Число образцов |
|---|---|---|---|---|---|
| 1 | Марокко (образцы местного ячменя) | 9 | 13 | Иран (то же) | 63 |
| 2 | Алжир (то же) | 22 | 14 | Туркменистан (то же) | 51 |
| 3 | Тунис (то же) | 7 | 15 | Узбекистан (то же) | 202 |
| 4 | Египет (то же) | 39 | 16 | Таджикистан, Горный Бадахшан (то же) | 3 |
| 5 | Эфиопия (то же) | 147 | 17 | Киргизстан (то же) | 47 |
| 6 | Турция (то же) | 93 | 18 | Афганистан (то же) | 84 |
| 7 | Сирия (то же) | 60 | 19 | Пакистан (то же) | 28 |
| 8 | Иордания (то же) | 34 | 20 | Китай (то же) | 96 |
| 9 | Ирак (то же) | 33 | 21 | Непал (то же) | 30 |
| 10 | Палестина (то же) | 8 | 22 | Индия (то же) | 47 |
| 11 | Израиль (то же) | 5 | 23 | Сорта ячменя, районированные в СССР с 1929 по 1991 гг. | 228 |
| 12 | Йемен (то же) | 89 | 24 | Сорта ярового и озимого ячменя из стран Европы, Азии, Америки, Африки | 619 |
Электрофорез гордеинов проводили в столбиках 12–14%-ного крахмального геля с 3 М мочевиной в алюминий-лактатном буфере с рН 3.1 по методике А.А. Созинова и Ф.А. Поперели [12] с некоторыми модификациями [13]. Анализ сходства исследованных местных популяций ячменя проводили методом главных компонент с помощью пакета программ SPSS 19.
РЕЗУЛЬТАТЫ И ОБСУЖДЕИЕ
В результате электрофоретического анализа гордеинов в 2244 образцах ячменя к настоящему времени обнаружено 156 вариантов блоков компонентов, контролируемых аллелями локуса Hrd A (рис. 2), 271 – локуса Hrd B и 5 – локуса Hrd F (рис. 3). Среди выявленного разнообразия гордеинов А мы выделяем группы фенотипически схожих вариантов блоков компонентов – семейства вариантов блоков компонентов. На рис. 4 в качестве примера приведены две такие группы блоков компонентов. Из рис. 4,а видно, что блок компонентов А100 отличается от А2 уменьшением подвижности компонента 4. Блок А30 отличается от А2 и А100 двойным компонентом 4. Вариант А9, в сравнении с А2, характеризуется снижением интенсивности компонента 1, увеличением подвижности компонента 2 и появлением двух минорных компонентов в районе компонента 4. У блока А149, в сравнении с А9, отсутствует компонент 4. Варианты А4 и А58 характеризуются отсутствием компонента 5 или уменьшением его электрофоретической подвижности, соответственно. Аналогичные различия по числу компонентов гордеина А, их подвижности и относительной интенсивности наблюдаются между блоками компонентов на рис. 4,б. Формирование таких семейств можно продемонстрировать на следующем примере. При электрофоретическом анализе гордеинов в ряде мутантных по морфологическим признакам линий, полученных Г.Г. Шведовым (Селекционно-генетический институт, Одесса) путем химического мутагенеза из сорта Одесский 46, нами были обнаружены формы, электрофореграммы гордеина А которых представлены на рис. 5. Из рисунка видно, что на электрофореграмме А-гордеина мутантов М564 и М109 в сравнении с электрофореграммой гордеина исходного сорта, отсутствуют компоненты 6 и 3 соответственно, а у мутанта М88 – изменилась подвижность компонента 5. Таким образом, вариант блока гордеина А3 и его мутантные варианты составляют одно семейство. Каждый из этих вариантов в дальнейшем может служить исходным для возникновения новых вариантов блоков компонентов в результате мутаций.
Рис. 2.
Варианты блоков компонентов гордеина А, обнаруженные при анализе 2244 образцов культурного ячменя.
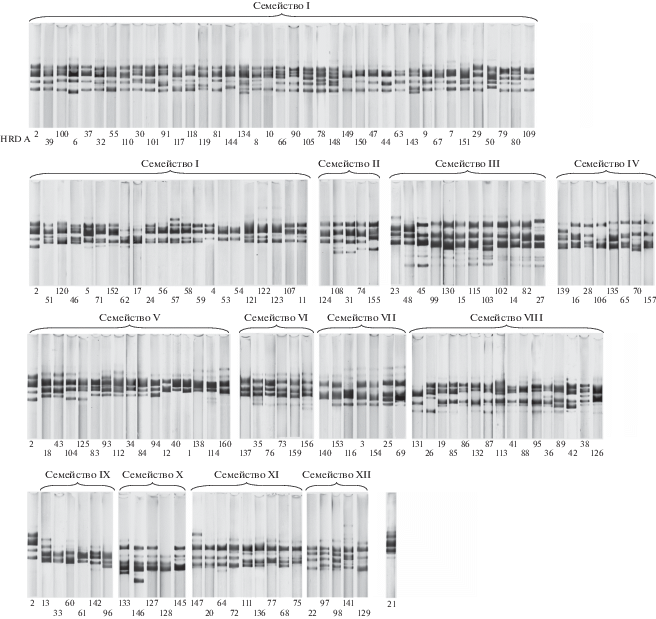
Рис. 3.
Варианты блоков компонентов гордеина В, обнаруженные при анализе 2244 образцов культурного ячменя.

Рис. 4.
Некоторые варианты фенотипически сходных блоков компонентов гордеина А, объединяемые в семейства: а – семейство I, б – семейство III.


Возникновение полиморфизма запасных белков злаков длительное время рассматривалось как результат посттрансляционных изменений, таких как агрегация, дезамидирование, протеолиз и гликозилирование. Однако экспериментальных данных, подтверждающих эти предположения, нет [14]. К настоящему времени известно, что группы полипептидов С- и В-гордеинов кодируются локусами Hor 1 (Hrd A) и Hor 2 (Hrd B), включающими несколько десятков генов [15, 16]. В результате анализа клонированных нуклеотидных последовательностей из различных сортов ячменя было показано, что гены в локусе Hor 1 могут располагаться относительно друг друга по типу “голова к голове” в непосредственной близости друг от друга. Были также обнаружены различия длин кодирующих последовательностей (363–1000 пн), что является молекулярной основой полиморфизма размеров С-гордеинов [17]. Кроме этого степень различий зависит от замен только одной аминокислоты (заряженная/незаряженная), а также от делеций/инсерций отдельных триплетов или блоков. Не исключены значительные делеции кодирующей части генов С-гордеинов [17, 18]. Обработка у ряда сортов и дигаплоидных линий ячменя рестриктазами генов В-гордеинов, амплифицированных с использованием праймеров, гомологичных районам, фланкирующим кодирующие последовательности, выявила полиморфизм, обусловленный приобретением или потерей сайтов рестрикции как внутри, так и между генотипами. При амплификации с использованием праймеров, гомологичных районам внутри кодирующей последовательности и фланкирующих множественные тандемные повторы некоторых генов В-гордеина, была выявлена вариация по длине повторяющегося домена гена В-гордеина как внутри, так и между сортами. Эта изменчивость объясняется изменением числа тандемных повторов в повторяющемся домене в результате неравного кроссинговера и проскальзывания нити ДНК (DNA-slippage) во время репликации [19]. Однако в литературе имеется информация только об одном доказанном рекомбинанте между генами в локусе Hor 2 [19, 20]. Таким образом, наблюдаемый полиморфизм гордеинов ячменя определяется не посттрансляционными изменениями белков, а различиями между генотипами на уровне ДНК.
Основываясь на данных о механизмах формирования полиморфизма годеинов и фенотипическом сходстве электрофоретических спектров, 156 вариантов блоков компонентов, контролируемых аллелями локуса Hrd A, мы объединили в 12 семейств, обозначенных римскими цифрами (AI– AXII). При этом вариант А21 не был отнесен ни к одному из семейств (табл. 2, рис. 2). Из табл. 2 следует, что семейства значительно различаются по числу входящих в них вариантов блоков компонентов. Самое большое семейство – AI – включает 60 (38.5%) вариантов. Сравнительно высокую численность имеют семейства AIII – 12 (7.7%) вариантов, AV и AVIII – по 16 (10.3%) вариантов. Остальные восемь семейств объединяют от 5 (3.2%) до 9 (5.8%) вариантов. Таким образом, четыре семейства из двенадцати объединяют 66.8% известных вариантов блоков компонентов гордеина А.
Таблица 2.
Число вариантов блоков компонентов гордеинов А и В, объединяемых в различные семейства
| Семейства вариантов блоков компонентов гордеина А | |||||
|---|---|---|---|---|---|
| семейство | число вариантов | семейство | число вариантов | семейство | число вариантов |
| AI | *58/60 | AVI | 3/6 | AXI | 8/9 |
| AII | 4/5 | AVII | 6/7 | AXII | 5/5 |
| AIII | 9/12 | AVIII | 15/16 | В21 | 1/1 |
| AIV | 8/8 | AIX | 6/6 | Всего | 142(91%)/156 |
| AV | 14/16 | AX | 5/5 | ||
| Семейства вариантов блоков компонентов гордеина В | |||||
| BI | 8/9 | BVII | 13/16 | BXIII | 35/41 |
| BII | 16/19 | BVIII | 10/12 | BXIV | 7/7 |
| BIII | 14/14 | BIX | 17/19 | BXV | 34/36 |
| BIV | 26/26 | BX | 17/22 | BXVI | 3/3 |
| BV | 5/5 | BXI | 7/7 | BXVII | 3/3 |
| BVI | 8/8 | BXII | 18/24 | Всего | 241(88.9%)/271 |
Среди идентифицированных в H. vulgare 271 вариантов гордеина В мы выделили 17 семейств (BI–BXVII) (рис. 3, табл. 2). Семейства вариантов гордеина В, как и семейства вариантов гордеина А, значительно различаются по числу входящих в них вариантов блоков компонентов. Наибольшее число вариантов объединяют семейства BXIII – 41 (15.1%) и BXV – 36 (13.3%). Сравнительно высокое число вариантов блоков включают так же семейства BIV – 26 (9.6%), BXII – 24 (8.9%), BX – 22 (8.1%), BII и BIX – по 19 (7.0%). Остальные десять семейств включают от 3 (1.1%) до 16 (5.9%) вариантов блоков компонентов гордеина В. Таким образом, семь семейств из семнадцати объединяют 69.0% вариантов HRD В, выделенных в культурном ячмене. Следует отметить, что от общего числа известных электрофоретических вариантов гордеинов А и В в исследованных местных популяциях ячменя обнаружено 91 и 88.9% вариантов блоков компонентов соответственно. Это на наш взгляд объясняется тем, что в основе генофонда современных селекционных сортов лежит генофонд местных популяций ячменя. Однако в современной селекции ячменя с целью поиска новых хозяйственно ценных признаков используется мутагенез. Также в селекционный процесс вовлекается дикий ячмень Hordeum spontaneum C. Koch [21–23]. Здесь мы не рассматриваем распространение пяти вариантов HRD F, так как локус Hrd F, контролирующий эти белки, тесно сцеплен с локусом Hrd B, величина рекомбинации между ними составляет 1.5% рекомбинации [3].
Возникает вопрос о причинах столь значительных различий между семействами гордеинов А и В по числу входящих в них вариантов блоков компонентов, а также как и откуда в культурном ячмене возникли собственно семейства дискретные по фенотипу? В настоящее время установлено, что диким предшественником H. vulgare является H. spontaneum [24]. Однако время и место доместикации ячменя до сих пор не установлены. Так например J.R. Harlan и D. Zohary, опираясь на исследования археологов, выдвинули гипотезу, что ячмень был введен в культуру в Дуге Плодородия в VII тысячелетии до н. э. [25]. При этом наиболее раннее использование ячменя человеком в Юго-Западной Азии, датируемое 17 тысячелетием до н. э., зафиксировано в Охало II (Ohalo II) около Галилейского моря [26]. В то же время зерна культурного ячменя, возраст которых оценивается в 18 000 лет, найдены в археологических раскопках в южном Египте, около Асуана в районе Каббания [27]. Следует отметить, что количество и качество археоботанических доказательств значительно варьируют от района к району. Имеется обширная информация по Балканским странам, Центральной и Западной Европе, но документация по России, Центральной Азии и Индии намного более разрознена. Из Африки только по Египту существуют достаточные данные по растительным остаткам [28]. Таким образом, по имеющимся данным, время доместикации ячменя насчитывает не менее 18 тыс. лет.
Учитывая, что формирование полиморфизма гордеинов во многом обусловлено возникновением различного рода мутаций в генах локусов Hrd A и Hrd B, можно предположить, что число вариантов блоков компонентов, входящих в то или иное семейство, отражает длительность существования такого семейства в культурном ячмене. Таким образом, самым “древним” семейством гордеина А может быть семейство AI, включающее 58 вариантов, обнаруженных в местных популяциях ячменя (табл. 2). При этом представители семейства AI с суммарными частотами от 0.1203 (Узбекистан) до 0.9429 (Тунис) были обнаружены в местных популяциях из 21 страны, кроме Горного Бадахшана (табл. 3). Наибольшее распространение имели три варианта, объединенные в это семейство: в 21-ной стране – А2 с частотами от 0.0719 (Йемен) до 0.4125 (Палестина); в 15 странах – А62 с частотами от 0.001 (Узбекистан) до 0.125 (Палестина), в 14 странах – А24 с частотами от 0.0033 (Иран) до 0.4572 (Эфиопия). Следующим по числу вариантов гордеина А семейством является AVIII – 15 вариантов. Представители этого семейства с суммарными частотами от 0.0020 (Узбекистан) до 0.9218 (Горный Бадахшан) обнаружены в популяциях из 16 стран. Самым распространенным вариантом является вариант А19, присутствовавший в образцах из 15 стран с частотами от 0.0020 в Узбекистане до 0.5527 в Горном Бадахшане. Высокие частоты этого варианта гордеина А обнаружены также в популяциях из Непала (0.2733), Ирака (0.2148), Израиля (0.1800), Китая (0.1417), Пакистана (0.1072). Остальные 14 вариантов присутствовали в местных популяциях ячменя от одной до пяти стран.
Таблица 3.
Распространение семейств блоков компонентов гордеина А в популяциях местных сортов ячменя в различных странах
| Страна | Численности и частоты семейств блоков компонентов гордеина А | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AI | AII | AIII | AIV | AV | AVI | AVII | AVIII | AIX | AX | AXI | AXII | |
| Марокко | *7/0.4444 | 0 | 0 | 0 | 0 | 0 | 2/0.3223 | 0 | 1/0.1222 | 0 | 1/0.0889 | 1/0.0222 |
| Тунис | 2/0.9429 | 0 | 0 | 0 | 0 | 0 | 1/0.0571 | 0 | 0 | 0 | 0 | 0 |
| Алжир | 7/0.4819 | 0 | 1/0.0317 | 2/0.0273 | 1/0.0455 | 0 | 1/0.1455 | 0 | 1/0.1273 | 0 | 3/0.1182 | 1/0.0226 |
| Египет | 9/0.6180 | 1/0.0769 | 1/0.0026 | 2/0.1001 | 1/0.0307 | 2/0.0589 | 0 | 1/0.0077 | 1/0.0256 | 0 | 3/0.0795 | 0 |
| Эфиопия | 17/0.8412 | 1/0.0068 | 1/0.0068 | 0 | 3/0.0221 | 0 | 0 | 1/0.0068 | 3/0.1163 | 0 | 0 | 0 |
| Йемен | 21/0.8498 | 1/0.0517 | 1/0.0048 | 1/0.0112 | 3/0.0033 | 0 | 0 | 2/0.0268 | 2/0.0449 | 2/0.0022 | 1/0.0011 | 2/0.0045 |
| Турция | 26/0.6913 | 2/0.0730 | 0 | 3/0.0398 | 1/0.0022 | 0 | 5/0.1141 | 1/0.0022 | 3/0.0634 | 1/0.0011 | 3/0.0129 | 0 |
| Сирия | 11/0.2899 | 0 | 2/0.0447 | 0 | 4/0.1549 | 2/0.005 | 3/0.0567 | 1/0.0067 | 3/0.2337 | 1/0.0717 | 4/0.0867 | 3/0.05 |
| Иордания | 9/0.1881 | 1/0.0088 | 2/0.0442 | 4/0.2648 | 3/0.1087 | 1/0.0059 | 1/0.0266 | 0 | 2/0.2854 | 3/0.0293 | 3/0.0235 | 3/0.0147 |
| Израиль | 2/0.2400 | 0 | 0 | 2/0.08 | 1/0.020 | 1/0.2000 | 1/0.2800 | 1/0.1800 | 0 | 0 | 0 | 0 |
| Палестина | 4/0.6625 | 0 | 0 | 2/0.125 | 0 | 0 | 1/0.1500 | 0 | 1/0.0250 | 0 | 1/0.0375 | 0 |
| Ирак | 2/0.2559 | 1/0.0294 | 2/0.2352 | 1/0.0294 | 1/0.0059 | 0 | 1/0.0589 | 4/0.3265 | 0 | 1/0.0294 | 1/0.0294 | 0 |
| Иран | 14/0.4796 | 1/0.1476 | 3/0.0142 | 2/0.0142 | 5/0.1524 | 0 | 1/0.0968 | 4/0.0412 | 2/0.0317 | 0 | 2/0.0064 | 1/0.0159 |
| Туркмения | 10/0.3235 | 0 | 5/0.2606 | 0 | 6/0.2746 | 0 | 1/0.0784 | 2/0.0491 | 2/0.0138 | 0 | 0 | 0 |
| Узбекистан | 10/0.1203 | 1/0.0020 | 3/0.0207 | 1/0.0296 | 5/0.7742 | 0 | 2/0.0404 | 1/0.0020 | 1/0.0049 | 0 | 2/0.0059 | 0 |
| Таджикистан | 0 | 0 | 1/0.0043 | 0 | 1/0.0739 | 0 | 0 | 5/0.9218 | 0 | 0 | 0 | 0 |
| Киргизия | 9/0.5702 | 0 | 1/0.0255 | 1/0.0213 | 4/0.2936 | 0 | 1/0.0511 | 0 | 0 | 0 | 1/0.0383 | 0 |
| Афганистан | 14/0.2535 | 1/0.0012 | 3/0.0929 | 0 | 8/0.4213 | 0 | 1/0.0667 | 5/0.1048 | 4/0.0536 | 0 | 1/0.0024 | 2/0.0036 |
| Пакистан | 3/0.1805 | 0 | 2/0.107 | 1/0.0081 | 4/0.5472 | 0 | 0 | 2/0.1429 | 1/0.0143 | 0 | 0 | 0 |
| Индия | 10/0.3873 | 0 | 3/0.1575 | 0 | 5/0.0979 | 0 | 1/0.0467 | 1/0.0776 | 2/0.0255 | 0 | 1/0.0213 | 1/0.1862 |
| Китай | 11/0.4062 | 2/0.0146 | 3/0.0667 | 2/0.0063 | 5/0.2104 | 0 | 1/0.0187 | 8/0.2646 | 0 | 0 | 0 | 1/0.0125 |
| Непал | 3/0.5267 | 0 | 1/0.0333 | 0 | 3/0.1400 | 0 | 0 | 3/0.3000 | 0 | 0 | 0 | 0 |
| Число стран | 21 | 10 | 17 | 13 | 19 | 4 | 16 | 16 | 15 | 5 | 14 | 9 |
* Здесь и в табл. 4 в числителе – число вариантов блоков компонентов, представляющих семейство, в знаменателе – суммарная частота вариантов блоков компонентов, входящих в семейство.
Семейство AV включает 14 вариантов блоков компонентов с суммарными частотами от 0.0022 (Турция) до 0.7742 (Узбекистан), обнаруженных в местных популяциях ячменя из 19 стран кроме Марокко, Туниса и Палестины (табл. 2, 3). Среди вариантов гордеина А, вошедших в это семейство, самое широкое распространение наблюдается для HRD A1 – в 11 странах с частотами от 0.0017 (Киргизия) до 0.1400 (Сирия), HRD A18 – в 11 странах с частотами от 0.0011 (Йемен) до 0.6441 (Узбекистан) и HRD A12 – в 10 странах с частотами от 0.0033 (Сирия) до 0.1362 (Киргизия). Одиннадцать вариантов блоков компонентов гордеина А из этого семейства были обнаружены в популяциях ячменя от одной до восьми стран.
В семейство AIII входят девять вариантов гордеина, идентифицированных в местных популяциях из 17 стран, с суммарными частотами от 0.0026 (Египет) до 0.2606 (Туркмения). При этом вариант HRD A14 был обнаружен в образцах ячменя из 15 стран с частотами от 0.0014 в Сирии до 0.1176 в Ираке. Остальные варианты присутствовали в популяциях ячменя из отдельных стран, за исключением HRD A15, найденного в образцах из шести стран с частотами 0.0026 (Египет) – 0.0357 (Пакистан).
Среди 17 семейств гордеина В наибольшее число вариантов блоков компонентов включают семейства BXIII – 35 вариантов, BXV – 34 варианта и BIV – 26 вариантов. В остальные 14 семейств входят от 3 до 18 вариантов (табл. 2). Представители семейства BXIII выявлены в популяциях ячменя из 21 страны, кроме Горного Бадахшана, с суммарными частотами от 0.0200 (Непал) до 0.4000 (Израиль) (табл. 4). Наибольшее распространение имел HRD B17, обнаруженный в образцах из 14 стран с частотами от 0.0011 (Йемен) до 0.0383 (Сирия). Вариант HRD B11 присутствовал в популяциях из 11 стран с частотами от 0.0049 (Узбекистан) до 0.2583 (Китай). Каждый из вариантов HRD B12 и HRD B82 с частотами от 0.0011 (Узбекистан) до 0.2507 (Йемен) и от 0.0010 (Узбекистан) до 0.1000 (Палестина) соответственно были обнаружены в образцах из 10 стран. Тридцать один вариант блоков компонентов горедина В, входящие в это семейство, обнаружены в образцах от одной до девяти стран.
Таблица 4.
Распространение семейств блоков компонентов гордеина В в популяциях местных сортов ячменя в различных странах
| Страна | Численности и частоты семейств блоков компонентов гордеина B | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| BI | BII | BIII | BIV | BV | BVI | BVII | BVIII | BIX | |
| Марокко | 0 | 0 | 0 | 1/0.2334 | 0 | 1/0.0222 | 2/0.3779 | 0 | 1/0.0444 |
| Тунис | 0 | 0 | 0 | 1/0.1429 | 0 | 0 | 1/0.4858 | 0 | 1/0.0444 |
| Алжир | 0 | 0 | 0 | 2/0.0728 | 0 | 1/0.0091 | 1/0.1727 | 0 | 3/0.1365 |
| Египет | 1/0.0256 | 0 | 1/0.0256 | 5/0.2744 | 0 | 0 | 1/0.1385 | 0 | 0 |
| Эфиопия | 4/0.0341 | 1/0.0068 | 0 | 4/0.0227 | 0 | 1/0.0125 | 1/0.0057 | 3/0.3511 | 3/0.0381 |
| Йемен | 0 | 0 | 3/0.0146 | 4/0.0338 | 1/0.0067 | 1/0.0022 | 2/0.0157 | 3/0.0123 | 5/0.1023 |
| Турция | 2/0.0430 | 1/0.0032 | 1/0.0011 | 3/0.0559 | 2/0.0990 | 1/0.0516 | 5/0.0420 | 3/0.0183 | 3/0.0914 |
| Сирия | 2/0.0117 | 2/0.005 | 3/0.0116 | 1/0.4686 | 2/0.0083 | 2/0.0150 | 1/0.0317 | 0 | 3/0.0133 |
| Иордания | 1/0.0088 | 2/0.0088 | 1/0.0765 | 9/0.2735 | 1/0.0029 | 1/0.0029 | 2/0.0176 | 1/0.0176 | 3/0.0707 |
| Израиль | 1/0.0400 | 0 | 0 | 0 | 1/0.0800 | 0 | 1/0.2400 | 0 | 1/0.0400 |
| Палестина | 0 | 0 | 0 | 1/0.175 | 1/0.0250 | 1/0.0250 | 2/0.150 | 0 | 2/0.1375 |
| Ирак | 1/0.0088 | 0 | 0 | 4/0.1118 | 0 | 0 | 1/0.0059 | 1/0.0118 | 4/0.4660 |
| Иран | 3/0.0222 | 4/0.0461 | 1/0.0016 | 5/0.0651 | 2/0.1016 | 4/0.0651 | 2/0.0825 | 2/0.0159 | 5/0.1602 |
| Туркмения | 2/0.0079 | 3/0.1117 | 3/0.0333 | 3/0.0824 | 0 | 1/0.0020 | 2/0.0236 | 1/0.0588 | 4/0.3901 |
| Узбекистан | 3/0.0060 | 5/0.4135 | 2/0.0059 | 4/0.0390 | 1/0.0413 | 4/0.0139 | 5/0.0109 | 1/0.0010 | 1/0.0030 |
| Таджикистан | 0 | 0 | 0 | 2/0.3708 | 0 | 0 | 0 | 0 | 1/0.0241 |
| Киргизия | 2/0.2808 | 2/0.1277 | 0 | 3/0.0638 | 1/0.0170 | 1/0.0213 | 1/0.0170 | 1/0.0426 | 3/0.2297 |
| Афганистан | 4/0.0274 | 1/0.1429 | 4/0.0476 | 8/0.0785 | 1/0.0274 | 1/0.0036 | 3/0.0096 | 4/0.0464 | 5/0.1572 |
| Пакистан | 0 | 3/0.2858 | 0 | 3/0.1428 | 0 | 0 | 1/0.0143 | 0 | 2/0.0500 |
| Индия | 2/0.0852 | 4/0.2638 | 1/0.0128 | 7/0.1745 | 1/0.0213 | 1/0.0128 | 2/0.0084 | 4/0.0425 | 0 |
| Китай | 3/0.0667 | 2/0.0208 | 2/0.0250 | 5/0.2209 | 2/0.0167 | 1/0.0042 | 1/0.0021 | 1/0.0104 | 3/0.0416 |
| Непал | 0 | 0 | 0 | 2/0.2200 | 0 | 0 | 0 | 0 | 0 |
| Число стран | 14 | 12 | 11 | 21 | 12 | 15 | 20 | 12 | 19 |
| Страна | Численности и частоты семейств блоков компонентов гордеина B | ||||||||
| BX | BXI | BXII | BXIII | BXIV | BXV | BXVI | BXVII | ||
| Марокко | 0 | 2/0.2333 | 0 | 2/0.0444 | 1/0.0444 | 0 | 0 | 0 | |
| Тунис | 1/0.0285 | 2/0.2857 | 0 | 1/0.0571 | 0 | 0 | 0 | 0 | |
| Алжир | 1/0.0455 | 4/0.1272 | 3/0.1454 | 5/0.2046 | 0 | 2/0.0862 | 0 | 0 | |
| Египет | 1/0.0128 | 2/0.2026 | 5/0.1462 | 3/0.1076 | 1/0.0051 | 2/0.0360 | 1/0.0256 | 0 | |
| Эфиопия | 1/0.0023 | 0 | 2/0.0250 | 7/0.0727 | 0 | 9/0.4290 | 0 | 0 | |
| Йемен | 0 | 0 | 0 | 7/0.3360 | 0 | 19/0.4349 | 0 | 3/0.0415 | |
| Турция | 7/0.1549 | 3/0.0312 | 6/0.1452 | 10/0.1375 | 2/0.0161 | 3/0.1096 | 0 | 0 | |
| Сирия | 4/0.0200 | 2/0.0850 | 5/0.0416 | 10/0.2649 | 1/0.0050 | 4/0.0133 | 1/0.0017 | 1/0.0033 | |
| Иордания | 2/0.1912 | 1/0.0176 | 6/0.1264 | 11/0.1442 | 0 | 1/0.0413 | 0 | 0 | |
| Израиль | 1/0.1200 | 0 | 0 | 4/0.4800 | 0 | 0 | 0 | 0 | |
| Палестина | 0 | 0 | 2/0.0500 | 5/0.4000 | 0 | 1/0.0375 | 0 | 0 | |
| Ирак | 0 | 0 | 3/0.1634 | 3/0.1765 | 0 | 2/0.0558 | 0 | 0 | |
| Иран | 2/0.0222 | 1/0.0143 | 6/0.1047 | 9/0.1111 | 3/0.0651 | 5/0.1128 | 0 | 1/0.0095 | |
| Туркмения | 3/0.0236 | 0 | 2/0.0274 | 8/0.1294 | 1/0.0176 | 4/0.0902 | 0 | 1/0.0020 | |
| Узбекистан | 3/0.0630 | 1/0.0020 | 4/0.0315 | 7/0.0886 | 2/0.0688 | 6/0.2067 | 1/0.0039 | 1/0.0010 | |
| Таджикистан | 3/0.4134 | 0 | 0 | 0 | 3/0.0951 | 1/0.0923 | 0 | 1/0.0043 | |
| Киргизия | 0 | 1/0.0255 | 2/0.0256 | 5/0.0596 | 0 | 1/0.0894 | 0 | 0 | |
| Афганистан | 4/0.1141 | 2/0.0464 | 4/0.0547 | 12/0.1655 | 2/0.0298 | 6/0.0334 | 0 | 1/0.0155 | |
| Пакистан | 1/0.0714 | 0 | 1/0.0214 | 1/0.0571 | 1/0.0143 | 2/0.1285 | 0 | 1/0.2144 | |
| Индия | 2/0.0426 | 0 | 1/0.0042 | 3/0.0893 | 0 | 5/0.1064 | 1/0.0042 | 1/0.1320 | |
| Китай | 2/0.0271 | 0 | 4/0.0271 | 10/0.3853 | 1/0.0021 | 4/0.1146 | 0 | 1/0.0354 | |
| Непал | 0 | 0 | 0 | 1/0.0200 | 0 | 3/0.2067 | 0 | 1/0.5533 | |
| Число стран | 16 | 11 | 16 | 21 | 11 | 19 | 4 | 11 | |
См. примечание к табл. 3.
Варианты, составляющие семейство BXV, присутствовали в исследованных популяциях из 19 стран, за исключением Марокко, Туниса и Израиля, с суммарными частотами от 0.0133 (Сирия) до 0.4349 (Йемен). Самым распространенным вариантом из этого семейства является HRD В84, идентифицированный в образцах из 12 стран с частотами от 0.0024 (Афганистан) до 0.1456 (Эфиопия). Сравнительно широкое распространение имел HRD B111, найденный в популяциях из семи стран с частотами от 0.0012 (Афганистан) до 0.1456 (Эфиопия). Остальные 32 варианта, составляющие семейство BXV, присутствовали в местных ячменях от одной до пяти стран.
Представители семейства BIV были обнаружены в местных популяциях ячменя из 21 страны, за исключением Израиля, с суммарными частотами от 0.0227 (Эфиопия) до 0.4686 (Сирия). Наибольшее распространение имел вариант HRD B14 – в 15 странах с частотами от 0.0032 (Иран) до 0.2334 (Марокко). В исследованных образцах из десяти стран был обнаружен вариант HRD B2 с частотами от 0.0010 (Узбекистан) до 0.0733 (Сирия). В популяциях из семи стран идентифицированы варианты блоков компонентов HRD B16 и HRD B65. При этом частоты HRD B16 варьировали от 0.0024 (Афганистан) до 0.1146 (Китай), а варианта HRD B65 – от 0.0011 (Турция) до 0.2180 (Сирия). Другие варианты блоков компонентов, входящие в это семейство, присутствовали в исследованных образцах от одной до пяти стран.
Таким образом, наблюдается мозаичность и неравномерность частот в распространении семейств вариантов блоков компонентов HRD А и HRD B в исследованных местных популяциях из различных стран мира. Матрицу суммарных частот семейств одновременно гордеинов A и B в местных популяциях ячменя из 22 стран подвергли кластерному анализу методом главных компонент. Первые две компоненты описывают 38.8% изменчивости частот вариантов семейств А и В (рис. 6). Как видно из рис. 6, по первой главной компоненте исследованные популяции разделились следующим образом: в отрицательную область значений попали образцы ячменя из стран Азии восточнее Ирана, а так же из Ирака, Йемена и Эфиопии. В положительной области значений первой главной компоненты оказались популяции из стран Северной Африки, Ближнего Востока, Передней Азии и Ирана. Таким образом, “восточные” и “западные” местные популяции культурного ячменя различаются как по составу, так и по суммарным частотам семейств блоков компонентов грдеинов А и В. Причиной такой мозаичности в распространении семейств вариантов HRD A и HRD B могут служить, на наш взгляд, несколько факторов: эффект основателя и неоднократная интродукция популяций культурного ячменя в те или иные регионы из различных стран [29]; зависимость распространения аллелей годреин-кодирующих локусов от распределения таких климатических факторов как количество осадков в год, средние температуры июля и континентальность климата [30], высота выращивания популяций над уровнем моря [31].
Рис. 6.
Положение местных популяций ячменя из 22 стран в координатах первой и второй главных компонент изменчивости частот вариантов семейств гордеинов А и В.

Однако каким образом возникли собственно семейства вариантов блоков компонентов? Можно предположить, что возникновение семейств связано с постепенным накоплением мутаций в гордеиновых локусах культурного ячменя в течение длительного периода. Но тогда следовало ожидать присутствие в культурном ячмене неких “переходных” вариантов от одного семейства к другому. Но, как видно из рис. 3 и 4, семейства дискретны, и мы не обнаружили промежуточных вариантов между ними. Вместе с тем, при исследовании полиморфизма гордеина в 116 образцах H. spontaneum из Ирана, Израиля, Сирии и Иордании [32–35] нами были обнаружены блоки компонентов идентичные вариантам, присутствующим в культурном ячмене и относящиеся к 8 семействам гордеина А и 11 семействам гордеина В (табл. 5). Полученные результаты позволяют сделать вывод, что существующий полиморфизм гордеинов А и В у культурного ячменя является следствием двух процессов: с одной стороны, интрогрессии аллелей (вариантов блоков компонентов) гордеин-кодирующих локусов от H. spontaneum в результате спонтанной гибридизации при распространении культурного ячменя, с другой стороны – накоплением мутаций в генах локусов Hrd A и Hrd B у H. vulgare.
Таблица 5.
Число вариантов блоков компонентов семейств гордеинов А и В, присутствующих в H. vulgare и обнаруженых в образцах H. spontaneum из Ирана, Израиля, Сирии и Иордании
| Страна | Семейства гордеинов А и В | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AI | AIII | AIV | AV | AVII | AIX | AXI | AXII | BI | BII | BIII | BIV | BV | BVI | BVII | BIX | BXII | BXIII | BXVII | |
| Иран | 3 | 1 | 3 | 1 | 0 | 0 | 1 | 2 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 1 |
| Израиль | 7 | 1 | 1 | 3 | 1 | 3 | 0 | 0 | 1 | 1 | 1 | 3 | 1 | 3 | 1 | 0 | 0 | 5 | 0 |
| Сирия | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| Иордания | 2 | 0 | 0 | 1 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0 |
Настоящая статья не содержит каких-либо исследований с использованием в качестве объекта животных.
Настоящая статья не содержит каких-либо исследований с участием в качестве объекта людей.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Dakir El.-H., Ruiz M.-L., Garcia P. et al. Genetic variability evaluation in Moroccan collection of barley, Hordeum vulgare L., by means of storage proteins and RAPDs // Genet. Res. and Crop Evol. 2002. V. 49. P. 619–631.
Созинов А.А., Нецветаев В.П., Григорян Э.М. и др. Картирование локусов Hrd у ячменя (Hordeum vulgare L. emed. Vav. et Bach.) // Генетика. 1978. Т. 14. № 9. С. 1610–1619.
Поморцев А.А., Нецветаев В.П., Попереля Ф.А и др. Идентификация шестого локуса, контролирующего синтез гордеина у озимого ячменя // Докл. ВАСХНИЛ. 1983. № 1. С. 7–11.
Нецветаев В.П., Образцов И.С., Созинов А.А. Картирование локуса Hrd G в хромосоме 5 ячменя // Молекулярные механизмы генетических процессов. V Всесоюзн. симп. Тез. докл. М.: Наука, 1983. С. 110.
Shewry P.R., Faulks A.J., Pickering R.A. et al. The genetic analysis of barley storage proteins // Heredity. 1980. V. 44. P. 383–389.
Blake T.K., Ullrich S.E., Nilan R.A. Mapping of the Hor-3 locus encoding D hordein in barley // Theor. Appl. Genet. 1982. V. 63. P. 367–372.
Поморцев А.А., Нецветаев В.П., Созинов А.А. Полиморфизм культурного ячменя (Hordeum vulgare L.) по гордеинам // Генетика. 1985. Т. 21. № 4. С. 629–639.
Miflin B.J., Shewry P.R. An introduction to the extraction and characterisation of barley and maize proteins // Techniques for the separation of barley and maize proteins / Eds Miflin B.J., Shewry P.R. Luxembourg: EEC, 1982. P. 13–25.
Ладогина М.П., Поморцев А.А., Нецветаев В.П. и др. Наследование и генетический контроль компонентов глютелина у ячменя (Hordeum vulgare L.) // Генетика. 1988. Т. 24. № 2. С. 316–323.
Shewry P.R., Parmar S. The Hrd F (Hor 5) locus encodes γ-type hordeins // Barley Genet. Newslet. 1987. V. 17. P. 32–34.
Shewry P.R., Parmar S., Franklin J. et al. Mapping and biochemical analysis of Hor 4 (Hrd G), a second locus encoding B hordein seed proteins in barley (Hordeum vulgare L.) // Genet. Res. Camb. 1988. V. 51. P. 5–12.
Созинов А.А., Попереля Ф.А. Методика вертикального дискового электрофореза в крахмальном геле и генетический принцип классификации глиадинов. Одесса: 1978. 16 с.
Поморцев А.А., Лялина Е.В. Оценка сортовой принадлежности и сортовой чистоты семян ячменя методом электрофоретического анализа запасных белков зерна // Методическое пособие к практикуму “Белковые маркеры для генетической паспортизации и улучшения геномов растений хозяйственно ценных видов”. М.: 2011. 86 с.
Kreis M., Shewry P.R., Forde B.G. et al. Structure and evolution of seed storage proteins and their genes with particular reference of those of wheat, barley and rye // Oxford Surveys Plant Mol. Cell Biol. 1985. № 2. P. 253–317.
Shewry P.R., Bunce N.A.C., Kreis M. et al. Polymorphism at the Hor 1 locus of barley (Hordeum vulgare L.) // Biochem. Genetics. 1985. V. 23. № 5/6. P. 391–403.
Bunce N.A.C., Forde B.G., Kreis M. et al. DNA restriction fragment length polymorphism of hordein loci: Application to identifying and fingerprinting barley cultivars/ / Seed Sci. Techn. 1986. V. 14. P. 419–429.
Саянова О.В., Мехедов С.Л., Буркитбаев Е.М. и др. Структурная организация генов С-гордеинов ячменя // Генетика. 1994. Т. 30. № 6. С. 749–755.
Sayanova O., Pomortsev A., Ladogina M. et al. Analysis of C-hordein deficient mutant of barley // Barley Genet. Newslet. 1992. V. 22. P. 53–56.
Kanazin V., Ananiev E., Blake T. Variability among members of Hor-2 multigene family // Genome. 1993. V. 26. P. 397–403.
Shewry P.R., Parmar S., Franklin J. Recombinant within the Hor 2 locus // Barley Genet. Newslet. 1988. V. 18. P. 31–33.
Wiberg A. Genetical studies of spontaneous sources of resistance to powdery mildew in barley // Heredity. 1974. V. 77. P. 89–148.
Mozeman J.G., Nevo E., Zohary D. Resistance of Hordeum spontaneum collected in Israel to infection with Erysiphe graminis hordei // Crop Sci. 1983. V. 23. P. 1115–1119.
Mastebroek H.D., Balkema-Boomstra A.G., Gaj M. Genetic analysis of powdery-mildew (Erysiphe graminis f. sp. hordei) derived from wild barley (Hordeum vulgare ssp. spontaneum) // Plant Breeding. 1995. V. 114. P. 121–125.
Provan J., Russell J.R., Booth A. et al. Polymorphic chloroplast simple sequence repeat primers for systematic and population studies in the genus Hordeum // Mol. Ecol. 1999. № 8. P. 505–511. https://doi.org/10.1046/j.1365-294X.1999.00545.x
Harlan J.R., Zohary D. Distribution of wild wheats and barley // Sciences. 1966. V. 153. № 3741. P. 1074–1080. https://doi.org/10.1126/science.153.3740.1074
Willcox G. Archeobotanical evidence for the beginnings of agriculture in Southwest Asia // The Origins of Agriculture and Crop Domestication. Proceed. of Harlan Symp. Aleppo, Syria. 10–14 May. 1997. P. 25–38.
Wendorf F., Schild R., El Hadidi N. et al. Use of barley in the Egyptian late paleolithic // Science. 1979. V. 205. P. 1341–1347. https://doi.org/10.1126/science.205.4413.1341
Zohary D., Hopf M. Domestication of Plants in the Old World. The Origin and Spread of Cultivated Plants in West Asia, Europe, and Nile Valley. Oxford: Clarendon Press, 1988. 245 p.
Поморцев А.А. Исследование полиморфизма гордеинов у ячменей Эфиопии // Генетика. 2001. Т. 37. № 10. С. 1371–1382.
Поморцев А.А., Калабушкин Б.А., Ладогина М.П. и др. Геногеография и закономерности распространения аллельных вариантов в трех гордеинкодирующих локусах ярового ячменя на территории бывшего СССР // Генетика. 1994. Т. 30. № 6. С. 806–815.
Поморцев А.А., Калабушкин Б.А., Бланк М.Л. и др. Изучение естественного отбора в искусственных гибридных популяциях ярового ячменя // Генетика. 1996. Т. 32. № 11. С. 1536–1544.
Поморцев А.А., Лялина Е.В. Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L.) и дикого (H. spontaneum C. Koch) ячменя в Иране (как части Дуги Плодородия) // Генетика. 2016. Т. 52. № 10 С. 1146–1158. https://doi.org/10.7868/S001667581610009X
Поморцев А.А., Болдырев С.В., Лялина Е.В. Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L.) и дикого (Hordeum spontaneum C. Koch) ячменя в Иордании (как части Дуги плодородия) // Генетика. 2019. Т. 55. № 3. С. 290–299. https://doi.org/10.1134/S0016675819030135
Поморцев А.А., Болдырев С.В., Лялина Е.В. Аллельное разнообразие гордеин-кодирующих локусов Hrd A и Hrd B у культурного (Hordeum vulgare L.) и дикого (Hordeum spontaneum C. Koch) ячменя в Сирии (как части Дуги плодородия) // Генетика. 2018. Т. 54. № 11. С. 1275–1284. https://doi.org/10.1134/S0016675818110127
Поморцев А.А., Рубанович А.В., Ковалева О.Н., Лялина Е.В. Аллельное разнообразин гордеин-кодирующих локусов Hrd A и Hrd B у дикого (Hordeum spontaneum C. Koch) и культурного (Hordeum vulgare L.) ячменя в Израиле и Палестине // Генетика. 2019. Т. 55. № 11. С. 1298–1311. https://doi.org/10.1134/S0016675819110092
Дополнительные материалы отсутствуют.