

Геоэкология. Инженерная геология, гидрогеология, геокриология, 2020, № 2, стр. 28-38
НЕКОТОРЫЕ ГЕОЭКОЛОГИЧЕСКИЕ АСПЕКТЫ УСТОЙЧИВОСТИ ЛЁССОВЫХ МАССИВОВ (БИОТИЧЕСКИЕ ФАКТОРЫ ФОРМИРОВАНИЯ ЛЁССОВОГО ПСЕВДОКАРСТА)
А. А. Лаврусевич 1, *, И. А. Лаврусевич 1, **
1 Национальный исследовательский Московский строительный государственный университет (НИУ МГСУ)
129337 г. Москва, Ярославское шоссе, 26, Россия
* E-mail: lavrusevich@yandex.ru
** E-mail: ivanlavrusevich@gmail.com
Поступила в редакцию 15.10.2019
После доработки 6.12.2019
Принята к публикации 16.12.2019
Аннотация
Лёссовый псевдокарст – очень стремительно развивающийся (месяцы, сутки и даже часы), сложный и многофакторный процесс разрушения лёссовых массивов. В формировании лёссового псевдокарста важную роль играет широкий диапазон биотических факторов: фитогенных, микогенных, зоогенных и микробиогенных. Это деятельность землероющих животных, растительности, почвенной фауны, перепончатокрылых, почвенных микроорганизмов и водорослей. Показано, что роль биотических факторов в некоторых случаях имеет двоякое значение. Они могут способствовать зарождению, развитию и многократному увеличению псевдокарстового процесса. Это дезинтеграция (нарушение целостности лёссового массива) за счет расширения корнями ксерофитной растительности первичных вертикальных трещин, связанных с генезисом лессовых пород, что приводит к проникновению воды на десятки метров в глубину лёссового массива и последующее его разрушение. Создание густой сети разветвленных каналов роющими животными или ходами червей-норников до глубины 15 м также способствует разрушению массива. И, напротив, влияние биотических факторов может быть направленно на сдерживание и предотвращение развития лёссового псевдокарста. Например, формирование на поверхности лёссовых пород плотной, карбонатизированной, слабо водонепроницаемой корочки, связанной с жизнедеятельностью водорослей, грибов и актиномицетатов; строительство на входе в псевдокарстовую полость дикими пчелами гнезд, состоящих из сотовых ячеек, плотно связанных с поверхностью лёссового грунта и предохраняющих псевдокарстовую полость от проникновения влаги.
ВВЕДЕНИЕ
Проблема исследования механизма лессового псевдокарста существует более 135 лет. Впервые мы находим описание необычных форм разрушения лессовых массивов в монументальном труде Ф. Рихтгофена “Китай” (1877 г.) [30]. Именно Ф. Рихтгофен впервые отметил, что рост оврагов в лессовых массивах идет не снизу вверх от базиса эрозии, а сверху вниз – от места проникновения воды в лессовый массив в средней или верхней части лессового массива. Немецкий исследователь Вальтер фон Кнебель использовал термин “псевдокарст” (нем. pseudoverkarstung) при описании миграции воды в лавовых потоках Исландии [28]. Л. Фуллер в 1922 г. описывал псевдокарстовые формы рельефа Китая, сопровождая все эффектными фотографиями [27]. В СССР лессовый псевдокарст изучали М.М. Решеткин, К.И. Лисицин, И.Г. Глухов и др. [10].
Занимаясь на протяжении всей жизни проблемой лесса в различных регионах мира, безусловно, не мог обойти эту проблему и Николой Иванович Кригер. В 1975 г. в трудах ПНИИИСа Н.И. Кригер публикует свой труд “Лессовый псевдокарст”, где рассматривает историю и анализ становления термина как у нас в стране, так и за рубежом [10]. Там же он впервые приводит классификацию многообразных форм лессового псевдокарста. Позже выходит целая серия статей, посвященных морфологии, динамике и методам исследований лессового псевдокарста [11–13 и др.]. В отечественной нормативной литературе термин “лессовый псевдокарст” впервые встречается в п.1.8 ВСН 33-2.2.06-86 [3].
Лессовый псевдокарст – результат гидромеханического, гравитационного, биологического и физико-химического зональных процессов, протекающих в недоуплотненных лессовых породах при их избыточном (как правило, техногенном) увлажнении и приводящих к образованию форм рельефа, очень напоминающих типичные карстовые (воронки, колодцы, провалы, пещеры, тоннели, останцы и др.) [16].
Для образования и развития лессового псевдокарста необходим обширный комплекс абиотических, биотических и техногенных факторов (геоморфологических, климатических, почвенных, литологических, биологических и др.). Лессовый псевдокарст развивается, если имеются лессовые породы мощностью не менее 3 м, с достаточно высокой пористостью и низкой влажностью, с невысоким содержанием карбонатов (до 12–15%), а также возможностью эпизодического проникновения воды в лессовый массив. Источником воды являются атмосферные осадки (климатический фактор), утечки воды из гидротехнических сооружений, водонесущих коммуникаций, не дозированный полив полей и др. [13]. Лессовый псевдокарст формируется на территориях с радиационным индексом сухости R/Lr от 1 до 2 (отношение радиационного баланса R [ккал/ см2] к скрытой теплоте испарения Lr) [11].
Вопрос о роли организмов в процессе формирования лессового псевдокарста уже поднимался в литературе [13, 23 и др.]. Указывалось, что в развитии лессового псевдокарста большое значение имеет деятельность позвоночных, высших растений, сине-зеленых и других водорослей.
ВЛИЯНИЕ РАСТИТЕЛЬНОСТИ
Защитная противоденудационная роль наземной растительности хорошо известна. При высокой степени покрытия растительностью, почва даже на склонах крутизной до 30о может быть защищена от смыва. Большое влияние на скорость развития лессового псевдокарста оказывает растительность в ложбинах временных потоков (балках, оврагах), где кратковременность стояния воды не мешает ее вегетации. В течение года (особенно весной) лучше всего защищают почву от эрозии многолетние травы. Почвозащитная эффективность однолетних культур в разные фазы их развития различна. До их созревания, как правило, происходит полив. Нередко, именно после проведения неоднократных поливов или после уборки этих культур и вспашки почвы возникает большая опасность проявления эрозии. По мере развития растительного покрова она уменьшается, так как сомкнутый травостой – надежный амортизатор ударов капель.
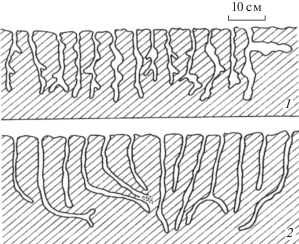
Важный биогенный фактор, влияющий на водопроницаемость почвы, – рост и разложение корней растений. Как известно, корни предпочитают распространяться по порам, ходам насекомых и червей, пустотам от разложившихся корешков, избегая как уплотненных участков почвы, так и больших пустот. Закупоривание пор корнями может временно значительно уменьшить водопроницаемость лессового массива. В целом можно констатировать, что растения с густой и неглубоко проникающей корневой системой закрепляют верхний слой почвы, снижая вероятность развития лессового псевдокарста, тогда как растения, обладающие мощными длинными корнями, могут способствовать развитию псевдокарста, подготавливая пути проникновения поверхностных вод в толщу лессового массива (рис. 1).
Рис. 1.
Юго-западный Таджикистан, Яванская долина. Глубокая эрозионная промоина (шириной до 40 см) на склоне псевдокарстового оврага вдоль корня полыни Artemisia baldshuanika Krasch et Zapr.

Действительно, при вертикальной ориентировке “корнеходов” происходит повышенная инфильтрация воды с поверхности, следовательно, увеличивается влажность массива. При наличии на 1 м2 котлована более 6 корнеходов значительно снижается прочность лессового массива [17]. Например, корни полыни Artemisia baldshuanika Krasch et Zapr и верблюжьей колючки Alhagikirghisorum Schrenk, достигая толщины в несколько сантиметров, проникают на глубину до 10–15 м.
На такую же глубину по данным Г.С. Сиворакши и А.А. Ядрова проникают толстые стержневые корни фисташки Ristacia vera L. – дикорастущего плодового дерева [24, 20 ]. Особое место среди древесных растений занимает Робиния псевдоакация (Robinia pseudoacacia L.), принадлежащая к семейству бобовых рода Робиния (рис. 2, 3).
Рис. 2.
Корни Robinia pseudoacacia L. в вертикальной трещине в лессовом массиве, обусловленной генезисом и структурой лесса. Сербия, правый берег р. Дунай.

Рис. 3.
Одиноко растущее дерево Robinia pseudoacacia L. При увеличении видно, что вертикальная трещина, рассекающая лессовый массив, приурочена к его основному корню [16].

М.А. Михеева отмечает [18], что для Робинии псевдоакации характерен наименьший водный дефицит и относительно повышенное водоудерживание, высокая засухо-, жаро- и газооустойчивость по сравнению с другими древесными растениями. Оценка жизненного состояния Робинии псевдоакации по соответствующим методикам показала, что именно у нее отмечается наибольший процент здоровых особей. Робиния обладает далеко разветвляющейся корневой системой, что обусловливает ее устойчивость к ветру.
Робиния псевдоакация происходит из Северной Америки, где растет на влажных известняковых почвах в лиственных лесах от Пенсильвании до Джорджии и Оклахомы. Широко известна в культуре стран Западной Европы, в некоторых областях ее также как и у нас ошибочно считают аборигенным видом. Авторы наблюдали посадки Робинии псевдоакации на лессовых склонах в провинции Ганьсу и Шеньси (Китай) [16].
Листопадное дерево до 30 м высотой, с диаметром ствола в 30–40 см, с просвечивающей, раскидистой, ажурной кроной, состоящей из обособленных ярусов. Растет быстро, особенно в первое десятилетие, хорошо приживается при искусственных посадках на террасированных склонах различной экспозиции и переносит обрезку ветвей и пересадку, очень светолюбива и засухоустойчива, в сплошных насаждениях высоко выносит крону, мало требовательна к плодородию и влажности почвы, однако на сырых, тяжелых почвах чаще страдает от морозов и гибнет.
По данным Е.И. Захаровой Робиния псевдоакация – весьма неприхотлива, однако полученные в результате экспериментов данные подтверждают мнение многих авторов о гибели Робинии лжеакации на мокрых почвах при избытке застойной влаги [1, 8, 21].
При искусственных посадках Робинии псевдоакации лессовый склон на глубину до 25 м будет “армирован” корнями этого растения и, безусловно, укрепит массив, предотвращая его разрушение [25]. Однако если растение по тем или иным причинам погибло, то оставшийся после разложения корня вертикальный ход будет являться идеальным водопроводом на глубину до 25 м.
Например, при обследовании псевдокарстового провала (диаметр 5.5 м, видимая глубина 8 м) около автодорожного моста Сиань-Биньсянь (рис. 4) было обнаружено, что его дно неровное и завалено обломками лессовой породы, а в стенках встречены фрагменты ствола и ветвей погибшей Робинии псевдоакации. Отчетливо выделялся обнаженный корень погибшей Робинии псевдоакации диаметром до 8 см на глубине 8.9 м (см. рис. 3). При более детальном обследовании провала можно было с уверенностью предположить, что первопричина формирования конкретно этого провала – инфильтрация атмосферных осадков вдоль основного корня Робинии псевдоакации. Видимо образование описанного провала произошло именно по этому сценарию.
Рис. 4.
Склон, осложненный серией провалов, в северной части магистрального моста автодороги Сиань-Биньсянь. Стрелкой указан обследованный провал [16].

При исследовании пещеры в лессовых породах близ селения Сурдук (Сербия) авторы [29] неоднократно ссылаются на то, что пещера такой величины (высота 12 м и диаметр до 3.5 м) могла сохраниться при наличии многочисленных деревьев рода Робинии псевдоакации.
За возникшими первоначально трещинами последовал воронкообразный провал. Однако дальнейшее разрушение псевдокарстовой пещеры прекратилось, благодаря наличию многочисленных деревьев Робинии псевдоакции, корневая система которых прочно скрепила массив вокруг возникшего провала. Сербские исследователи приводят аналогичные противооползневые мероприятия, описанные Л. Бииком с соавторами для корневой системы Алеппской сосны (Aleppo pine) на юго-востоке Испании [25].
ВЛИЯНИЕ ДЕЯТЕЛЬНОСТИ БЕСПОЗВОНОЧНЫХ
Сильно изменяет инженерно-геологические свойства лессовых грунтов развитие почвенной фауны. Дождевые черви разрыхляют почвенные горизонты, увеличивая водопроницаемость, в результате их жизнедеятельности меняется состав почвы, разрушаются почвенные минералы. Глубина проникновения червей-норников в грунт достигает 8–10 м. Н.И. Кригер при инженерно-геологических исследованиях в Рудном Алтае прослеживал вертикальные ходы червей в лессе до глубины 10–15 м и более [14]. Предполагают, что гумусовый горизонт почвы был неоднократно пропущен через пищеварительный тракт дождевых червей. Черви не любят кислых почв, предпочитая карбонатные (известковые). Копролиты дождевых червей обычно богаты известью, что способствует кальцификации почвы, образованию агрегатов и известковых конкреций. Ходы-норки червей имеют своеобразное строение – их открытые части длиной от нескольких дециметров до 1 м (редко больше) сменяются участками, заполненными рыхлым грунтом с экскрементами червей. Это защищает червей от жуков-хищников и минимизирует затраты на выброс лишнего грунта на поверхность. О роли дождевых червей в проницаемости почвы могут свидетельствовать следующие данные: норки глубинных дождевых червей представляют собой ходы диаметром 8–10 мм в количестве до 500 и более на 1 м2 [14]. По данным Д.Г. Звягинцева и др. [9], под 1 м2 поверхности почвы общая длина ходов червей превышает 1 км, иногда достигая 8 км. При такой “пористости” интенсивность впитывания воды значительно увеличивается. Поскольку активность почвенной фауны тесно связана с временами года, постольку ее значение, как фактора водопроницаемости, также носит сезонный характер. Черви плохо переносят засуху, высокие и низкие температуры. С началом сезона активной деятельности фауны количество пор в почве увеличивается, а после ее спада поры начинают разрушаться и заплывать. В среднеазиатском лессе при низкой влажности менее 10% породы оказывают сопротивление внедрению в нее организмов, поэтому распространение их ограниченно. Следует отметить повышенное содержание в лессах Средней Азии СаСО3, что также препятствует развитию червей. Черви служат источником питания многочисленных норных животных. После завершения жизненного цикла черви представляют собой богатый белковый субстрат, быстро утилизируемый микроорганизмами. В кишечнике червей обитают микромицеты, грибы и бактерии.
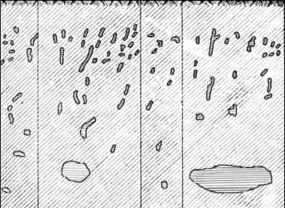
Менее важна, но достаточно интересна, деятельность мокриц рода Oniscoidea [9]. Там, где в массивах лессовых пород не живут дождевые черви, их роль выполняют мокрицы (рис. 5). Их колонии составляют в среднем 80 особей на 1 м2. Их норки проникают на глубину 0.40–0.50 м, иногда достигая глубины 0.8 м. В результате своей жизнедеятельности мокрицы за год могут вынести до 5 кг/м2 почвы.
Особый интерес вызвала находка (левый берег Нурекского водохранилища, Юго-Западный Таджикистан) на входе в псевдокарстовую полость плотной “оболочки”, образованной колонией земляных пчел семейства Apidae, полностью (мешкообразно) перекрывающей на глубину 0.4 м всю полость хода (рис. 6) [15]. Оболочка представляет собой плотное и водонепроницаемое известковистое образование темно-зеленого цвета, состоящее из множества тесно переплетенных трубочек-сот, как правило, запечатанных, внутри которых находятся перга и личинки пчел. Толщина такой оболочки от 5 до 8 см, глубина проникновения в лессовую породу 3–4 см, она с трудом режется ножом и отрывается от грунта. В данном случае мы, по всей видимости, имеем дело с явлением, когда организмы (перепончатокрылые) препятствуют развитию лессового псевдокарста, полностью перекрывая вход в псевдокарстовую полость [15].
Рис. 6.
Устье псевдокарстовой полости (а) и схема строения сформированной земляными пчелами “оболочки”, перекрывающей вход в нее (б). 1 – лесс, 2 – водонепроницаемое известковистое образование, состоящее из трубочек-сот. Первомайская впадина, район Нурекского водохранилища, Таджикистан. Фото А.А. Лаврусевича, 1984 г. (для масштаба в левой части снимка лежит крышка от объектива фотоаппарата Зенит).

Большую роль в преобразовании верхних горизонтов почв аридных и семиаридных зон играют муравьи и термиты, где их роль в перераспределении солевых отложений особенно велика. Практически нет участка в пустынных и полупустынных регионах, где не обитали бы один или несколько видов муравьев и термитов. В отличие от термитов и муравьев влажных тропических и субтропических регионов, которые вынуждены для спасения от воды селиться в дуплах или на ветвях деревьев, муравьи и термиты минеры аридных и семиаридных районов живут глубоко под землей. Их отвердевшие ходы встречаются на глубине 8 и даже 10 м [5, 7]. Наиболее часто встречающиеся виды: туркестанский термит (Hodotermes turkestanicus Jac) и средний сакаспийский (Hodotermes vagans Septentrionalis Jac). Среди муравьев это Mirmicasp, Catalyphisaenescens, Crematogaster, Teyramorium, род Messor и др. Целый раздел посвятил описанию муравьев знаменитый исследователь А.П. Федченко в своем произведении “Путешествие в Туркестан” [22]. При проникновении вглубь гнезда обнаруживаются многочисленные камеры до нескольких метров протяженностью и нескольких сантиметров высотой. В литературе приведен пример, когда к ножке муравья привязали тончайшую шелковую нить с целью измерить глубину ходов гнезда. 25-метровая нить вся исчезла в земле [23]. На глубине более 1 м количество камер, как правило, уменьшается. Сильная семья муравьев выносит на поверхность и наращивает вокруг гнезда до 50–75 м3 грунта, а ходы от центра гнезда к периферии тянутся на сотни метров [23]. Отдельные виды муравьев создают громадное количество гнезд, связанных друг с другом на площади нескольких гектаров. Известны случаи, когда крупные глыбы, под которыми поселились муравьи, постепенно утопали в почве [23]. На опытных площадях по изучению и наблюдению за термитами практически не было свободных участков от деятельности черепах, ящериц, змей, полевок и др. животных. “Свободная от деятельности землероев площадь на поверхности почвы (а также и в нижележащих слоях) оказалась бы весьма незначительной” [5, с. 161]. Термиты и муравьи поднимают с глубины влажные комочки глины, которыми они заделывают изъяны ходов внутри гнезда, и уплотняют ими свои подземные сооружения. Влажная глина находится выше УГВ в зависимости от капиллярной каймы на 2–3 м. Но, если глубина УГВ 20 м, то термитам приходится опускаться за ними до 18 м [5]. Стенки ходов муравьев и термитов уплотнены, совершенно гладкие, так что они являются идеальными водопроводами для поверхностных вод в глубокие грунтовые горизонты. В результате своей жизнедеятельности муравьи и термиты выносят значительные массы грунта с нижних горизонтов, что сопровождается изменениями химического, механического и петрографического состава поверхностных грунтов. В муравейниках и термитниках, как правило, отмечают повышенное содержание солей и гипса, pH, концентрацию важнейших биогенных элементов. На участках развития муравейников и термитников образуются провалы, в которые устремляется вода, проникающая вглубь массива, в результате образуются псевдокарстовые пустоты, трещины, обвалы и оползни. Таким образом, муравейники и термитники имеют важное инженерно-геологическое значение, так как способствуют формированию ослабленных зон в лессовых массивах, вызывая просадку, формирование трещин, поглощение воды и образование псевдокарста.
Отдельно скажем несколько слов о туркестанской степной черепахе (Nestudo horsfieldi), широко распространенной в пределах лессовых адыров и оказывающей существенное влияние на их устойчивость. Черепахи роют норы глубиной до 2 м. Количество особей на 1 га достигает сотен штук, которые своей роющей деятельностью иногда провоцируют разрушение дамб и оросительных каналов, уничтожают эфемерную растительность в пустынях, снижая ценность пастбищ [19].
ВЛИЯНИЕ ДЕЯТЕЛЬНОСТИ ПОЗВОНОЧНЫХ
Еще одно из широко распространенных явлений, в результате которого происходит интенсивное разрушение целостности верхних горизонтов пород, – деятельность так называемых землероющих животных, чрезвычайно распространенных в пределах развития лессовых пород. Наибольшие разрушения верхних горизонтов толщи пород производят грызуны (тушканчики, полевые мыши, суслики и др.). Большая их часть фитофаги, питающиеся исключительно растениями. В результате своей жизнедеятельности они создают сложную систему подземных ходов и камер с множеством выходов на поверхность. Ходы землероев-грызунов распределяются весьма неравномерно. Есть участки практически свободные от ходов, а нередко образуются скопления на больших участках (до 24 кротовин на 1 м2). В отдельных случаях длина ходов землероев достигает 50–80 м, при диаметре от 1 до 15 см [17]. Такое количество нор способствует быстрому перемещению воды из верхних слоев в нижние горизонты.
Среди насекомоядных позвоночных землероев в лессовых массивах следует отметить крота, ежа и землеройку. Эти животные питаются червями, личинками насекомых, моллюсками, но могут употреблять и растительную пищу. Крот роет неглубокие, как правило, горизонтальные норы небольшого до 6 см в диаметре. Часть почвы крот выбрасывает на поверхность, образуя небольшие, до 30 см в диаметре и по высоте холмики. На лугах кроты выбрасывают до 160 т/га земли в год. Н.А. Богословский отмечает наличие многочисленных кротовин в пористых и карбонатизированных лессовидных суглинках вдоль железнодорожной линии Москва–Савелово [2].
Среди зайцеобразных отметим пищуху (Ochotona), роющую норы большой протяженности в лессовых массивах, запасая на зиму корм из сухой травы и поедая собственные капролиты, чем она способствует увеличению щелочности почвы.
Широко развит в пределах лессовых массивов бухарский суслик (Сitellus fulvus), он роет наклонные в сторону склона норы глубиной до 8 м простого строения с обязательными вертикальными ходами и гнездовой камерой с тупиками-отнорниками небольшой протяженности. Возле нор образуются холмики земли (курганчики), которые выделяются более густой и разнообразной растительностью [6]. Большое количество подземных нор сравнительно большого диаметра (0.3–0.6 м), располагающихся преимущественно в нижней части лессовых откосов, создают сравнительно крупные животные (дикобразы, барсуки, лисицы и др.) (рис. 7).
Рис. 7.
Развертка шурфа №180 в южной части Туткаульской впадины с ходами землероев (левый берег Нурекского водохранилища, Юго-Западный Таджикистан).
Склоны южной и юго-восточной экспозиции более интенсивно поражаются ходами и норами землероев, чем склоны северной и северо-западной экспозиции, так как на них быстрее сходит снежный покров, и они быстрее прогреваются весной, что способствует заселению землероев. Глубина поражения толщи лессовых пород ходами землероев достигает в пределах адырных возвышенностей 6 м, в пределах дна глубоких межадырных долин до 4 м. Наибольшей нарушенностью отличаются при этом верхние слои лессовых пород до глубины 2–2.5 м.
ВЛИЯНИЕ ДЕЯТЕЛЬНОСТИ МИКРООРГАНИЗМОВ
Изучая псевдокарстовые полости, было отмечено практически повсеместное присутствие в них различных микроорганизмов. Взятые на анализ образцы “корочки лессовых пород” с опытных участков исследований показали присутствие целого ряда микроскопических организмов. Это позволило сделать предположение, которое впоследствии было доказано, о существенной роли микроорганизмов в устойчивости и разрушении лессовых массивов. На сегодняшний день имеется достаточное количество данных о инженерно-геологическом значении микроорганизмов. Помимо того, что они – основные агенты биологического преобразования верхней части литосферы, которая является зоной жизни человечества, они оказывают существенное влияние на устойчивость уже возведенных инженерных сооружений, нередко вызывая их интенсивное разрушение. Этому вопросу в последнее время уделяется все большее внимание [2, 6 и др.]. Особенность микроорганизмов – способность к очень активному размножению. Например, при благоприятных условиях через 20–30 мин бактерии удваивают свое первоначальное количество клеток. Таким образом, за одни сутки может появиться до 60 поколений одной клетки. Если принять массу одной клетки за 2.5 ⋅ 10–12 г, за 2–4 суток может образоваться биомасса порядка 1010 т [6]. Основные составляющие микроорганизмов – бактерии, грибы, актиномицеты и водоросли.
Количество микроорганизмов в верхних слоях грунта огромно. Суммарная длина грибного мицелия на 1 г почвы составляет километры, а максимальная 35 км; гифы актиномицетов – сотни метров. “Вес живой биомассы на 1 га для бактерий может изменяться от сотен до нескольких тысяч килограммов, а для грибов – от сотен килограммов до десятков тонн на 1 га” [9, с. 29]. Мицелий грибов агрегирует частицы и структурирует грунт.
Споры бактерий весьма устойчивы к неблагоприятным условиям и высушиванию, что связано с наличием особого слоя в клеточной стенке. Клеточные стенки снаружи бывают окружены слизистым веществом, так называемые капсулы, слои, чехлы или влагалища [9]. Актиномицеты – бактерии, обладающие очень тонким мицелием, который проникает в почвенный субстрат, а часть развивается над субстратом, это так называемый воздушный мицелий, характеризуются способностью к формированию ветвящегося мицелия, что определяет их сходство с грибами. Микроорганизмы принимают самое активное участие в деструкции минералов почвообразующих пород. Развиваясь на поверхности лессового массива грибы, выделяют массу спор и продуктов жизнедеятельности (метаболитов). Это до 10% внеклеточных выделений в виде щелочи, уксусной, щавелевой и лимонной кислот с концентрацией от 0.01 до 3%, кремнийорганические соединения и хелаты.
Однако микроорганизмы участвуют не только в рассеивании элементов. Очень часто карбонатные минералы в лессах, имеют биогенное происхождение (рис. 8). Кальцит образуется при осаждении кальция углекислотой, выделяемой при метаболизме почвенных организмов.
Рис. 8.
Фрагмент плотной известковой корочки в псевдокарстовой форме на поверхности лесса, образованной в результате жизнедеятельности микроорганизмов (водорослей, актиномицетов, бактерий).

Изучая поверхность лессовых массивов свободных от растительности в различных регионах мира, авторы обратили внимание на плотную, темного цвета корочку, покрывающую повсеместно открытые участки лессовой породы. Были отобраны образцы, результаты обработки (посевы Агар-агар) на почвенном факультете МГУ под руководством профессора Г.М. Зеновой представлены в табл. 1.
Таблица 1.
Численность бактерий, актиномицетов и грибов на 1 г субстрата (по результатам Г.М. Зеновой)
| Численность, КОЕ/г | ||
|---|---|---|
| бактерий | актиномицетов | грибов |
| 2.5 × 100 000(10) | 2.4 × 10 000(102 × 102) | 3.8 × 1000 (103) |
Наблюдения показали, что повсеместное присутствие плотной корочки на открытых участках лессовой породы обнаруживается спустя 1 год после обнажения лесса. Корочка долгоживущая, плотная и обладает способностью противостоять эрозии. В псевдокарстовых пустотах на поверхности пород также обнаружена плотная корочка. Она формируется в результате жизнедеятельности, в основном, почвенных водорослей. Подавляющее большинство водорослей, населяющих почву, представляют собой микроскопические формы. Однако, образуя при помощи слизи и выростов иногда целые колонии, они становятся видны на поверхности почвы невооруженным глазом как различного рода оформленные разрастания – кожистые или войлокообразные пленки, слизистые слоевища, либо как позеленение почвы (цветение почвы), обусловленное массовым развитием микроскопических форм. В период цветения почвы на 1 см2 поверхности может развиваться до 40 млн клеток, биомасса которых может достигать 1.5 т/га [9].
Численность водорослей и их биомасса зависят, в первую очередь, от влажности, освещения и солевого режима. Важная особенность синезеленых и зеленых водорослей – способность накапливать в слоевище соединения кальция, что обусловливает ее твердость и способность противостоять эрозии [4].
В наибольшем количестве видов и примерно в равных соотношениях в пробах, отобранных из опытных участков, представлены сине-зеленые (отдел Cyanophita), зеленые (отдел Clorophita) и диатомовые водоросли (отдел Bacillariophyta). Весьма характерны, хотя и не так многочисленны, виды желто-зеленых водорослей (отдел Hantophita). Почвенные водоросли обладают свойством “эфемерности” – способностью быстро переходить из состояния покоя к активной вегетации и наоборот. Наземные корочки водорослей, высыхающие в сухие периоды, пробуждаются при увлажнении уже через несколько часов.
При изучении лессового псевдокарста Средней Азии, Китая, Сербии и России сине-зеленые и зеленые почвенные водоросли обнаружены в образцах, отобранных со дна, стен и сводов пещер и тоннелей, протяженность некоторых из них более 50 м. Количество особей водорослей увеличивалось от потолка к дну тоннеля, что объясняется, по-видимому, большей влажностью на дне полости.
Эрозия – одна из главных причин разрушения почвенного покрова. Поэтому факты, показывающие, что водоросли могут повышать устойчивость почвы против водной и ветровой эрозии представляют большой интерес. По наблюдениям Бооса, который наблюдал развитие сине-зеленых водорослей в США, соотношение эрозии из мест под водорослевой пленкой и с почвы без нее составило 1:22 [26].
Как правило, в подземных псевдокарстовых полостях среди микроорганизмов преобладают водоросли, в то время как на открытой поверхности лессовых массивов грибы и актиномицеты. Видимо, водоросли более жизнеспособны, несмотря на отсутствие достаточного количества света во влажной среде подземных пустот.
ВЫВОДЫ
По наблюдениям авторов, биотические факторы развития лессового псевдокарста весьма существенны. Именно первичное разрушение лессового массива в результате жизнедеятельности организмов и растительности приводит к формированию псевдокарстовых форм рельефа. Диапазон воздействий весьма широк: в результате метаболизма микроорганизмов изменяется кислотность, щелочность, формируются кремнийорганические соединения в толще грунтов, землероющие и растения разрушают грунтовую толщу, формируя идеальные водотоки в глубину массива.
Однако, механизм воздействия почвенных водорослей на поверхности эродированных или эрозионноопасных почв двоякий. С одной стороны, слизистые вещества чехлов и клеточных оболочек склеивают почвенные частицы, а переплетающиеся нити водорослей механически скрепляют их. Накапливающиеся в слоевище водорослей соединения кальция обусловливают ее твердость и способность противостоять эрозии. С другой стороны, при проникновении воды через эту “заграждающую пленку” образуются промоины и другие псевдокарстовые формы больших размеров. Водорослевая пленка покрывает поверхность этих форм и вода, движущаяся по псевдокарстовым тоннелям, не растекается в пористом лессе, а движется концентрированным потоком, приобретая большую кинетическую энергию, и многократно увеличивает разрушающий эрозионный процесс в лессовом массиве.
Список литературы
Альбенский А.В., Дьяченко А.Е. Разведение быстрорастущих и ценных деревьев и кустарников. М.: Сельхозгиз, 1940. 223 с.
Болотина И.Н., А.В. Минервин, Усупаев Ш.Э. Микроорганизмы лессовых грунтов // Инженерная геология.1983. № 5. С. 47–54.
ВСН 33-2.2.06-86. Мелиоративные системы и сооружения. Оросительные системы на просадочных грунтах. Нормы проектирования. Основные положения. п. 1.8. URL: https://base.garant.ru/12182089/ (дата обращения: 22.09.2019)
Голлербах М.М., Штина Э.А. Почвенные водоросли, Л.: Изд-во АН СССР, 1969. 228 с.
Димо Н.А. Роль и значение термитов в жизни почв и грунтов Туркестана // Русский почвовед. 1916. № 7–10. С. 153–190.
Ерофеев В.Т., Смирнов В.Ф., Морозов Е.А. Микробиологическое разрушение материалов. Уч. пос. М.: АСВ, 2008. 123 с.
Жизнь животных. Членистоногие: трилобиты, хелицеровые, трахейнодышащие. Онихофоры / Под ред. М.С. Гилярова, Ф.Н. Правдина. 2-е, перераб. М.: Просвещение, 1984. Т. 3. 463 с.
Захарова Е.И. Влияние лесорастительных условий на сохранность и рост робинии лжеакации в лесных культурах при интродукции // Известия Оренбургского государственного аграрного университета. 2009. 1 (21). С. 58–61.
Звягинцев Д.Г., Бабьева И.П., Зенова Г.М. Биология почв. М: Изд-во МГУ, 2005. 445 с.
Кригер Н.И. Лессовый псевдокарст // Вопросы теории и методики инженерной геодинамики. Тр. ПНИИИС. М.: Стройиздат, 1975. Вып. 32. С. 25–50.
Кригер Н.И., Лаврусевич С.А. Происхождение лессового псевдокарста // Докл. АН Тадж. ССР. 1981. Т. 24. № 7. С. 444–448.
Кригер Н.И., Гранит Б.А. и др. Опыт комплексного изучения лессового псевдокарста для строительных целей (на примере окрестностей Алма-Аты) // Комплексные исследования в инженерных изысканиях для строительства. Тр. ПНИИИС. М.: Стройиздат, 1982. С. 50–63.
Кригер Н.И., Ботников В.И., Лаврусевич С.А. и др. Псевдокарст в лессовых породах // Геоморфология. 1983. № 3. С. 79–84.
Кригер Н.И. Инженерно-геологическое значение деятельности Eisenia Magnifica // Геоэкология. 1993. № 5. С. 95–103.
Лаврусевич А.А., Лаврусевич С.А. Псевдокарст прибрежной зоны Нурекского водохранилища // Докл. АН Тадж. ССР. 1986. Т. 29. № 3. С. 172–175.
Лаврусевич А.А., Крашенинников В.С., Лаврусевич И.А. Лессовый псевдокарст и опыт укрепления лессовых массивов и откосов искусственными посадками некоторых растений (на примере лессового плато КНР, провинции Ганьсу и Шеньси) // Инженерная геология. 2012. № 1. С. 48–58.
Ларионов А.К., Приклонский В.А., Ананьев В.П. Лессовые породы СССР и их строительные свойства. М.: Наука, 1959. 366 с.
Михеева М.А. Геоэкологическая оценка биоразнообразия и устойчивости древесных растений в условиях городской среды (на примере г. Воронежа): автореф. дис. на соиск. уч. ст. канд. географ. наук. Воронеж. 2009. 23 с.
Определитель земноводных и пресмыкающихся фауны СССР. Уч. пос. М.: Просвещение, 1977. 415 с.
Сиворакша Г.С. Фисташковые рощи Баба-Тага // Советские субтропики. 1939. № 2–3. С. 18–35.
Собинов А.М. Лесные культуры быстрорастущих ценных пород. М.–Л.: Гослестехиздат, 1947. 116 с.
Федченко А.П. Путешествие в Туркестан. СПб., М.: Тип. М. Стасюлевича, 1875. 160 с.
Халифман И.А. Муравьи. М.: Молодая гвардия. 1963. 304 с.
Ядров А.А. Миндаль, фисташка // Орехоплодные Таджикистана. Душанбе: Тадж. Госиздат, 1958. С. 287–296.
Beek L., Wint, J., Cammeraat L., Edwards J. Observation and Simulation of Root Reinforcement on Abandoned Mediterranean Slopes // Plant and Soil. 2005. V. 278. P. 55–74.
Booth W.E. Algae pioneers in plant successions and their importance in erosion control // Ecology. 1941. V. 22. № 1. P. 38–46.
Fuller M.L. Some unusual erosion features in the loess of China // Geographical Review. 1922. Review 12. P. 570–584.
von Knebel W. Die Bedeutung der Dolinen für die Entstehung von Tälern. In: Höhlenkunde mit Berücksichtigung der Karstphänomene. Die Wissenschaft (Sammlung Naturwissenschaftlicher und Mathematischer Monographien), Vieweg + Teubner Verlag, Wiesbaden, 1905, vol. 15.
Lukić T., Marković S., et al. The loess “cave” near the village of Surduk -an unusual pseudokarst landform in the loess of Vojvodina, Serbia. Acta Carsologica, Postojna. 2009. 38/2-3. P. 227–235.
Richthofen F. China. Bd. 1. Berlin, 1877. 726 p.
Дополнительные материалы отсутствуют.
Инструменты
Геоэкология. Инженерная геология, гидрогеология, геокриология