

Генетика, 2021, T. 57, № 6, стр. 674-681
Цитогенетическая дифференциация популяций мышей Apodemus peninsulae Северного и Южного Прибайкалья по добавочным хромосомам
Ю. М. Борисов 1, А. А. Калинин 1, З. З. Борисова 1, И. В. Крищук 2, Б. И. Шефтель 1, *
1 Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
119071 Москва, Россия
2 Научно-производственный центр по биоресурсам, Национальная академия наук Беларуси
220072 Минск, Беларусь
* E-mail: borissheftel@yahoo.com
Поступила в редакцию 15.05.2020
После доработки 25.12.2020
Принята к публикации 12.01.2021
Аннотация
Проведено исследование кариологической изменчивости мышей Apodemus peninsulae в ряде новых пунктов Иркутской обл. и в Бурятии. Выявлены различия по вариантам систем макро- и микро- В‑хромосом между мышами данного вида с территории Иркутской обл. и из Бурятии. Это позволило отнести их к разным популяциям со своей историей происхождения. Результаты работы подтверждают, что при цитогенетической дифференциации популяций A. peninsulae определяющее значение имеют вариации по числу микро-В-хромосом.
Первые сведения о добавочных хромосомах появились в начале XX в. В 1906 г. Е. Уильсоном [1] было открыто наличие в кариотипах насекомых “экстра хромосом”. Позднее в исследовании на кукурузе был предложен термин “В-хромосомы” [2], чтобы отличить их от А-хромосом. В-хромосомы особенно хорошо и полно изучены у насекомых и растений.
В ранних обзорах зарубежных авторов даются подробные сведения о видовом разнообразии растений и животных (главным образом, беспозвоночных), несущих в кариотипах В-хромосомы [3–5]. С продолжением цитогенетических изучений число видов, имеющих дополнительные структуры, постоянно пополняется. К настоящему времени В-хромосомы описаны в большинстве таксонов эукариот.
У млекопитающих они отмечены у 85 видов. Одиннадцать видов из них показывают различия в размере и морфологии B-хромосом [6, 7]. В хромосомных наборах многих видов встречаются точечные (или микро) В-хромосомы с неясной морфологической структурой, поскольку они столь малы, что невозможно различить положение центромеры. Например они выявлены в кариотипах мышей A. peninsulae [8–10], лисиц Vulpes vulpes [11], крыс рода Thomomys [12]. У млекопитающих чаще всего отмечены мелкие и средние добавочные хромосомы. В зависимости от положения центромеры, по морфологии различают: мета-, субметацентрические и акроцентрические В-хромосомы.
У Perognatus baileyi выявлен сложный внутри- и межпопуляционный хромосомный полиморфизм [13]. В пяти исследованных популяциях этого вида диплоидное число хромосом изменялось в пределах 46–48, 46–49, 48–50, 49–54, 49–55 за счет непостоянного числа (1–10) мелких акро- и метацентрических В-хромосом и 1–2 средних субметацентрических В-хромосом. Межпопуляционный полиморфизм по В-хромосомам описан у американских хомячков Reithrodontomys megalotis [14, 15]. Число мелких В-хромосом у них варьирует от ноля до семи, и диплоидные числа соответственно изменяются от 42 до 49. Наименьшее число добавочных хромосом найдено в популяциях на юго-западе США, наибольшее – в Юте и прибрежных районах Калифорнии.
Большая часть видов млекопитающих с добавочными хромосомами принадлежит к отряду грызунов (Rodentia) – 55 видов. Например из 20-ти видов мышей рода Apodemus дополнительные В‑хромосомы выявлены у шести видов [6, 7]. Одним из них является восточно-азиатская мышь A. peninsulae. Этот вид распространен на Алтае, в Сибири, Прибайкалье и Забайкалье, в Хабаровском крае, в Приморье, на о-вах Сахалин и Хоккайдо, на северо-востоке Казахстана, в Северной Монголии, в Корее, в Северо-Восточном и Центральном Китае до Цинхай-Тибетского плато на западе и провинции Юньнань на юге [16, 17].
В индивидуальных кариотипах большинства особей этого вида отмечены добавочные хромосомы, исключения редки [10]. Они имеют разные формы и количество: их число может доходить до 30 [10]. В настоящее время известно, что только у восточноазиатских мышей, обитающих на Сахалине и на о-ве Стенина (залив Петра Великого, Приморский край) в кариотипе нет B-хромосом [9], однако на о. Хоккайдо (Япония) B-хромосомы у восточно-азиатских мышей есть [8]. Наибольшее число В-хромосом (до 30) выявлено у зверьков этого вида, обитающих в Сибири [9, 10]. У этого вида найдено три типа изменчивости по В-хромосомам: межпопуляционной, межиндивидуальной и внутрииндивидуальной (мозаицизм). Эти положения обусловливают перспективность использования восточноазиатских мышей в качестве модельного объекта при исследовании феномена В-хромосом в кариотипах млекопитающих. Это может внести ясность в понимание природы, роли и происхождения этих загадочных элементов геномов эукариот [10, 18, 19].
Регион Байкала представляет собой уникальный научный полигон для исследований разнообразия, а также стабильности и изменчивости систем В-хромосом Аpodemus peninsulae в различных эколого-географических провинциях Прибайкалья [20]. Ранее нами была изучена изменчивость макро- и микро-В-хромосом на южном побережье оз. Байкал и в Северной Монголии [20], но северная часть региона оставалась практически не изученной, особенно окрестности г. Северо-Байкальска (бассейн Верхней Ангары). Здесь наблюдаются значительные эколого-климатические отличия от южного побережья Байкала с его умеренным и влажным климатом [21].
Цель настоящей работы – анализ кариотипических особенностей и в первую очередь системы добавочных В-хромосом восточноазиатских мышей, обитающих в окрестностях оз. Байкал.
МАТЕРИАЛЫ И МЕТОДЫ
В 2016–2017 гг. были отловлены 68 восточноазиатских мышей в девяти пунктах Иркутской обл. и Республики Буртия. Номера пунктов указаны на карте (рис. 1). Отловы в основном проводились в 2017 г., а в окрестностях дер. Сарма (пункт 4) – в 2016 г. Семь пунктов отлова находятся на берегу оз. Байкал, а два пункта – в Иркутской обл., в долине р. Ангара, были удалены от него. Пункт №1 – окрестности пос. Илир Братского р‑на Иркутской обл. (с.ш. 55°09′18′′, в.д. 100°35′34′′) был расположен в 480 км от побережья Байкала, пункт №2 – окрестности пос. Барда, Эхирит-Булагатский р-н, Иркутская обл. (с.ш. 52°49′51′′, в.д. 104°24′33′′) находился в 90 км от Байкала. Три пункта были расположены на северном и северо-западном берегах Байкала: пункт №3 – окрестности пос. Душкачан, побережье Ангарского Сора, Северо-Байкальский р-н, Иркутская обл. (с.ш. 55°51′06′′, в.д. 109°40′58′′); пункт №4 – дер. Сарма, Ольхонский р-н, Иркутская обл. (с.ш. 53°06′02′′, в.д. 106°50′09′′); пункт №5 – пос. Голоустное (с.ш. 52°02′23′′, в.д. 105°24′37′′) и пос. Нижний Кочергат (с.ш. 52°08′15′′, в.д. 105°16′26′′) Иркутского р-на Иркутской обл. Четыре пункта были расположены на Южном Берегу Байкала: пункт №6 – окрестности г. Байкальск, Иркутская обл. (с.ш. 51°31′02′′, в.д. 104°09′22′′), пункт №7 – окрестности пос. Танхой, Кабанский р-н, Республика Бурятия (с.ш. 51°33′16′′, в.д. 105°06′54′′); пункт №8 – окрестности г. Бабушкин, Кабанский р-н, Республика Бурятия (с.ш. 51°43′, в.д 105°52′); пункт №9 – окрестности пос. Истомино, Кабанский р-н, Республика Бурятия (с.ш. 52°07′57′′, в.д. 106°17′45′′). Количество экземпляров восточноазиатских мышей, отловленных в каждом пункте, приведено в табл. 1.
Рис. 1.
Пункты отлова Apodemus peninsulae в Байкальском регионе. 1 – окрестности с. Илир, Братский р-н, Иркутская обл.; 2 – дер. Барда, Эхирит-Булагатский р-н, Иркутская обл.; 3 – окр. пос. Душкачан, Северо-Байкальский р-н, Бурятия; 4 – пос. Сарма, Ольхонский р-н, Иркутская обл.; 5 – окр. пос. Б. Голоустное и пос. Нижний Кочергат, Иркутская обл.; 6 – окр. г. Байкальск, Иркутская обл.; 7 – окр. пос. Танхой-Выдрино, Кабанский р-н, Бурятия; 8 – окр. г. Бабушкин, Кабанский р-н, Бурятия; 9 – окр. пос. Истомино, Кабанский р-н, Бурятия.
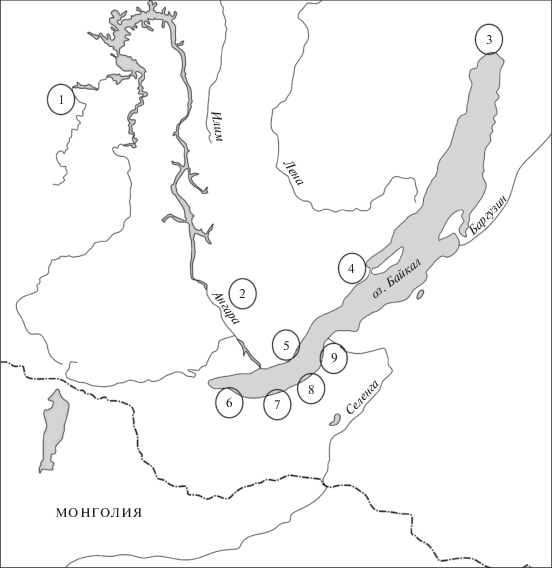
Таблица 1.
Формулы вариантов системы B-хромосом (Bs) Apodemus peninsulae Байкальского региона
| № пункта | № особи | Пол | 2n | Bs | Формула Bs |
|---|---|---|---|---|---|
| 1 | 1 | ♀ | 51 | 3 | 3.0.0.0.0 |
| 2 | ♂ | 51 | 3 | 2.0.0.1.0 | |
| 2 | 3 | ♂ | 51 | 3 | 3.0.2.1.0 |
| 4 | ♀ | 51 | 3 | 0.0.1.2.0 | |
| 5 | ♀ | 51 | 3 | 0.0.1.2.0 | |
| 6 | ♀ | 54 | 6 | 0.0.2.1.0 | |
| 7 | ♀ | 51 | 3 | 0.0.3.0.0 | |
| 3 | 8 | ♂ | 50 | 2 | 0.0.2.0.0 |
| 9 | ♂ | 49 | 1 | 1.0.0.0.0 | |
| 10 | ♀ | 51 | 3 | 1.0.1.1.0 | |
| 11 | ♂ | 51 | 3 | 0.1.1.1.0 | |
| 4 | 12 | ♂ | 50 | 2 | 2.0.0.0.0 |
| 13 | ♂ | 49 | 1 | 1.0.0.0.0 | |
| 14 | ♀ | 50 | 2 | 1.0.0.1.0 | |
| 15 | ♀ | 50 | 2 | 1.0.0.1.0 | |
| 16 | ♀ | 51 | 3 | 0.0.1.2.0 | |
| 17 | ♀ | 51 | 3 | 0.0.1.2.0 | |
| 5 | 18 | ♂ | 51 | 3 | 0.0.3.0.0 |
| 19 | ♂ | 49 | 1 | 0.0.0.1.0 | |
| 20 | ♂ | 49 | 2 | 1.0.0.1.0 | |
| 21 | ♀ | 50 | 2 | 0.0.0.2.0 | |
| 22 | ♀ | 50 | 2 | 0.0.0.2.0 | |
| 23 | ♂ | 50 | 2 | 0.0.1.1.0 | |
| 24 | ♀ | 50 | 2 | 0.0.0.2.0 | |
| 6 | 25 | ♀ | 53 | 5 | 0.2.0.0.3 |
| 26 | ♂ | 54 | 6 | 0.1.0.0.5 | |
| 27 | ♀ | 54 | 6 | 0.1.2.0.3 | |
| 28 | ♀ | 54 | 6 | 0.2.0.0.4 | |
| 29 | ♀ | 55 | 7 | 0.2.0.0.5 | |
| 30 | ♀ | 56 | 8 | 0.1.0.0.7 | |
| 31 | ♀ | 59 | 11 | 0.0.1.0.10 | |
| 32 | ♂ | 61 | 13 | 0.1.0.0.10 | |
| 33 | ♀ | 60 | 12 | 0.0.0.0.12 | |
| 34 | ♂ | 60 | 12 | 0.1.1.0.10 | |
| 35 | ♂ | 60 | 12 | 0.2.1.0.9 | |
| 36 | ♂ | 61 | 13 | 0.1.1.0.11 | |
| 37 | ♂ | 55 | 7 | 0.2.0.0.5 | |
| 38 | ♂ | 57 | 9 | 0.0.2.0.7 | |
| 39 | ♂ | 58 | 10 | 0.1.1.0.8 | |
| 40 | ♂ | 58 | 10 | 0.2.0.0.8 | |
| 41 | ♂ | 58 | 10 | 0.2.0.0.8 | |
| 42 | ♂ | 59 | 11 | 0.0.3.0.8 | |
| 43 | ♀ | 59 | 11 | 0.0.3.0.8 | |
| 44 | ♂ | 59 | 11 | 0.1.2.0.8 | |
| 45 | ♀ | 54 | 6 | 0.2.3.0.1 | |
| 46 | ♀ | 60 | 12 | 0.2.0.0.10 | |
| 7 | 47 | ♂ | 52 | 4 | 0.2.0.0.2 |
| 48 | ♀ | 52 | 4 | 0.3.0.0.1 | |
| 49 | ♀ | 57 | 9 | 0.2.0.0.7 | |
| 50 | ♂ | 58 | 10 | 0.2.0.0.8 | |
| 51 | ♂ | 54 | 6 | 0.0.1.0.5 | |
| 52 | ♂ | 57 | 9 | 0.0.1.0.8 | |
| 53 | ♂ | 58 | 10 | 0.0.1.0.9 | |
| 54 | ♂ | 58 | 10 | 1.0.1.1.7 | |
| 55 | ♀ | 59 | 11 | 0.0.2.0.9 | |
| 8 | 56 | ♀ | 53 | 5 | 0.1.1.0.3 |
| 57 | ♀ | 54 | 6 | 0.2.0.0.4 | |
| 58 | ♂ | 55 | 7 | 0.0.1.0.6 | |
| 59 | ♂ | 57 | 9 | 0.2.1.0.6 | |
| 60 | ♀ | 59 | 11 | 0.0.0.0.11 | |
| 61 | ♀ | 59 | 11 | 0.0.1.0.10 | |
| 62 | ♀ | 59 | 11 | 0.1.0.0.10 | |
| 63 | ♀ | 60 | 12 | 0.1.0.0.11 | |
| 64 | ♀ | 60 | 12 | 0.2.0.0.10 | |
| 9 | 65 | ♂ | 59 | 11 | 0.1.1.0.9 |
| 66 | ♂ | 60 | 12 | 0.2.1.0.9 | |
| 67 | ♂ | 59 | 11 | 0.2.1.0.8 | |
| 68 | ♂ | 54 | 6 | 0.0.1.0.5 |
Примечание. Локализацию мест сбора см. на рис. 1.
Хромосомные препараты готовили прямым методом из клеток костного мозга мышей с предварительным введением внутрибрюшинно 0.5 мл 0.04%-ного раствора колхицина [22]. Препараты делали также из клеток селезенки с применением технических приемов, предложенных Е.Ю. Крысановым с коллегами [23]. Для характеристики кариотипа каждого животного использовали анализ не менее, чем 20 метафазных пластинок. Работа с препаратами осуществлялась с помощью микроскопа “Leica D-500”. Подсчет хромосом выполняли в программе LAS V3.8. При обработке материала собственных и литературных данных мы учитывали только вариант с модальным числом В-хромосом мышей, отловленных в различных пунктах Байкальского региона (табл. 1). В таблице приведены формулы вариантов системы B-хромосом (Bs) кариотипов изученных мышей Apodemus peninsulae Байкальского региона.
В настоящей работе нами использована система классификации В-хромосом и формула цифрового кодирования [10]. Для сопоставления индивидуальных характеристик набора В-хромосом мышей их разделяли по размерно-морфологическим классам и записывали для каждой особи в виде формулы цифрового кодирования (табл. 1). Таким образом было выделено пять классов В‑хромосом по их размерам и морфологическим характеристикам. Согласно данной формуле: I класс – двуплечие крупные В-хромосомы, равные по размерам 1–8 парам А-хромосом; II класс – двуплечие В-хромосомы средних размеров, равные 9–16 парам А-хромосом; III класс – мелкие двуплечие В-хромосомы, равные 17–23 парам А-хромосом; IV класс – мелкие акроцентрические В‑хромосомы, равные 17–23 парам А-хромосом; V класс – точечные (или микро) В-хромосомы с неясным положением центромеры, в несколько раз мельче самых мелких А-хромосом набора. B‑хромосомы I–IV классов мы объединяли в группу макрохромосом, а B-хромосомы V класса вошли в группу микрохромосом.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
У всех 68 особей A. peninsulae был определен основной стабильный набор А-хромосом данного вида. Он представлен 23 парами акроцентрических аутосом и двумя половыми хромосомами: крупной акроцентрической Х-хромосомой и мелкой акроцентрической Y-хромосомой (рис. 2). Вариабельная часть кариотипа у этих мышей представлена 1–12 дополнительными В-хромосомами, среди них: 1–4 макрохромосом и 1–11 микрохромосом (табл. 1, рис. 2–4). Таким образом, кариотипы исследованных восточноазиатских мышей имеют от 55 до 62 хромосом. Кариотипы мышей, отловленных в двух пунктах, удаленных от оз. Байкал, объединяет отсутствие микрохромосом (класс V) и наличие отдельных особей с тремя большими двуплечими В-хромосомами (класс I), такие особи не были встречены в пунктах, расположенных на берегу Байкала. В кариотипах зверьков, отловленных в пунктах, расположенных на северном и северо-западном берегах Байкала (№ 3–5), также не найдены микрохромосом (класс V), но у всех особей выявлено от одной до трех макрохромосом (I–IV классов). Таким образом, в кариотипах 24-х восточноазиатских мышей, отловленных к северу от Байкала, микро-В-хромосомы не встречены.
Рис. 2.
Кариотип особи № 9 из пункта 3: а – 48 акроцентрических хромосом основного набора, б – крупная метацентрическая В-хромосома I класса.

Рис. 3.
Кариотип особи № 8 из пункта 3: а – 48 акроцентрических хромосом основного набора, б – две мелкие метацентрические В-хромосомы III класса.

Рис. 4.
Кариотип особи № 65 из пункта 9: а – 48 акроцентрических хромосом основного набора, б – одна средняя, одна мелкая метацентрические В-хромосомы III класса и девять микро-В-хромосом.

Напротив, в кариотипах всех 44-х особей, отловленных в четырех пунктах (№ 6–9) на южном берегу Байкала, отмечено от одной до двенадцати микрохромосом (V класс). Причем у подавляющего большинства (36 особей) найдено пять и больше таких В-хромосом (табл. 1). Кроме того, у всех этих особей, кроме одной, выявлены от одной до трех макро-В-хромосом всех классов.
Ранее нами [20] уже были описаны кариотипы 30 особей A. peninsulae из трех пунктов Южного Прибайкалья, из окрестностей г. Байкальск (1984, 1988 гг.), г. Танхой (2002, 2003 гг.) и г. Бабушкин (1984 г.). То есть исследования проводились в тех же самых пунктах, что и теперь (рис. 1, пункты 6–8). Все эти особи имели от 1–2 макро- и до 10 микро-В-хромосом [20]. То есть, формула В-хромосом в одних и тех же пунктах, не смотря на значительный временной интервал между исследованиями принципиально не менялась. Это дает нам основание предполагать, что формула В‑хромосом – это устойчивый признак, характеризующий географические популяции.
В настоящей работе мы выявили значительные отличия по вариантам системы В-хромосом у мышей A. peninsulae северного Прибайкалья и бассейна Ангары от мышей южного Забайкалья. Это позволяет отнести их к разным популяциям со своей историей происхождения. Популяция Южного Прибайкалья A. peninsulae сходна по своей структуре макро- и микро-В-хромосом с еще более южной популяцией бассейна Селенги (окрестности монастыря Амар-Хийд, Северная Монголия) [20]. А популяции Прибайкалья и бассейна Ангары, в которых выявлены только макро-В-хромосомы, сходны с популяцией A. peninsulae из окрестностей пос. Новоангарск на самом севере ее ареала, в 40 км от устья реки Ангары. Там также были выявлены от пяти до девяти двуплечих макро-В-хромосом, и среди 15 изученных мышей не было мышей с микро-В-хромосомами [10].
Ранее при изучении кариотипов восточноазиатских мышей из разных областей Сибири было обнаружено, что они отличаются по наборам В‑хромосом, это позволило высказать предположение, что системы макро и микро-В-хромосом могут являться интегрирующим и дифференцирующим признаком популяций мышей Сибири [10, 20]. Результаты данного исследования подтверждают, что В-хромосомы могут быть использованы как маркеры различных популяций восточноазиатской мыши. Определяющее значение имеет соотношение микро и макро-В-хромосом.
Авторы благодарны М.Ю. Борисову, Е.С. Гайдученко, Т.А. Мышлявкиной за помощь в отлове мышей A. peninsulae на южном побережье оз. Байкал. Благодарим А.Ю. Александрову за помощь в приготовлении хромосомных препаратов восточноазиатских мышей в ряде пунктов исследования.
Работа выполнена при финансовой поддержке РФФИ, проект № 16-08-1185.
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Авторы заявляют, что у них нет конфликта интересов.
Список литературы
Wilson E.B. Note on the chromosome-groups of metapodius and banasa // Biol. Bull. 1907. V. 12. № 5. P. 303–313.
Randolph L.A. Types of supernumerary chromosomes in maize // Anatomical Record. 1928. V. 41. P. 102.
Battaglia E. Cytogenetics of B-chromosomes // Caryologia. 1964. V. 17. № 1. P. 245–299.
Jones R.N., Rees H. B Chromosomes. L., N.Y., Paris: Acad Press, 1982. 266 p.
Beukeboom L.W. Bewildering Bs: An impression of the 1 B-chromosome conference // Heredity. 1994. V. 73. № 3. P. 328–335.
Vujosevic M., Blagojević J. B-chromosomes in populations of mammals // Cytogenet. and Genome Res. 2004. V. 106. № 2–4. P. 247–256. https://doi.org/10.1159/000079253
Vujošević M., Rajičić M., Blagojević J. B Chromosomes in populations of mammals revisited // Genes. 2018. V. 9. № 10. P. 487. https://doi.org/10.3390/genes9100487
Hayata J. Chromosomal polymorphism caused by supernumerary chromosomes in field mouse, Apodemus giliacus // Chromosoma. 1973. V. 42. № 4. P. 403–414.
Kartavtseva I.V., Roslik G.V. A complex B chromosome system in the Korean field mouse Apodemus peninsulae // Cytogenetic and Genome Res. 2004. V. 106. № 2–4. P. 271–278. https://doi.org/10.1159/000079298
Borisov Y.M., Zhigarev I.A. B Chromosome system in the korean field mouse Apodemus peninsulae Thomas 1907 (Rodentia, Muridae) // Genes. 2018. V. 9. № 10. P. 472. https://doi.org/10.3390/genes9100472
Беляев Д.К., Волобуев В.Т., Раджабли С.И., Трут Л.Н. Полиморфизм и мозаицизм по добавочным хромосомам у серебристо-черных лисиц // Генетика. 1974. Т. 10. № 2. С. 58–67.
Patton J.L., Sherwood S.W. Genome evolution in pocket gophers (genus Thomomys). I. Heterochromatin variation and speciation potential // Chromosoma. 1982. V. 85. P. 149–162.
Patton J.L. A complex system of chromosomal variation in the pocket mouse, Perognathus baileyi Merriam // Chromosoma. 1972. V. 36. № 3. P. 241–255.
Shellhammer H.S. Chromosome polymorphism in California populations of harvest mice // J. Mammal. 1968. V. 49. № 4. P. 726–731.
Shellhammer H.S. Supernumerary chromosomes of the harvest mouse, Reithrodontomys megalotis // Chromosoma. 1969. V. 27. № 1. P. 102–108.
Борисов Ю.М., Жигарев И.А., Шефтель Б.И. В-хромосомы восточноазиатских мышей (Apodemus peninsulae Thomas, 1907 (Rodentia, Muridae)) на восточных склонах Цинхай-Тибетского плато (КНР) // Генетика. 2020. Т. 56. № 10. С. 1184–1188. https://doi.org/10.31857/S0016675820090039
Громов И.М., Ербаева М.А. Млекопитающие фауны России и сопредельных территорий. Зайцеобразные и грызуны. СПб: ЗИН РАН, 1995. 522 с.
Trifonov V.A., Perelman P.L., Kawada S.I. et al. Complex structure of B-chromosomes in two mammalian species: Apodemus peninsulae (Rodentia) and Nyctereutes procyonoides (Carnivora) // Chromosome Res. 2002. V. 10. № 2. P. 109–116.
Rubtsov N.B., Borisov Y.M. Sequence composition and evolution of mammalian B chromosomes // Genes. 2018. V. 9. № 10. P. 490–496. https://doi.org/doi:10.3390/genes9100490
Борисов Ю.М., Шефтель Б.И., Сафронова Л.Д., Александров Д.Ю. Устойчивость популяционных систем В-хромосом восточноазиатской мыши Apodemus peninsulae Прибайкалья и Северной Монголии // Генетика. 2012. Т. 48. № 10. С. 1190–1199.
Климат и растительность Южного Прибайкалья // Сб. науч. трудов / Ред. Ладейщиков Н.П., Моложников В.Н. Новосибирск: Наука, сиб. отд., 1989. 151 с.
Орлов В.Н. Кариосистематика млекопитающих. Цитогенетические методы в систематике млекопитающих. М.: Наука, 1974. 208 с.
Крысанов Е.Ю., Демидова Т.Б., Шефтель Б.И. Простой метод приготовления препаратов хромосом мелких млекопитающих // Зоол. журн. 2009. Т. 88. № 2. С. 234–238.
Дополнительные материалы отсутствуют.