

Геохимия, 2020, T. 65, № 10, стр. 1009-1024
Моделирование устьичной, мезофильной и биохимической регуляции фотосинтеза сосны обыкновенной на основе экспериментальных данных
А. В. Соколов a, b, *, В. К. Болондинский c
a Институт геохимии и аналитической химии им. В.И. Вернадского РАН
119991 Москва, ул. Косыгина, 19, Россия
b Институт проблем передачи информации им. А.А. Харкевича РАН
127051 Москва, Б. Каретный пер., д. 19, стр. 1, Россия
c Институт леса КарНЦ РАН
185910 Петрозаводск, Пушкинская, 11, Россия
* E-mail: alexander.v.sokolov@gmail.com
Поступила в редакцию 10.04.2020
После доработки 29.04.2020
Принята к публикации 11.05.2020
Аннотация
Разделить сложный биохимический механизм фотосинтеза растения на составляющие (устьичная проводимость, проводимость мезофилла, фиксация углекислого газа в хлоропластах и дыхание) – трудная задача. Для ее решения мы использовали как экспериментальные данные, полученные в естественных условиях (временные ряды регистрируемых показателей фотосинтеза, транспирации и внешних условий), так и специально поставленный эксперимент – определение углекислотных компенсационных точек. Специальный метод обработки данных (сбалансированная идентификация) и соответствующая информационная технология позволили рассмотреть ряд моделей, объективно оценить значимость рассматриваемых гипотез, определить погрешности описания используемого набора данных. Эволюционный способ модификации моделей (от простого к сложному, с критерием отбора – погрешностью моделирования) позволил выбрать модель, сложность которой соответствует используемым экспериментальным данным. На основе построенной модели проведены расчеты ассимиляции CO2 лесными экосистемами и показателя эффективности использования воды растениями WUE (Water Use Efficiency).
ВВЕДЕНИЕ
Рост глобальной температуры, наблюдаемый в последние десятилетия, связывается многими исследователями со стремительным ростом содержания в атмосфере парниковых газов, и прежде всего СО2 [Grace 2000; IPCC 2013], темп роста концентрации которого в конце 2000-х годов достиг 2.20 ± 0.01 ppm/год (или 1.7% за год) и продолжает увеличиваться (https://www.esrl.noaa.gov/ gmd/ccgg/trends/global.html#global). Максимальная концентрация CO2, измеренная в обсерватории Мауна-Лоа (на Гавайях 19°29′ с.ш. 155°36′ з.д. на высоте 3400 м), в январе 2020 г. достигла 412.30 ppm. Не существует однозначного мнения касательно наблюдаемого роста концентрации СО2 в глобальном масштабе. Возможно, этот процесс связан с естественными колебаниями, и происходит не только за счет возрастающего объема поступлений из антропогенных источников (Peters et al., 2012).
Растительность активно обменивается СО2 с атмосферой, тем самым выполняя важную регулирующую функцию для климатической системы. На континентальных участках, на которые сильно влияют местные биосферные поглотители, концентрация CO2, особенно в дневные часы, существенно ниже. Лесные массивы северной и средней тайги являются мощным источником поглощения CO2, и моделирование фотосинтеза является важным элементом при оценке баланса СО2 в биогеоценозах и в биосфере в целом.
Для исследования процессов взаимодействия растительного покрова и атмосферы в настоящее время используется широкий спектр методов, включающих системы наземных и дистанционных наблюдений, а также построенные на их основе математические модели разного уровня сложности, различного пространственного и временного масштаба. Полученные модельные результаты одного уровня допускают обобщение на другой. Например, на основе модели динамики газообмена побега сосны можно получить оценки газообмена дерева, гектара леса и т.д. (переход на другой пространственный масштаб), в минуту, час, сутки т.д. (изменение временного масштаба).
Данная статья является продолжением серии работ по моделированию физиологических процессов сосны обыкновенной (Pinus sylvestris). В работе (Соколов и др., 2019а) мы подробно рассмотрели набор моделей динамики транспирации и выбрали их модель, соответствующую набору имеющихся экспериментальных данных. Выбор осуществлялся на основе метода сбалансированной идентификации (Соколов, Волошинов, 2018; Sokolov, Voloshinov, 2019; Соколов и др., 2019б) путем минимизации среднеквадратичной ошибка кроссвалидации. С использованием полученных результатов (временные ряды значений устьичной проводимости и дефицита воды в растении) и дополнительных наборов данных, полученных в результате пассивных (наблюдения в естественных условиях) и активных экспериментов, мы попытаемся описать путь СО2 в листе, оценить работу составных частей биохимического конвейера фиксации CO2.
В связи с многообразием вариантов описания функционирования фотосинтетического аппарата в настоящее время не удается достигнуть полной однозначности между совокупностью экспериментальных данных и объясняющей их моделью, так как всегда приходится выбирать между несколькими возможными вариантами модели, различающимися по сложности внутренних взаимосвязей (Laisk, Oja, 1998). Технология сбалансированной идентификации позволяет объективно оценить, насколько различные варианты модели соответствуют имеющемуся экспериментальному материалу, что позволяет определить точку прекращения дальнейшего усложнения модели.
При использовании технологии сбалансированной идентификации для относительно простых задач можно ограничиться заданием источников данных (файлов) и математическим описанием модели – остальную “работу” выполнит соответствующее ПО с привлечением (если это необходимо) дополнительных внешних (распределенных) вычислительных ресурсов. Конечно, чтобы грамотно сформулировать сложную математическую модель, требуется не только глубокие знания в предметной области, но и некоторое математическое образование. В этом случае для корректной постановки задачи идентификации (и ее переформулировки в виде файла задания для соответствующего ПО) требуется задание дополнительных параметров, например, параметров дискретизации, позволяющих заменить вариационную задачу поиска функций на конечномерную задачу математического программирования поиска наборов чисел (параметризаций искомых функций). Так как целью данной работы является моделирование конкретного процесса, многие технические подробности не приводятся. Мы будем использовать технологию сбалансированной идентификации как “черный ящик”, который позволяет объединить набор знаний (гипотез в виде уравнений) с набором экспериментальных данных, оценить “успешность” такого объединения (рассчитать погрешность моделирования данных) и найти неизвестные функции и параметры.
Технология сбалансированной идентификации объединяет в единое целое несколько областей математики, современное программное обеспечение и мощные распределенные вычислительные средства. Таким образом, появляется возможность корректно ставить и оперативно решать сложные задачи обработки экспериментальных данных с целью построения математических моделей. В том числе, только недавно появилась возможность обработать данные, полученные в 80-х годах прошлого века, лежащие в основе данной работы.
Вообще говоря, описание динамики содержания CO2 в различных частях листа предполагает использование аппарата дифференциальных уравнений. Однако, принимая во внимание характерное время моделирования (шаг 0.5 часа) и быстроту биохимических процессов, ограничимся квазиравновесным приближением, т.е. будем считать, что концентрации углекислого газа в растении определяются внешними и внутренними условиями в настоящий момент и не зависят от предыдущего момента.
РОЛЬ ВНЕШНИХ ФАКТОРОВ СРЕДЫ И ВНУТРЕННИХ ПАРАМЕТРОВ В МОДЕЛЯХ ФОТОСИНТЕЗА НА УРОВНЕ ЛИСТА
Процесс поглощения CO2 растением (и соответствующую модель) можно представить в виде потоковой диаграммы (рис. 1).
Рис. 1.
Потоки CO2 в листе. Схематическое представление модели конвейера поглощения CO2 (обозначения см. в табл. 1).
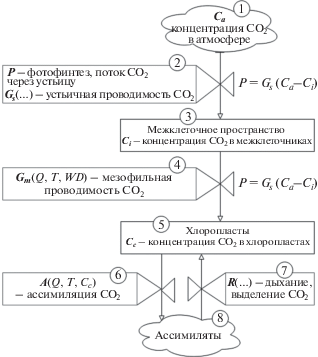
Углекислый газ из атмосферы (форма 1) через устьица (форма 2) поступает (диффундирует согласно закону Фика) в межклеточное пространство (форма 3), откуда молекулы СО2 диффундируют (согласно закону диффузии растворенных газов в жидкости) сквозь стенки мезофильных клеток (форма 4) к реакционным центрам в хлоропластах (форма 5), где и вступают в биохимический цикл фотосинтеза (форма 6), результатом которого является синтез ассимилятов (форма 8). На рис. 1 отображен и противоположный процесс – дыхание (форма 7), т.е. потребление богатых химической энергией веществ на биохимические процессы в различных частях листа. Подробное изучение дыхания не является целью данной работы, поэтому для простоты предположим, что появляющийся при этом углекислый газа попадает непосредственно в хлоропласты.
На рис. 1 приведены и различные некоторые внешние параметры, влияющие на скорости процессов, формулы и уравнения. Расшифровка, используемых здесь и далее обозначений, проведена в табл. 1.
Таблица 1.
Используемые обозначения и единицы измерения
| Обозначение | Расшифровка | Единицы измерения |
|---|---|---|
| t | Время с начала моделирования (с 20 июня) | час |
| h | Время суток | час |
| Е | Транспирация | ммоль м–2 с–1 |
| P | Наблюдаемый или нетто-фотосинтез | мкмоль м–2 с–1 |
| A | Ассимиляция CО2 (брутто-фотосинтез) | мкмоль м–2 с–1 |
| R | Дыхание (R =А – P) | мкмоль м–2 с–1 |
| Q | Солнечная радиация (освещенность) | мкмоль м–2 с–1 |
| VPD | Дефицит давления водяного пара в воздухе | Па |
| Т | Температура воздуха | °С |
| WD | Дефицит воды в растении | л |
| Сa | Концентрация CO2 в воздухе | мкмоль моль–1, ppm |
| Сi | Концентрация CO2 в межклеточниках | мкмоль моль–1, ppm |
| Сc | Концентрация CO2 в хлоропластах | мкмоль моль–1, ppm |
| Gs | Устьичная проводимость листа для CO2 | моль м–2 с–1 |
| Gm | Мезофильная проводимость листа для CO2 | моль м–2 с–1 |
| WUE | Эффективность использования воды | мкмоль CO2 (ммоль Н2О)–1 |
Рассмотрим более подробно элементы диаграммы на рис. 1
В основном, в процессе фотосинтеза используется CO2 атмосферы (форма 1). Небольшая часть углекислоты может доставляться к листьям вместе с ксилемным соком, но она обычно не превышает нескольких процентов.
Концентрация СО2 имеет выраженную суточную динамику, определяемую процессами фотосинтеза и дыхания (Lloyd et al., 2002). Суточный максимум атмосферной концентрации CO2 регистрируется в 6:00–7:00 ч местного времени, минимум – в вечерние часы (16:00–19:00 ч). Максимальная величина суточной амплитуды для CO2 наблюдается в июле около поверхности земли и составляет в среднем 21 ppm (Тимохина и др., 2014). Результаты, полученные в Швеции (Perttu et al., 1980) в конце августа в 120-летнем сосновом древостое (высота – 15.8 м) показали, что с 6 до 21 ч колебания концентрации СО2 во времени составляли 10–12 ppm от среднего уровня. Небольшое увеличение концентрации СО2 происходило с 20 до 24 ч. В дневное время у поверхности почвы концентрация СО2 на 10–15 ppm выше, чем в кронах.
Известно, что при концентрациях, близким к естественным, зависимость фотосинтеза сосны от величин концентрации СО2, близких к естественным, носит линейный характер (Голомазова, 1987, Щербатюк и др., 1992) и некоторое ее уменьшение днем снижает интенсивность процесса. В работе А.Г. Молчанова (1983) выявлено воздействие изменений естественной концентрации СО2 на световые кривые фотосинтеза. Снижение концентрации СО2 на 7% вызывало падение интенсивности фотосинтеза на плато до 30%.
Молекулы атмосферного СО2 через пограничный слой воздуха над листом, попадают через устьичные отверстия (форма 2) в межклеточное пространство (форма 3). Проводимость пограничного слоя для хвои сосны высока – 8.1 моль H2O м–2 с–1 (Stangl et al., 2019), т.е. сопротивление пограничного слоя достаточно мало, и им в расчетах можно пренебречь или включить его в устьичное.
Закон Фика
позволяет рассчитать устьичную проводимость для воды Gsw при известных значениях транспирации E, концентрации водяных паров в атмосфере Wa и насыщающей концентрации водяного пара в межклетниках при температуре листа Wi. В предположении равенства температуры листа и воздуха разность концентраций может быть заменена в первом приближении на дефицит давления водяного пара в воздухе VPD (Барри, Даунтон, 1987).
Из множества факторов, влияющих на устьичную проводимость, выделим солнечную радиацию Q и дефицит воды в растении WD, которые оказались ведущими факторами в модели водного режима (Соколов и др., 2019а). В летнее время (а именно, к этому сезону относится моделируемый период) температура в диапазоне 14–23°С оказывает меньшее влияние.
Аналогично водяному пару, поток CO2 через устьица (фотосинтез) P определяется разностью концентраций углекислоты в воздухе (Cа) и межклетниках (Ci) и устьичной проводимостью для углекислого газа (Gs). Согласно закону Фика для CO2:
причем устьичные проводимости для воды и углекислого газа связаны соотношениям (Jarvis, 1971):Поток молекул CO2 из межклеточного пространства (форма 3) через мезофилл (форма 4) в центры карбоксилирования в хлоропластах (форма 5) описывается законом диффузии растворенных газов в жидкости:
где Gm – мезофильная проводимость, Cс и Ci –концентрации CO2 в хлоропластах и межклеточниках.Мезофильная проводимость характеризует диффузию CO2 из межклеточного пространства листа до реакционных центров в хлоропластах и включение в биохимический цикл реакций темновой стадии фотосинтеза (Цельникер и др., 1993, Laisk, Loreto, 1996). В ряде работ Gm разделяют на две составляющих: физическую и химическую компоненты (Laisk, Oja, 1998). Первая зависит в основном от длины пути СО2 в жидкой фазе клетки и суммарной поверхности хлоропласта. Химическая составляющая определяется активностью карбоксилирующих ферментов, прежде всего РДФ-карбоксилазы. Анализ компонент мезофильного сопротивления показал, что доля физической компоненты значительно меньше, чем химической (Цельникер, 1978; Laisk, Oja, 1998). Мезофильную проводимость (обе составляющие) или, как ее иногда называют, внутреннюю проводимость связывают с первоначальным наклоном кривой зависимости ассимиляции A от Ci (Moldau, Kull, 1993; Laisk, Loreto, 1996; Eichelmann, Laisk, 1999; Eichelmann et al., 2004).
Однако, в последнее время все больше авторов термин “мезофильная проводимость” связывают только с диффузией CO2 через мезофилл листа: межклеточные воздушные пространства, клеточную стенку и межклеточную жидкость. То есть, мезофильную проводимость рассматривают как синоним диффузионной проводимости листа, а химическая часть, которая часто лимитирует фотосинтез, уже не входит в понятие мезофильной проводимости. Появление новых приборов и методик привело к значительному росту числа работ, где изучается мезофильная проводимость – за последние 15 лет число ссылок, связанных с различными аспектами мезофильной проводимости для CO2, увеличивалось экспоненциально, и превысило тысячу публикаций (Flexas et al., 2008, Stangl et al., 2019). Ошибки, вызываемые пренебрежением Gm при создании моделей фотосинтеза, значительно ухудшают их качество (Flexas et al., 2008).
Мезофильную проводимость в полевых условиях оценить значительно труднее, чем устьичную Gs, так как измерения Gm требуют одновременных измерений газообмена и изотопной дискриминации 13C (∆13C). Дискриминация может быть выведена из сигнатуры δ13C продуктов фотосинтеза, то есть растворимых сахаров листа, флоэмного содержимого (Hu et al., 2010; Ubierna, Marshall, 2011). Эти методы технически трудны, особенно в полевых условиях. Лишь в сравнительно недавно удалось с приемлемой точностью провести длительные одновременные измерения газообмена на уровне побега и 13C дискриминацию в столетнем древостое сосны обыкновенной на севере Швеции, получить почасовые оценки Gm параллельно с Gs, Р и E (Stangl et al., 2019).
Другим способом оценить концентрацию Сс, а затем подсчитать Gm, является проведение (активных) экспериментов по определению углекислотных компенсационных пунктов (УКП). Для этого побег помещают в замкнутую систему, где СО2 постепенно “выедается” и через некоторое время СО2-газообмен становится равным нулю. Концентрация углекислоты в камере, при которой это происходит, получила название УКП. В ряде моделей компенсационный пункт приравнивают к Сс(Лайск, 1977; Цельникер и др., 1993). У С4 растений УКП близок к нулю. У С3-растений, к которым относится сосна, в связи с фотодыханием концентрация CO2 на реакционных центрах (Сс) может быть значительно больше нуля (Цельникер и др., 1978, Eichelmann еt al., 2004), особенно при низкой освещенности (Голомазова, 1987).
Проводимости Gm и Gs реагируют на изменение окружающих условий по-разному. Описаны быстрые реакции Gm на некоторые переменные окружающей среды. Эти переменные включают интенсивность или качество света (Loreto et al., 2009; Campany et al., 2016), межклеточную концентрацию CO2 (Ci) (Flexas et al., 2007) и температуру листа (Warren, 2008).
При постоянной Gs рост Gm будет увеличивать эффективность использования воды (WUE) (Flexas et al., 2010), которая определяется как отношение нетто-ассимиляции углерода (Р) к потере воды в ходе транспирации (E). Это происходит из-за того, что увеличение Gm не имеет прямого влияния на транспирацию, но может увеличить фотосинтез, что приводит к увеличению отношения Р/E. В моделях WUE Gm часто вводится как постоянная, эмпирическая поправка в зависимости между Ci/Ca или экстраполируется на основании ее корреляции с Gs, хотя корреляция с Gs не всегда высокая (Klein et al., 2015).
Доставленный в хлоропласты СО2 в центрах карбоксилирования вступает в биохимический цикл фотосинтеза (форма 6). Здесь, с помощью накопленных (в реакциях световой фазы фотосинтеза) богатых энергией веществ, CO2 превращается (в ходе темновой фазы фотосинтеза) в глюкозу, а затем в крахмал (форма 8). Основные факторами, определяющими скорость фиксации CO2 являются ФАР (фотосинтетически активная солнечная радиация), температура и концентрация Cc.
ОБЪЕКТ ИССЛЕДОВАНИЯ И ЭКСПЕРИМЕНТАЛЬНЫЕ ДАННЫЕ
Исследования проводились на базе полевого стационара Института леса КНЦ РАН, расположенного в 50 км к северу от г. Петрозаводска (62°13′ с.ш. и 34°10′ в.д.). Район исследований, пробные площади и конкретный объект изучения – 55-летнее сосна обыкновенная (Pinus sylvestris L.) высотой 14 м описаны в (Болондинский, Кайбияйнен, 2003, Соколов и др., 2019а). В ходе исследования, наряду с транспирацией (Веселков, Тихов, 1984) и микроклиматическими параметрами, исследовался СО2-газообмен побегов в разных частях кроны (Болондинский, Кайбияйнен, 2003).
Для непрерывной автоматической регистрации СО2-газообмена побегов сосны использовалась 6-канальная автоматическая установка на базе инфракрасного газоанализатора Infralyt-4 (Junkalor, DDR). В дальнейшем ряд показателей СО2-газообмена уточнялся с помощью портативной газометрической системы LI-6200 (LI-COR, США). Измерения СО2-газообмена проводились с частотой 10 показаний/ч по каждому каналу. По ним рассчитывались средние получасовые значения. Интенсивность фотосинтеза рассчитывалась как на сухой вес, так и на площадь хвои (мкмоль СО2 м–2 с–1). Относительная погрешность измерений составляла 10–12%.
Ассимиляционные камеры изготавливались из прозрачного жесткого полиэтилена и устанавливались с южной стороны кроны на однолетних побегах 4–5-ой мутовок. В ходе измерения обеднение по CO2 воздуха в камере не превышало 10%. В летнее время, чтобы избежать перегрева хвои, мы использовали дополнительную систему для увеличения потока воздуха через камеры. При прокачке воздуха через камеру со скоростью 2.8 л/мин воздух сменялся в камере каждые 10 с, и хвоя не перегревалась больше, чем на 1°С.
Синхронно с фотосинтезом автоматически регистрировалась интенсивность приходящей солнечной радиации над пологом леса, а также внутри кроны деревьев.
Температура воздуха рядом с камерами регистрировалась термодатчиками прибора ТЭТ-2 (Гидрометприбор, Латвия), защищенными от попадания прямых солнечных лучей цилиндрическим жестяным экраном. Разницу между температурой хвои и температурой окружающего воздуха измерялась медно-константановыми термопарами (диаметр проволоки 0.1 мм). В каждом побеге закреплялись 4–5 термопар.
Относительную влажность воздуха измеряли гигрографом, показания которого контролировались психрометром Астмана. По данным температуры и относительной влажности воздуха рассчитывали дефицит упругости водяного пара воздуха.
Наиболее тщательно с минимальным количеством пропусков, не превышающим 1–2 суток в месяц, СО2-газообмен измерялся с 18 апреля по 30 октября. Для моделирования был выбран фрагмент данных с 20 июня по 23 июля. Всего 1632 измерения. Данные по E имеют пропуски (1537 точек). Отсутствие данных было вызвано, в основном, техническими проблемами. Выбранный период попадал на окончание интенсивного апикального роста ствола (27.05–20.06). После ослабления аттрагирующих свойств верхушечной меристемы часть углеводов транспортировалась в зону камбиального роста и к корням. В конце июня начался интенсивный радиальный рост, закончившийся в 3 декаду июля. В июле наблюдался также интенсивный рост хвои и ее дозревание. В результате, в исследуемый период у дерева, также, как и во время апикального роста в высоту, наблюдалась повышенная потребность в ассимилятах.
В рассматриваемый период (июнь-июль) в 50-см слое почвы содержалось 35–45 мм влаги, что несколько меньше, чем в мае – 65 мм. Последняя величина близка к полевой влагоемкости. Минимум влажности почвы (4–5%) регистрировался в зоне наибольшего распространения корней (10–30 см) обычно в июле-августе. Хорошая увлажненность почвы объясняется существованием довольно мощного пласта грунтовых вод (уровень грунтовых вод 1–1.5 м) и достаточным количеством осадков в период исследований – 92 и 392 мм за вегетацию и за год соответственно. В период, выбранный для моделирования, сосна не испытывала недостатка почвенной влаги (Кайбияйнен, 1990).
В начале июля в течение трех суток проводились измерение содержания CO2 в воздухе на разных высотах. Погрешность измерений не превышала 1%. Несмотря на сложную картину смены восходящих и нисходящих потоков СО2, связанных с изменением мощности истоков и стоков СО2 (фотосинтез, дыхание ветвей и ствола, выделение СО2 из почвы), на фиксированных высотах в определенный момент времени концентрация СО2 изменялась слабо. Полученные в результате значения приведены в табл. 2.
Таблица 2.
Концентрация углекислоты (мкмоль/моль) у почвы (h = 2 м) и над кронами (h = 21 м) в различное время суток в сосняке черничном свежем в начале июля 1984 г.
| Время | h = 2 м | h = 21 м |
|---|---|---|
| 2.00 | 350 ± 10 | 322 ± 2 |
| 6.00 | 350 ± 5 | 340 ± 1 |
| 8.00 | 320 ± 2 | 320 ± 1 |
| 11.00 | 314 ± 5 | 314 ± 1 |
| 17.00 | 333 ± 6 | 315 ± 1 |
| 20.00 | 338 ± 3 | 317 ± 1 |
| 23.00 | 341 ± 10 | 321 ± 2 |
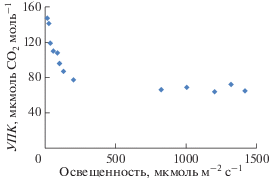
Углекислотный компенсационный пункт у сосны нами определялся как с помощью портативной системы Li-6200, так и на стационарной установке. Величину УКП мы приравнивали к концентрации CO2 на реакционных центрах (Сс). В зависимости от внешних факторов среды и периода вегетации величина УКП колебалась днем (при больших значениях радиации) в пределах 55–67 мкмоль/моль (июле-августе – 65–68 мкмоль м–2 с–1). Уменьшение величины освещенности, начиная с 200 мкмоль м–2 с–1, приводило к увеличению УКП (концентрации CO2) (рис. 2). Примерно такая зависимость отмечалась и другими исследователями (Голомазова, 1987).
ПРИМЕНЕНИЕ ТЕХНОЛОГИИ СБАЛАНСИРОВАННОЙ ИДЕНТИФИКАЦИИ ДЛЯ АНАЛИЗА МОДЕЛЕЙ ФОТОСИНТЕЗА
Будем постепенно шаг за шагом, гипотеза за гипотезой строить модель, соответствующую схеме на рис. 1. При этом особое внимание уделяется ошибке кроссвалидации – если при модификации модели (усложнении или упрощении) погрешность убывает – это серьезный аргумент в пользу изменения. В процессе модификации всякая последующая модель является развитием предыдущей. Найденные ранее решения используются как начальные приближения, что позволяет быстрее найти решения. Кроме того, такая постепенная (эволюционная) модификация является и необходимой – рассматриваемые постановки являются сложными двухуровневыми оптимизационными задачами (как правило, многоэкстремальными) и их решение требует значительных ресурсов. Поэтому поиск решения без “правдоподобного” начального приближения потребовал бы слишком больших вычислительных ресурсов и, кроме того, нет оснований предполагать, что найденное в этом случае решение (один из локальных минимумов оптимизационной задачи) будет иметь предметную интерпретацию, удовлетворяющую исследователя.
Формальные постановки задач идентификации не приводятся – предполагается, что используемая технология сбалансированной идентификации позволяет для заданной модели явления и используемого набора данных определить неизвестные функции и оценить погрешность построенной модели. Подробности можно найти в (Соколов, Волошинов, 2018; Sokolov, Voloshinov, 2019).
Для рассматриваемых ниже моделей основным множеством измерений
Для оценки погрешности моделирования используется процедура перекрестного оценивания с разбиением множества измерений D на 34 подмножества: каждое содержит измерения за одни сутки (до 48 точек). Полученную для такого разбиения методом сбалансированной идентификации среднеквадратичную ошибку кроссвалидации σcv, которую будем называть погрешностью моделирования данных, можно трактовать как меру предсказательной точности модели. Здесь и далее все погрешности даны в процентах от величины стандартного отклонения измерений (квадратный корень из дисперсии), что обеспечивает нормировку от 0% (при полном совпадении модельных значений со всеми измерениями) до 100% (для примитивной модели в форме постоянной функции, равной среднему значению измерений).
По единой методике для каждой модели найдем оптимально сбалансированное решение и рассчитаем погрешность моделирования измерений. Сначала рассмотрим модели простых функциональных зависимостей фотосинтеза от внешних факторов, затем воспользуемся временным рядом устьичной проводимости, рассчитанным в работе (Соколов и др., 2019а). Это позволит перейти к моделированию ассимиляции CO2 (брутто-фотосинтеза) и дыхания и привлечь дополнительные экспериментальные данные по динамике CO2 в атмосфере и по углекислотным компенсационным пунктам. Наконец, в последних трех моделях предпринимается попытка моделирования мезофильной проводимости.
Замечание. Чтобы избежать путаницы, в обозначениях функций используются дополнительные строчные буквы, соответствующие аргументам. Например, Pq(Q) или Pqvpd(Q, VPD).
Модель 1: Pq(Q)
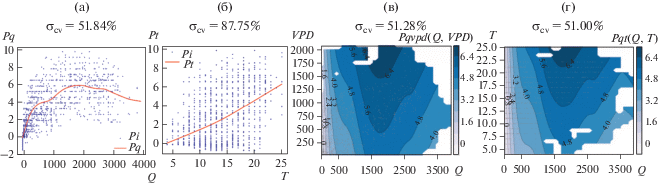
Результаты моделирования фотосинтеза как функции солнечной радиации приводятся на рис. 3а. Погрешность моделирования σcv = 51.84%.
Рис. 3.
Простые функциональные зависимости. (а) Линия – фотосинтез от солнечной радиации. Точки – экспериментальные данные. (б) Линия – фотосинтез от температуры. Точки – экспериментальные данные. (в) Линии уровня зависимости фотосинтеза от солнечной радиации и дефицита давления водяного пара. (г) Линии уровня зависимости фотосинтеза от солнечной радиации и температуры. На в и г результаты моделирования отображаются только там, где имеются данные. Там, где данные отсутствуют, – белые пятна.
Модель 2: Pt(T)
Результаты моделирования фотосинтеза как функции температуры приводятся на рис. 3б. Погрешность моделирования σcv = 87.75%.
Модель 3: Pqvpd(Q, VPD)
Результаты моделирования фотосинтеза как функции солнечной радиации и дефицита давления водяного пара приводятся на рис. 3в. Погрешность моделирования σcv = 51.28%.
Дефицит давления водяного пара в воздухе является основным внешним фактором, определяющим водный режим растения. Мы не будем продолжать эту ветвь моделирования – водный режим растения будет учтен ниже через устьичную проводимость.
Модель 4: Pqt(Q, T)
Результаты моделирования фотосинтеза как функции солнечной радиации и температуры на рис. 3г. Погрешность моделирования σcv = 51.00%.
Результаты моделирования показывают, что основным фактором, определяющим зависимость фотосинтеза от внешних параметров, является солнечная радиация. Как и следовало ожидать, добавление к основному фактор (Q) других факторов (VPD и T) не сильно уменьшает оценку погрешности. Это объясняется тесной взаимосвязью факторов внешней среды (некоторая синхронность или когерентность).
Для определения направления дальнейшей модификации моделей, проведем для модели 4 анализ остатков как функции времени суток. Для этого остатки, подсчитанные по формуле:
аппроксимируем функцией, зависящей от времени суток. Результаты, приведенные на рис. 4, показывают занижение модельного фотосинтеза в утренние часа и его завышение в вечерние. Это объясняется отсутствием в модели устьичного регулирования газообмена – утром, когда воды в растении достаточно, устьица открыты полностью, а после полудня устьица прикрываются из-за дефицита воды в растении.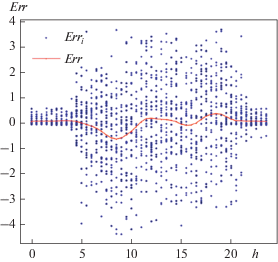
Модель 5: Устьичная проводимость
Воспользуемся устьичной проводимостью, рассчитанной ранее в модели транспирации (Соколов и др., 2019а). Это позволит на основе закона Фика рассчитать концентрацию СО2 в межклетниках Ci:
и представить фотосинтез в виде где функции мультипликативного представления должны удовлетворять условиям монотонностиПоследние три условия представляются оправданными для биохимического процесса фотосинтеза – при увеличении потока энергии (солнечной радиации) и концентрации реагента (СО2) скорость реакции не должна убывать, при отсутствии света фотосинтез не идет (но имеется ночное дыхание, включенное в P). Такие предположения подтверждаются экспериментально, например, световые кривые фотосинтеза демонстрируют насыщение по радиации при постоянстве других условий (Болондинский, Виликайнен, 2014).
Расчеты показывают заметное снижение погрешности σcv = 50.44%.
При анализе найденных функций мультипликативного представления фотосинтеза (рис. 5) следует отметить, что функция Pci(Ci) работает на фотосинтез при Ci в диапазоне 90–250 ppm – ночная концентрация CO2 (около 360 ppm) быстро “выедается” при появлении света.
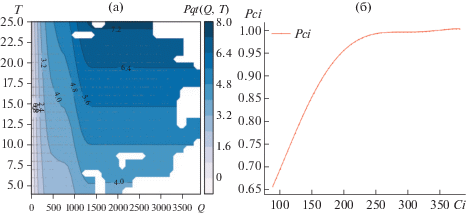
Модель 6: Ассимиляция (брутто фотосинтез) минус дыхание
Разделим (нетто) фотосинтез на две (знакоопределенные) составляющие: ассимиляцию CO2 (брутто фотосинтез) и дыхание:
Обе функции мультипликативного представления ассимиляции предполагаются неубывающими и неотрицательными, при отсутствии света ассимиляция равна 0. Дыхание предполагается периодической, неотрицательной и ограниченной функцией времени суток. Погрешность снизилась σcv = 50.20%. Полученная функция дневной динамики дыхания приведена на рис. 6.
В анализируемый период ночное дыхание у однолетней хвои, в отличие от растущей хвои, имело низкий уровень. Отчасти этому способствовали низкие ночные температуры. При температурах 14–16°С дыхание достигало 0.2 мкмоль м–2 с–1. В утренние часы темновое дыхание также имело невысокие значения, постепенно возрастая к вечеру. Если температура воздуха была в дневное время умеренной (18–20°С), величины дыхания не превышали 0.2–0.3 мкмоль м–2 с–1 Дыхание в дневные часы значительно возрастало, если имела место полуденная депрессия фотосинтеза. Однако, в рассматриваемый период последнее явление практически не наблюдалось.
Модель 7: Изменение CO2 в окружающем воздухе
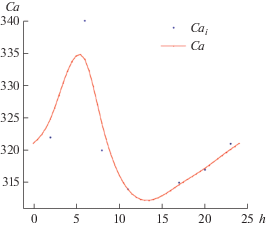
Воспользуемся табл. 2, содержащей набор измерений атмосферного СО2 в разное время суток, для аппроксимации суточного хода атмосферного СО2 периодической функцией (рис. 7).
Рис. 7.
Суточный ход СО2 в атмосфере: точки – измерения, кривая – аппроксимация их периодической функцией времени суток.
Заменим в модели постоянное значение СО2 на найденную функцию времени суток. Результат – погрешность σcv = 50.07%.
Модель 8: Углекислотные компенсации точки
В точке компенсации фотосинтез равен нулю:
т.е. ассимиляция (брутто фотосинтез) равна дыханию. Воспользуемся собранным материалом по УКП (рис. 2) и добавим в задачу идентификации (в целевую функцию) специальный штраф за отклонение ассимиляции от дыхания для всех записей УКП. Таким образом, измерения УКП доопределяют ассимиляцию при малых значениях концентрации CO2. Добавление в задачу дополнительной качественной (наличие точек компенсации) и количественной (значения параметров в точках компенсации) информации приводит к уменьшению погрешности и существенному изменению функции Aci. Теперь, как и следовало ожидать, при минимальных значениях концентрации Сi скорость реакции ассимиляции СО2 близка к нулю (см. рис. 8).Рис. 8.
Ассимиляция как мультипликативная функция (модель 8). Два сомножителя: (а) зависимость от радиации и температуры (линии уровня); (б) зависимость от концентрации СО2 в межклетниках.
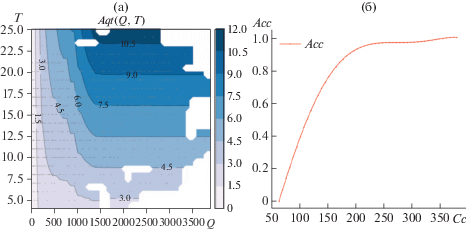
Результат – погрешность σcv = 49.91%.
Модель 9: Мезофильная проводимость
Это последняя надежная модель. Она соответствует потоковой диаграмме, приведенной на рис. 1. Мы шаг за шагом, проверяя различные гипотезы, модифицировали модели. Собранные вместе уравнения в виде большого фрагмента файла задания, содержащего полное описание модели фотосинтеза с учетом мезофильной проводимости приведены в Приложении.
В этой модели появились два новых объекта:
Gm ∊ [0, 0.3] – ограничение на параметр проводимость мезофилла;
P = (Ci – Cc)Gm × 1.0816 – уравнение, формализующее закон Фика для мезофилла.
Как и следовало ожидать, в результате расчетов величина Gm достигла верхнего ограничения 0.3, а погрешность моделирования почти не изменилась – σcv = 49.90%.
Модель 10: Мезофильная проводимость как функция радиации Gm(Q)
Погрешность увеличилась: σcv = 50.14. Это свидетельствует о том, что модель стала слишком сложной для имеющихся данных – слишком много промежуточных искомых функций, не подкрепленных экспериментом. Тем не менее, приводим найденную функцию (рис. 9 а). Полученный результат можно объяснить – при выходе фотосинтеза на насыщение при высоких значениях Q (более 2000) нет необходимости стимулировать поток СO2 путем обеспечения высокой мезофильной проводимости (например, при помощи повышения концентрации и скорости регенерации рибулозобисфосфаткарбоксилаза/оксигеназы).
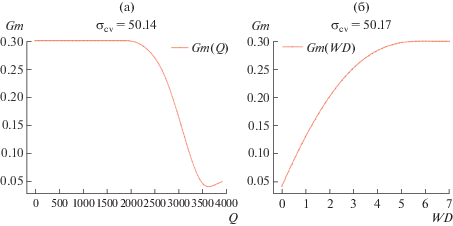
Модель 11: Мезофильная проводимость как функция дефицита воды в растении Gm(WD)
Погрешность (по сравнению с моделью 9) опять увеличилась – σcv = 50.17%. Опять нарушен баланс между сложностью модели и количеством и качеством данных, что приводит к ненадежным результатам – слишком много промежуточных искомых функций, не подкрепленных экспериментом. Тем не менее, приводим график функции (рис. 9 б), имеющий логичную интерпретацию – мезофильная проводимость возрастает при увеличении дефицита влаги в растении, чтобы компенсировать соответствующее уменьшение устьичной проводимости (Цельникер и др., 1993, Stangl et al., 2019).
РЕЗУЛЬТАТЫ МОДЕЛИРОВАНИЯ И ИХ ОБСУЖДЕНИЕ
Формально можно рассматривать проведенное исследование как попытку выбрать лучшую комбинацию гипотез и экспериментальных данных и объединить их в математической модели. Модификация моделей проводилась эволюционным способом от простого к сложному. В целом, критерием такого “эволюционного отбора” является погрешность кроссвалидации – если погрешность модифицированной модели уменьшалась, она “выживала” и использовалась как основа дальнейшей модернизации. Некоторая часть такого отбора была рассмотрена выше, другая (не меньшая по объему), содержащая тупиковые ветви, была опущена. Одна из таких ветвей была связана с мультипликативным представлением:
Сначала это направление успешно эволюционировало, однако, при добавлении в модель УКП проиграло основной ветке в борьбе за минимизацию погрешности.
Анализ (табл. 3) динамики погрешности моделирования данных σcv позволяет выделить ключевые, существенные гипотезы, привлечение которых приводит к заметному уменьшению погрешности: учет устьичной проводимости, учет дыхания и использование точек компенсации.
Таблица 3.
Погрешность моделирования для различных моделей
| № | Название | σcv |
|---|---|---|
| 1 | Pq(Q) | 51.84 |
| 2 | Pt(T) | 87.75 |
| 3 | Pqvpd(Q,VPD) | 51.25 |
| 4 | Pqt(Q,T) | 51.00 |
| 5 | Устьичная проводимость | 50.44 |
| 6 | Ассимиляция минус дыхание | 50.20 |
| 7 | Динамика CO2 в атмосфере | 50.07 |
| 8 | Точки компенсации | 49.91 |
| 9 | Мезофильная проводимость Gm | 49.90 |
| 10 | Мезофильная проводимость Gm(Q) | 50.14 |
| 11 | Мезофильная проводимость Gm(WD) | 50.17 |
Полученные модели позволили рассчитать некоторые среднесуточные показатели, приведенные на рис. 10.
Рис. 10.
Среднесуточные значения: (а) фотосинтеза (P); (б) транспирации (E). (в) Эффективности использования воды (WUE). (г) Дефицит воды в растении. Прерывистые линии – соответствующие средние за период моделирования значения: aP = 2.52, aE = 0.196, aWUE = 12.8, aWD = 2.32.
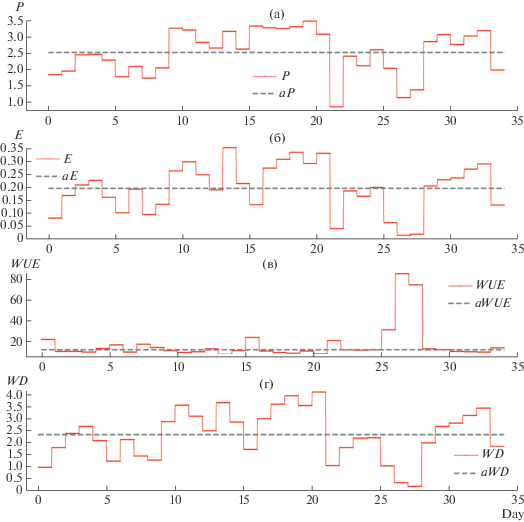
Анализ среднесуточных значений фотосинтеза и транспирации (рис. 10а, 10б) показывает значительную синхронность их изменений в течение первых 25 дней анализируемого периода. В результате и величины WUE (рис. 10в) сохраняли определенное постоянство и имели всего два небольших выброса – на 16 и 22 день (5 и 11 июля). Как в прохладные пасмурные дни начала периода, так и далее на протяжении нескольких солнечных дней с 28 июня по 2 июля среднеднесуточные транспирационные расходы составляли 0.15 ± 0.06 ммоль м–2 с–1, что несколько ниже обычных значений для начала июля. В какой-то мере это было обусловлено невысокой температурой воздуха (18–20°С) и VPD, не превышающего 1 кПа.
В анализируемый период наблюдался радиальный рост дерева, а также рост молодой хвои. Наличие мощных аттрагирующих центров обуславливала большую потребность в ассимилятах, результатом чего был высокий среднесуточный уровень фотосинтеза (до 3.5 мкмоль м–2 с–1). Учитывая, что устьичная проводимость была ниже среднего уровня и составляла в первую половину дня 0.04–0.06 моль м–2 с–1, фотосинтез не снижался благодаря высокой мезофильной проводимости. Расчеты показали (Соколов и др., 2019а), что днем наблюдалась дегидратация ствола, запасы влаги в котором уменьшались. Обезвоживание дерева в солнечные дни постепенно нарастало (рис. 10г). Далее, в ходе кратковременного потепления на 14-й день (до 25°С) и увеличения VPD до 2 кПа наблюдался максимальный за все время уровень транспирации (1.2 ммоль м–2 с–1), который был обусловлен большим VPD. При этом устьичная проводимость снизилась на 10–15%. В целом ряде исследований последних десятилетий установлена обратная связь устьичной проводимости как с VPD, так и с WD (Warren, 2008; Campany et al., 2016; Stangl et al., 2019). Это подтверждают и наши предыдущие исследования (Соколов и др., 2019а)
В следующие дни дерево использовало низкий уровень VPD в утренние часы для максимального раскрытия устьиц. При этом диффузия CO2 через устьица была значительной при минимальных потерях влаги. Это отразилось на 16-й день в величине WUE, возросшей на 80%.
А прохладную, преимущественно пасмурную погоду начала исследуемого периода дефицит воды в дереве (WD), был в целом ниже среднего уровня (1–2 л). В последующий солнечный период с более высокими температурами воздуха WD возрос до 4 л. Слабое снижение величин WUE было обусловлено последовательно возрастающим WPD, приводящим к небольшому росту транспирации, несмотря на уменьшение устьичной проводимости. Далее после нескольких дней с дождями водный баланс дерева пришел в норму (WD < 0.3 л) и величины WUE сохраняли относительное постоянство около среднего уровня.
Нужно отметить, что дегидратация ствола на протяжении всего июля оказывала значительное влияние на устьичную проводимость, что обусловило, как правило, ее невысокие значения на протяжении дня. Лишь в дождливые периоды при минимальных значениях VPD устьичная проводимости была высокой, что обеспечивало относительно высокий уровень поглощения CO2 даже при низкой инсоляции. Именно этим объясняются очень большие выбросы WUE на 26–28-й день – до 85 мкмоль CO2 (ммоль Н2О)–1.
Используя средние за период моделирования среднесуточные значения фотосинтеза – 2.52 мкмоль м–2 с–1, а также транспирации – 0.196 ммоль м–2 с–1 можно оценить величины поглощения CO2 и испарения Н2О на 1 га древостоя сосняка черничного.
Измерения, проведенные на модельном дереве 45–55-летнего возраста (высота 14.7 м, диаметр 16 см, протяженность кроны 8 м) показали, что площадь всей хвои составила 45 ± 5 м2. При этом площадь световой хвои и хвои 3-го года и выше составляла 33% и 35% соответственно. Используя скорости транспирации (данные Б.М. Веселкова), связанные с освещенностью и возрастом, можно привести площадь всей хвои кроны к площади хвои одного типа, например, освещенной 1–2-летнего возраста. То есть, если бы вся крона дерева состояла из освещенной хвои 1–2-летнего возраста, то именно данная площадь обеспечила бы такую скорость транспирационного расхода влаги, как и реальное наше дерево. Проведенный по нескольким методикам расчет (Кайбияйнен, 1990) показал, что приведенная площадь хвои всей кроны к площади 1–2-летней световой хвои имела величину 32 ± 5 м2. Все эти количественные расчеты не очень точны, есть ошибки в интегрировании расхода влаги побегами на всю крону.
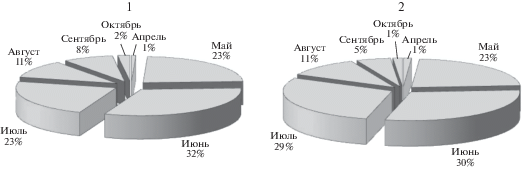
Для пересчета на гектар воспользуемся данными измерений параметров модельных деревьев, полученных при изучении таксационных характеристик нашего древостоя (Казмиров и др., 1977). В сосняке черничном свежем вся хвоя составляет 4% от общей фитомассы всех деревьев – 47.6 ц/га. С учетом соотношений, полученных для нашего модельного дерева, поглощать углекислоту как 1–2-летняя хвоя будут примерно 33.3 ц/га. Достаточно простой расчет показал, что за выбранный период 1 гектар леса поглотил 14.6 т, при этом расход воды составил 464 т. Используя наши данные о распределении месячных сумм СО2-газообмена и транспирации побегов сосны (рис. 11), можно для гектара леса за вегетацию дать приблизительную оценку поглощения CO2 – 34 т, и испарения Н2О – 1351 т. Погрешность этих оценок на уровне 20–25%.
Рис. 11.
Распределение месячных сумм транспирации (1) и фотосинтеза (2) побегов в верхней части кроны экспериментальной сосны.
За месяцы вегетации выпало 92 мм осадков (Кайбияйнен, 1990), на 1 га – 920 м3 воды. По нашим расчетам гектар леса испарил несколько больше. Но нужно учесть, что к моменту начала вегетации в почве содержались значительные запасы талых вод – около 1000 тонн/га. Таким образом, величина испарения с 1 га в целом соответствует литературным данным.
ЗАКЛЮЧЕНИЕ
Объединение знаний о процессах фиксации СО2 растением, (раздел “Роль внешних факторов среды и внутренних параметров в моделях фотосинтеза на уровне листа”), с экспериментальными данными (раздел “Объект исследования и экспериментальные данные”) на основе методов математического моделирования (раздел “Применение технологии сбалансированной идентификации для анализа моделей фотосинтеза”) позволило шаг за шагом модифицировать (усложнять) описание объекта и в результате выбрать модель, сложность которой соответствует количеству и качеству экспериментального материала. Использование УКП позволило оценить динамику концентрацию CO2 в хлоропластах и зависимость от нее ассимиляции СО2. Однако надежно рассчитать проводимость мезофилла, как функции внешних условий не удалось. Возможно, проведение дополнительных активных экспериментов с целью получения УКП в более широком диапазоне переменных и привлечение дополнительных знаний о дневном дыхании (фотодыхании) позволит продвинуться дальше.
Еще одно направление развития – это объединение моделей транспирации и фотосинтеза. Мы использовали модель транспирации как источник данных и не рассматривали обратную связь: влияние ключевых параметров фотосинтеза, прежде всего концентраций CO2 в разных компартментах хвои на транспирацию растения, на работу устьиц.
Работа была выполнена с использованием оборудования центра коллективного пользования “Комплекс моделирования и обработки данных исследовательских установок мега-класса” НИЦ “Курчатовский институт”, http://ckp.nrcki.ru/.
В расчетах использовался сервис решения наборов независимых задач оптимизации https://optmod. distcomp.org/apps/vladimirv/solve-set-opt-probs. Это один из нескольких сервисов оптимизационного моделирования https://optmod.distcomp.org на платформе Эверест, http://everest.distcomp.org/ (Sukhoroslov O. et al., 2015).
Работа выполнена сотрудниками ИППИ РАН при финансовой поддержке РНФ проект 16-11-10352 (разд. “Применение технологии сбалансированной идентификации для анализа моделей фотосинтеза” и “Результаты моделирования и их обсуждение”). Финансовое обеспечение исследований сотрудниками Института леса КарНЦ РАН осуществлялось из средств федерального бюджета на выполнение государственного задания КарНЦ РАН (0220-2014-0010) (разд. “Роль внешних факторов среды и внутренних параметров в моделях фотосинтеза на уровне листа” и “Объект исследования и экспериментальные данные”).
ПРИЛОЖЕНИЕ: ФРАГМЕНТ ФАЙЛА ЗАДАНИЯ ДЛЯ МОДЕЛИ МЕЗОФИЛЬНОЙ ПРОВОДИМОСТИ
Ключевое слово “Var:” означает неизвестную функцию-переменную, которую необходимо идентифицировать (определить), “Param:” – известную (заданную) функцию, “EQ:” – уравнение. Знак “#” означает комментарий
Var: P(t) # фотосинтез
Param: Q(t) # радиация
T(t) # температура
h(t) # время суток
Param: Ca(h) # концентрация CO2 в атмосфере от времени суток h
Var: Ci(t ≥ 64 # концентрация CO2 в межклетниках
Cc(t) ≥ 64 # концентрация CO2 в хлоропластах
A(t) ≥ 0 # ассимиляция CO2
Aqt(Q, T) ≥ 0 # зависимость ассимиляции от Q и T
Acc(Cc) ≥ 0 # зависимость ассимиляции от Cc
R(h) ∈ [0, 0.2]; R(0) = R(24) # дыхание от времени суток
Param: Gs(t) # устьичная проводимость
Var: Gm ∊ [0, 0.3] # проводимость мезофилла
EQ: P = (Ca(h) – Ci) Gs × 1.0816 # закон Фика для устьичной проводимости
P = (Ci – Cc)Gm × 1.0816 # закон Фика для мезофилла
P = A – R (h) # фотосинтез = ассимиляция – дыхание
A = Aqt(Q, T)Acc(Cc) # ассимиляция
Aqt(0, T) = 0 # при Q = 0 ассимиляция не идет
Acc (360) = 1 # нормировка
d/dgCc(Acc(Cc)) ≥ 0 # Acc не убывает при росте концентрации Cc
d/dgQ(Aqt(Q, T)) ≥ 0 # Aqt не убывает при росте концентрации Q
Список литературы
Барри Д. А., Даунтон У.Д.С. (1987) Зависимость фотосинтеза от факторов окружающей среды. Фотосинтез. Т.2 (ред. Говинджи). М.: Мир, 273-364.
Болондинский В.К., Виликайнен Л.М. (2014) Исследование световой зависимости фотосинтеза у карельской березы и березы повислой в условиях разной обеспеченности элементами минерального питания. Труды Карельского научного центра РАН. Серия Экспериментальная биология. (5), 207-213.
Болондинский В.К., Кайбияйнен Л.К. (2003) Динамика фотосинтеза в сосновых древостоях Физиология растений 50(1), 105-114.
Веселков Б.М., Тихов П.В. (1984) Связь транспорта воды по ксилеме с интенсивностью транспирации у сосны обыкновенной. Физиология растений (31), 1099-1107.
Голомазова Г.М. (1987) Влияние внешних факторов на фотосинтез хвойных. Красноярск: Изд-во Краснояр. ун-та, 120 с.
Казимиров Н.И., Волков А.Д., Зябченко С.С., Иванчиков А.А., Морозова Р.М. (1977) Обмен веществ и энергии в сосновых лесах Европейского Севера. Л.: Наука, 304 с.
Кайбияйнен Л.К. (1990) Экофизиология водного режима сосны и сосновых древостоев: Автореф. дис. … докт. биол. наук. М.: ИЭМЭЖ им. Северцова АН СССР, 45 с.
Лайск А.Х. (1977) Кинетика фотосинтеза и фотодыхания Сз-растений. М.:Наука, 195с.
Молчанов А.Г. (1983) Экофизиологическое изучение продуктивности древостоев. М.: Наука, 135 с.
Оя В.М., Расулов Б.Х. (1981) Двухканальная газометрическая аппаратура для исследования фотосинтеза листа в полевых условиях. Физиология растений (28), 887-895.
Соколов А.В., Болондинский В.К., Волошинов В.В. (2019а) Технология сбалансированной идентификации для выбора математической модели транспирации сосны. Математическая биология и биоинформатика 14(2), 665-682.
Соколов А.В., Мамкин В.В., Авилов В.К., Тарасов Д.Л., Курбатова Ю.А., Ольчев А.В. (2019б) Применение метода сбалансированной идентификации для заполнения пропусков в рядах наблюдений за потоками СО2 на сфагновом верховом болоте. Компьютерные исследования и моделирование 1(11), 153-171.
Соколов А.В., Волошинов В.В. (2018) Выбор математической модели: баланс между сложностью и близостью к измерениям. International J. Open Information Technologies 6(9), 33-41.
Тимохина А.В., Прокушкин А.С., Панов А.В. (2014) Суточная и сезонная динамика концентрации СО2 и СН4 в атмосфере над экосистемами Западной Сибири (Приенисейская часть). Вестник КрасГАУ (12), 83-88.
Цельникер Ю.Л. (1978) Физиологические основы теневыносливости древесных растений. Л.: Наука, 215 с.
Цельникер Ю.Л., Малкина И.С., Ковалев А.Г., Чмора С.Н., Мамаев В.В., Молчанов А.Г. (1993) Рост и газообмен СО2 у лесных деревьев. М.: Наука, 256 с.
Щербатюк А.С., Русакова Л.В., Суворова Г Г., Янькова Л.С. (1991) Углекислый газообмен хвойных Предбалькалья. Новосибирск: Наука, 134 с.
Campany C.E., Tjoelker M.G., Caemmerer von S., Duursma R.A. (2016) Coupled response of stomatal and mesophyll conductance to light enhances photosynthesis of shade leaves under sunflecks. Plant, Cell Environ 39(12), 2762-2773.
Eichelmann H., Laisk A. (1999) Ribulose-1,5-bisphosphate carboxylase/oxygenase content, assimilatory charge, and mesophyll conductance in leaves. Plant Physiology (119), 179-189.
Eichelmann H., Oja V., Rasulov B., Padu E., Bichele I., Pettai H., Niinemets Ü., Laisk A. (2004) Development of leaf photosynthetic parameters in Betula pendula Roth leaves: correlations with photosystem I density. Plant Biology (6), 307-318.
Flexas J., Diaz-Espejo A., Galmés J., Kaldenhoff R., Medrano H., Ribas-Carbo M. (2007) Rapid variations of mesophyll conductance in response to changes in CO2 concentration around leaves. Plant, Cell Environ 30(10), 1284-1298.
Flexas J., Galmés ., Gallé A., Gulias J., Pou A., Ribas-Carbo M., Tomas M., Medrano H. (2010) Improving water use efficiency in grapevines: potential physiological targets for biotechnological improvement. Aust J Grape Wine Res 16(s1), 106-121.
Flexas J., Ribas-Carbo M., Diaz-Espejo A., Galmés J., Medrano H. (2008) Mesophyll conductance to CO2: current knowledge and future prospects. Plant, Cell Environ 31(5), 602-621.
Grace J. (2000) The Carbon Cycle // The Encyclopedia of Biodiversity. 1, 609-628.
Hu J., Moore D.J.P., Riveros-Iregui D.A., Burns S.P., Monson R.K. (2010) Modeling whole-tree carbon assimilation rate using observed transpiration rates and needle sugar carbon isotope ratios. New Phytol 185(4), 1000-1015.
IPCC (2013) Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the IPCC. – Cambridge: Cambridge University Press, 1535 p.
Jarvis P.G. (1971) The estimation of resistances to carbon dioxide transfer. Plant photosynthetic production manual of methods (ed. Sestak Z., Catsky J., Jarvis P.G.), 566-631.
Klein T., Rotenberg E., Tatarinov F., Yakir D. (2015) Association between sap flow-derived and eddy covariance-derived measurements of forest canopy CO2 uptake. New Phytol 209(1), 436-446.
Laisk A., Loreto F. (1996) Determining photosynthetic parameters from leaf CO2 exchange and chlorophyll fluorescence. Ribulose-1,5-bisphosphate carboxylase/oxygenase specificity factor, dark respiration in the light, excitation distribution between photosystems, alternative electron transport rate, and mesophyll diffusion resistance. Plant Physiology (110), 903-912.
Laisk A., Oja V. (1998) Dynamics of leaf photosynthesis. Rapid-response measurements and their interpretations. Techniques in Plant Sciences (1), 1-160.
Lloyd J., Shibistova O., Zolotoukhine D. (2002) Seasonal and annual variations in the photosynthetic productivity and carbon balance of a central Siberian pine forest. Tellus (54 B), 590-610.
Loreto F., Tsonev T., Centritto M. (2009) The impact of blue light on leaf mesophyll conductance. J Exp Bot 60(8), 2283-2290.
Moldau H., Kull O. (1993) Differential susceptibility of mesophyll CO2 exchange to ozone in soil- or sand-grown Phaseolus vulgaris L. plants. Photosynthetica (28), 37-44.
Perttu K., Bischof W., Grip H., Jansson P.-E., Lindgren Å., Lindroth A., Norén B. (1980) Micrometeorology and hydrology of pine forest ecosystems. I. Field studies. Structure and Function of Northern Coniferous Forests. Ecol. Bull. (Stockholm) (32), 75-121.
Peters G.P., Marland G., Quere Le (2012) Rapid growth in CO2 emissions after the 2008–2009 global financial crisis. Nature Climate Change (2), 2-4.
Sukhoroslov O., Volkov S., Afanasiev A. (2015) A Web-Based Platform for Publication and Distributed Execution of Computing Applications // 14th International Symposium on Parallel and Distributed Computing (ISPDC). IEEE, pp. 175-184.
Sokolov A.V., Voloshinov V.V. (2019) Balanced Identification as an Intersection of Optimization and Distributed Computing, https://arxiv.org/pdf/1907.13444.pdf
Stangl Z.R., Tarvainen L., Wallin G., Ubierna N., Räntfors M., Marshall J.D. (2019) Diurnal variation in mesophyll conductance and its influence on modelled water use efficiency in a mature boreal Pinus sylvestris stand. Photosynth Res. (141), 53-63. https://doi.org/10.1007/s11120-019-00642-9
Ubierna N., Marshall J.D. (2011) Estimation of canopy average mesophyll conductance using δ13C of phloem contents. Plant, Cell Environ 34(9), 1521-1535.
Warren C.R. (2008) Does growth temperature affect the temperature responses of photosynthesis and internal conductance to CO2? A test with Eucalyptus regnans. Tree Physiol 28(1), 11-19.
Дополнительные материалы отсутствуют.