

Геоморфология и палеогеография, 2020, № 1, стр. 53-67
НОВЫЕ ДАННЫЕ О ПОЗДНЕЧЕТВЕРТИЧНОМ СКЛАДКООБРАЗОВАНИИ И ДЕФОРМАЦИИ РЕЛЬЕФА НА СЕВЕРЕ ТАМАНСКОГО П-ОВА, КРАСНОДАРСКИЙ КРАЙ
О. В. Гайдаленок 1, *, С. А. Соколов 1, Я. А. Измайлов 2, П. Д. Фролов 1, 3, В. В. Титов 4, А. С. Тесаков 1, В. Г. Трифонов 1, А. В. Латышев 5, Н. А. Орлов 1
1 Геологический институт РАН
Москва, Россия
2 Сочинское отделение РГО
Сочи, Россия
3 Лаборатория макроэкологии и биогеографии беспозвоночных СПбГУ
Санкт-Петербург, Россия
4 Южный научный центр РАН
Ростов-на-Дону, Россия
5 Институт физики Земли имени О.Ю. Шмидта РАН
Москва, Россия
* E-mail: ogaydalen@yandex.ru
Поступила в редакцию 11.07.2019
После доработки 06.09.2019
Принята к публикации 08.10.2019
Аннотация
Исследованы фрагменты брахиантиклинальной складки на юго-западном берегу Динского залива Таманского п-ова. Уточнен древнеэвксинский (конец раннего – начало среднего неоплейстоцена) возраст как северного крыла антиклинали, представленного изгибом слоев до 40–42°, так и юго-восточного, где слои выполаживаются от 15–16° до почти горизонтального залегания. Впервые проведено палеомагнитное опробование разреза юго-восточного крыла, а в его основании найден рог оленя, датируемый ранним плейстоценом (гелазий–начало калабрия). Выявленную складчатую деформацию в сглаженном виде повторяет установленный геоморфологическими методами изгиб поверхности свода, сложенный делювиальными суглинками и супесями верхов среднего и/или верхнего неоплейстоцена. На основе описанной и подобных ей деформаций в других частях Таманского п-ова делается вывод об интенсивных складкообразовательных движениях на полуострове во второй половине неоплейстоцена.
ВВЕДЕНИЕ
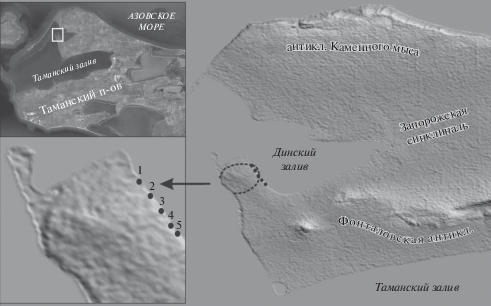
Представлены новые данные о строении и средне-позднечетвертичном возрасте сравнительно небольшой антиклинальной брахискладки (Динского купола), исследованной авторами в ходе полевых работ 2018 и 2019 гг. Определение возраста складки является существенным вкладом в обоснование молодого возраста складчатости Таманского полуострова. Брахиантиклиналь расположена к северу от западного периклинального замыкания крупной Фонталовской антиклинали (рис. 1). Слагающие брахиантиклиналь отложения вскрыты в береговых обрывах на юго-западном берегу Динского залива Таманского п-ова. Наличие среди них древнеэвксинских отложений (конец нижнего – средний неоплейстоцен) впервые отметил Н.С. Благоволин [1], а присутствие как древнеэвксинских, так и чаудинских (нижний неоплейстоцен) отложений констатировал Г.И. Попов [2]. Он же охарактеризовал заключенную в тех и других фауну моллюсков. Полученные нами новые данные уточняют строение и возраст антиклинали.
Обсуждаемые в статье стратиграфические подразделения российской черноморско-азовской шкалы соотносятся с международной шкалой [3, 4] следующим образом. Понт примерно соответствует мессинию (~6.4–5.3 млн лет), палеоплейстоцен – гелазию, эоплейстоцен – калабрию, а чаудинская трансгрессия, близкая по возрасту к бакинской трансгрессии Каспийского моря, охватывала временной интервал изотопно-кислородных стадий от MIS 19 до MIS 13 (0.78–0.47 млн лет). Возраст древнеэвксинской трансгрессии дискуссионен. Обосновывался вариант, согласно которому она представляла собой позднюю стадию развития чаудинской трансгрессии и, возможно, приходилась на MIS 15 или MIS 13 [5]. Согласно другому варианту, древнеэвскинская трансгрессия соответствует стадии MIS 11 [6, 7]. Судя по соотношениям в разрезах, древнеэвксинская трансгрессия близка по возрасту к чаудинской, но, вероятно, отделена от нее стратиграфическим перерывом. Если допустить, что чаудинская трансгрессия закончилась в конце стадии MIS 15, можно сопоставить древнеэвксинскую трансгрессию с MIS 13. Однако, исходя из палеонтологических данных и палеогеографических реконструкций, более вероятным представляется ее сопоставление с теплой эпохой MIS 11, т.е. лихвинским межледниковьем Русской равнины (~0.4 млн лет). Возраст карангатской трансгрессии определен в интервале 0.13–0.08 млн лет – начало позднего неоплейстоцена, стадия MIS 5 с максимумом в 5e, примерно совпадающая с микулинским межледниковьем Русской равнины [6, 8].
МАТЕРИАЛЫ И МЕТОДИКА
При подготовке к полевым работам использовались геологические карты м-ба 1 : 200 000 [9, 10], карта четвертичных образований м-ба 1 : 200 000 [11], цифровая модель рельефа [12], снимки Google Earth и данные GPS-навигации. В ходе полевых работ детально описаны и опробованы береговые разрезы и определено строение свода Динской антиклинали. Для уточнения возраста отложений, участвующих в ее строении, определен видовой состав моллюсков и идентифицированы найденные фрагмент рога оленя и зуб мелкого млекопитающего. При определении ископаемых моллюсков использованы работы [13–17]. Впервые проведено палеомагнитное опробование отложений юго-восточного крыла антиклинали, на основании чего делается вывод о палеомагнитной эпохе накопления отложений, формирующих разрез.
ПОЛУЧЕННЫЕ РЕЗУЛЬТАТЫ
Проявление антиклинали в рельефе. Сводовая часть Динской складки является частью довольно плоской террасовидной поверхности. Как видно в обнажениях на берегу залива (в прибрежном сечении), эта поверхность полого понижается к северу и ЮЮВ, очерчивая брахиантиклиналь. Вместе с тем плоская сводовая часть протягивается от берега Динского залива на ЗЮЗ, слабо повышаясь. Поверхность повсеместно сложена покровными суглинками и супесями и вблизи берега моря прорезана небольшими оврагами. Судя по плановым очертаниям этой поверхности, складка выражена в рельефе пологим брахиантиклинальным сводом, слабо удлиненным в ВСВ направлении (см. рис. 1). В береговом сечении сводовая часть антиклинали плоская и протяженная, тогда как участки существенного наклона поверхности узки по сравнению со сводом. Это придает складке слабо выраженную коробчатую форму.
Геологическое строение антиклинали. Покровные суглинки и супеси, слагающие поверхность антиклинали, содержат линзы и включения более грубообломочного материала и несут следы водной переработки. По аналогии с другими подобными образованиями Таманского п-ова их можно предположительно датировать концом среднего плейстоцена – поздним плейстоценом [18, 19]. Прорезающие этот покров голоценовые (?) овраги отмечают верхний предел возраста покровного комплекса. Мощность покрова, обычно не превышающая на своде 1 м, возрастает до 1.5–2.5 м на крыльях брахиантиклинали [1] (рис. 2, б), что свидетельствует о росте складки как во время накопления покрова, так и позднее, когда поверхность была уже сформирована.
Рис. 2.
(а) – Изгиб террасовидной поверхности, очерчивающий антиклинальную складку. Показаны обнажения, приуроченные к этой складке на юго-западном побережье Динского залива (1 – 45°22'5.13''С, 36°47'12.90''В; 2 – 45°21'57.13''С, 36°47'18.19''В; 3 – 45°21'51.69''С, 36°47'22.65''В; 4 – 45°21'44.66''С, 36°47'27.08''В; 5 – 45°21'41.19''С, 36°47'29.97''В). (б) – Схематический геологический разрез Динской антиклинали: 1 – покровные суглинки, супеси и современная почва, 2 – древнеэвксинские пески (QIIde), 3 – чаудинские пески и алевриты (QIIch), 4 – эоплейстоценовые суглинки (QE), 5 – глины и пески понта (N1p) с глыбами мэотических известняков (N1m), 6 – овраги, 7 – конусы выноса, 8 – геологические границы (a– достоверные, б – предполагаемые), 9 – фаунистические находки (a – моллюски, б – крупные млекопитающие, в – мелкие млекопитающие). (в) – Геологическое строение отдельных обнажений: 1 – глины, 2 – песок, 3 – алеврит, 4 – покровный комплекс, 5 – фаунистические находки (а – моллюски, б – крупные млекопитающие, в – мелкие млекопитающие), 6 – почвенно-растительный слой, 7 – палеомагнитные пробы.
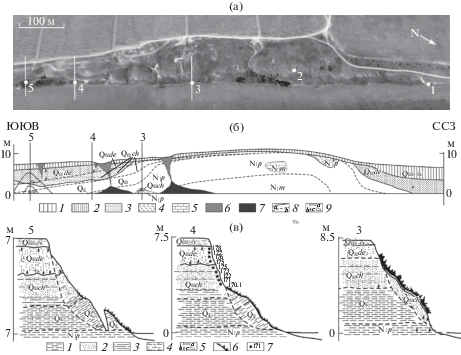
Подстилающие антиклиналь отложения изучены в береговом уступе Динского залива. Ее северное крыло, обнажающееся в уступе высотой до 8 м (1 на рис. 1 и рис. 2), представлено резким флексуроподобным изгибом слоев (рис. 3). В строении крыла складки принимают участие следующие отложения (снизу вверх):
Рис. 3.
Наклон верхнепонтических (а) и древнеэвксинских (б) отложений на юго-западном берегу Динского залива (1 на рис. 1 и рис. 2, а). Сплошной линией показана граница геологических отложений, пунктирной линией – маркирующий горизонт внутри одного возрастного диапазона. (в) – стратиграфическая колонка обнажения в точке наблюдения 1 (усл. знаки см. рис. 2, в).

1. Глины мелкооскольчатые, серо-коричневые, жирные, видимой мощностью до 1 м.
2. Пески тонкозернистые, серые, с тонкой параллельной слоистостью и пропластками среднезернистого песка; 0.2 м.
3. Глины серо-коричневые, слоистые, с пропластками среднезернистых ожелезненных песков и прослоем (0.1 м) тонкозернистого слабосцементированного ожелезненного песчаника в кровле; 1.8–1.9 м. Обильная верхнепонтическая малакофауна с характерными двустворчатыми моллюсками Paradacna abichi и Dreissenа anisoconcha.
4. Пески тонкозернистые, литологически однородные, желтоватого цвета, с параллельной слоистостью и ожелезнением; 6 м. Обильная малакофауна древнеэвксинского облика с типичными Corbicula fluminalis и Pyrgula dimidiata (рис. 4). Нижние горизонты песков, не содержащие фауну, возможно, имеют чаудинский возраст [2].
Рис. 4.
Фототаблица фауны моллюсков. Масштабная линейка равна 5 мм. 1–3 – Dreissena anisoconcha Andrusov, 1897; слой 1 (2 на рис. 1 и рис. 2, а), верхний понт: 1 – правая створка изнутри, 2 – правая створка снаружи, 3 – левая створка снаружи; 4 – Paradacna abichi (R. Hörnes, 1874); слой 1 (2 на рис. 1 и рис. 2, а), верхний понт: правая створка снаружи; 5–6 – Plagiodacna modiolaris (Rousseau, 1842); слой 2 (2 на рис. 1 и рис. 2, а) киммерий (переотложение): 5 – левая створка снаружи, 6 – правая створка снаружи; 7–8 – Prosodacna semisulcata (Rousseau, 1842); слой 2 (2 на рис. 1 и рис. 2, а), киммерий (переотложение): 7 – левая створка снаружи, 8 – правая створка снаружи; 9–12 – Didacna tamanica Nevesskaya, 1963, слой 2 (2 на рис. 1 и рис. 2, а), чауда: 9 – левая створка изнутри, 10 – левая створка снаружи, 11 – правая створка изнутри, 12 – правая створка снаружи; 13–16 – Didacna cf. rudis subvulgaris Nevesskaya, 1963; слой 2 (2 на рис. 1 и рис. 2, а), чауда: 13 – левая створка изнутри, 14 – левая створка снаружи, 15 – правая створка изнутри, 16 – правая створка снаружи; 17–20 – Didacna pontocaspia (Pavlov, 1925); верхний прослой слоя 3 (3 на рис. 1 рис. 2, а), древний эвксин: 17 – левая створка изнутри, 18 – левая створка снаружи, 19 – правая створка изнутри, 20 – правая створка снаружи; 21–24 – Corbicula fluminalis (O.F. Müller, 1774); слой 4 (1 на рис. 1 и рис. 2, а), древний эвксин: 21 – левая створка изнутри, 22 – левая створка снаружи, 23 – правая створка изнутри, 24 – правая створка снаружи.

5. Суглинки и супеси светло-коричневые с белесыми пропластами в нижней части, вверху переходящие в современный почвенный слой; до 2.5 м.
Слои 1–3 и пачка 4 вскрыты в двух соседних обнажениях, разделенных задернованным участком, так что контакт между ними не виден. Наиболее крутые углы падения на крыле складки приходятся на выходы пачки 4, где наклоны составляют 40–42° в направлении 345°. Северо-западнее, в обнажении со слоями 1–3, они выполаживаются до 32° по азимуту 20°, а далее до 4° по азимуту 335.
В 460 м юго-восточнее (3 на рис. 1 и рис. 2) вскрыт фрагмент другого крыла брахискладки с углами падения слоев 12–15° по азимуту 95°. Здесь непосредственно на берегу залива снизу вверх обнажаются:
1. Глины серые с ожелезненными желваками; видимая мощность 0.2 м. Малакофауна верхнепонтического облика с характерными Paradacna abichi и Dreissenа anisoconcha.
2. Пески светло-бежевые; 0.6 м. Содержат ископаемые моллюски чаудинского облика, среди которых Didacna tamanica, D. cf. rudis subvulgaris, а также Dreissena sp. и Viviparus sp., наряду с которыми присутствуют переотложенные из киммерия моллюски: Plagiodacna modiolaris и Prosodacna semisulcata (см. рис. 4).
Чуть севернее и выше по склону обнаружены небольшие выходы песков с ожелезнением и волнистой слоистостью, падающих по азимуту 135° под углом до 16° и, возможно, принадлежащих древнеэвксинским слоям.
Еще в 220 м к юго-востоку (4 и 5 на рис. 1 и рис. 2) слои выполаживаются, и в береговом уступе высотой 7–8 м обнажается субгоризонтально залегающая толща песчаного состава с глинистым слоем в основании. Разрез толщи снизу вверх таков:
1. Глины видимой мощностью до 2 м. Найден ископаемый сброшенный левый рог оленя Cervus cf. nestii (5 на рис. 2; рис. 5, а, б). Вершина рога разрушена. Основание рога повреждено, что не позволяет точно определить высоту первого разветвления, а венчик имеет следы небольшой окатанности. Высота отхождения первого отростка больше, чем переднезадний диаметр основания рога (таблица). Первый отросток округлый в сечении, не разветвляющийся, в дистальной половине плавно загибающийся наружу. Штанга отклоняется назад и наружу; дистально незначительно расширяется, вероятно, образуя разветвление в сагиттальной плоскости. Угол отхождения первого отростка от штанги – около 70°. На штанге отсутствуют дополнительные отростки. Основание рога и штанга субтреугольные в сечении, уплощены в латерально-медиальном направлении. Индекс уплощенности основания рога – 0.9, штанги – 0.8.
Рис. 5.
Левый рог Cervus cf. nestii (Azzaroli, 1948). (a) – медиальная сторона, (б) – вид спереди, нижний плейстоцен. Правый первый верхний коренной Arvicola mosbachensis Schmidtgen, 1911, (в) – жевательная поверхность, средний плейстоцен. Местонахождение Динское.
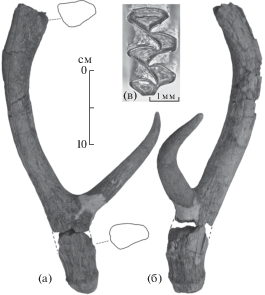
Размеры рога, наличие одиночного латерально изогнутого первого отростка, расположенного относительно высоко над венчиком, отсутствие дополнительных отростков на уплощенной штанге позволяют отнести находку к некрупным раннеплейстоценовым оленям, сходным с Cervus nestii (Azzaroli, 1947). Стратиграфическое распространение C. nestii на территории западной Европы определяется в рамках начала – середины раннего плейстоцена (средний виллафранк – начало позднего виллафранка; поздний плиоцен – ранний плейстоцен Италии) [20]. Сходная форма оленя с немного более примитивной морфологией зубов Cervus abesalomi, Kahlke, 2001, описана из местонахождения Дманиси (Грузия) [20]. В общем, интервал существования оленя из слоя 1 совпадает со временем обитания на юге Восточной Европы хапровского и псекупского фаунистических комплексов. Однако до сих пор данная форма оленя с территории Приазовья и Предкавказья не указывалась. Помимо крупных представителей Cervidae – Eucladoceros и Arvernoceros для этих ассоциаций указывался мелкий олень Cervus (Rusa) philisi (=Metacervocerus rhenanus, Dubois, 1904) [21, 22]. Описываемая находка из слоя 1 отличается от рогов M. rhenanus относительно большими размерами и большей уплощенностью основания рога и штанги. Находки остатков оленей сходного размерного класса известны из ряда местонахождений таманского териокомплекса в Приазовье. Однако их малочисленность и фрагментарность не позволяют определить таксономическую принадлежность. Таким образом, недостаточность материала из слоя 1 не позволяет на данный момент провести более точное определение находки, а имеющиеся данные определяют возраст вмещающих отложений нижним плейстоценом европейской шкалы. Вероятный более узкий стратиграфический интервал – палеоплейстоцен – начало эоплейстоцена (гелазий – начало калабрия). Сходные отложения описаны ниже чаудинских и древнеэвксинских слоев в близлежащем более южном разрезе урочища Малый Кут [23, 24].
2. Пески тонкозернистые желтовато-серые с элементами косой слоистости и тонкими прослоями глин, а также прослоем (0.5 м) алеврита в основании; 2.5 м. По трещинам отмечены ожелезнение и редкие примазки карбонатов. 5 отобранных палеомагнитных образцов показали нормальную полярность (Брюнес). На основе намагниченности, сопоставления с соседними разрезами и древнеэвксинского возраста вышележащих песков слой 2 определен как чаудинский.
3. Пески тонкозернистые бежевые, выше преимущественно серые, с параллельной или косой слоистостью, в нижней части ожелезненные; 3–3.5 м. Отделены от слоя 2 эрозионным контактом. В нижней (слой 3.1) и верхней (слой 3.3.) частях пачки содержится обильная морская древнеэвксинская малакофауна, включая Didacna pontocaspia и Corbicula fluminalis (см. рис. 4), а также единичный остаток водяной полевки Arvicola mosbachensis (5 на рис. 2; рис. 5, в) из базального горизонта слоя. Изолированный правый первый верхний коренной зуб некорнезубой цементной полевки по морфологии и размерам определен как Arvicola mosbachensis Schmidtgen, 1911 (=A. cantianus (Hinton, 1910)). Длина, ширина и высота зуба составляют 3.0, 1.6 и 5.1 мм. Эмалевая стенка зуба дифференцирована по толщине по мимомисному типу с более толстыми ведущими и более тонкими замыкающими краями. Индекс дифференциации эмали BTQ (=SDQ3) составляет 167, индекс SDQ6 – 152; измерения проведены согласно ван Колфсхотену и Тесакову [25, 26]. В среднем позднем плейстоцене Европы у водяных полевок выявлен эволюционный тренд от древних форм с мимомисной дифференциацией эмали к формам с современной микротусной дифференциацией [27], этот процесс используется в качестве надежного биохронометра [28, 29]. Водяная полевка с мимомисной эмалью является характерной формой теплых межледниковий середины среднего плейстоцена, MIS11 и MIS9, т.н. гольштейнского комплекса [30, 31] или “большого лихвинаˮ. Нахождение древней водяной полевки непосредственно в древнеэвксинских отложениях Тамани является первой точкой прямой корреляции малакологической и микротериологических стратиграфических шкал и повышает надежность возрастной модели Динской антиклинали. Четыре отобранных палеомагнитных образца показали прямую намагниченность.
4. Суглинки и супеси с линзами более грубообломочного материала; до 1.5 м.
На обзорных геологических картах в ядре брахискладки указаны мэотические и сарматские отложения [9, 10]. Однако недостаточная обнаженность не позволяет точно охарактеризовать отложения ядра. Лишь в одном прибрежном обнажении (2 на рис. 1 и рис. 2) неслоистые глины понта (?) содержат глыбы мэотических известняков, указывающие на существование близкого источника сноса, который сейчас не обнажен.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Развитие Динской антиклинали. Самым ранним зафиксированным свидетельством роста Динской брахиантиклинали является, возможно, уменьшение мощности чаудинских и древнеэвксинских отложений в пределах складки (1 и 3 на рис. 2; рис. 3) по сравнению с ее южным обрамлением (4 и 5 на рис. 2). Наиболее ярко рост складки проявился в деформации чаудинских и древнеэвксинских слоев, наклон которых достигает 40–42° на северо-восточном и 15–16° на юго-западном крыльях брахиантиклинали. Эта деформация произошла позже 0.4 млн. лет назад, но преимущественно в начале этого временного интервала, поскольку ее продолжение, выраженное уменьшением мощности средне-позднеплейстоценовых покровных отложений от крыльев к ядру складки и последующим искривлением их поверхности, гораздо слабее. Вместе с тем искривление поверхности показывает, что рост складки продолжался в конце плейстоцена, а повышенная концентрация береговых голоценовых оползней в ядре складки позволяет допустить, что деформация не прекратилась до сих пор.
Стоит отметить возможное объяснение залегания чаудинских отложений непосредственно на верхнепонтических в точке 3 (см. рис. 1 и рис. 2б, в). Поскольку на юго-западном берегу Динского залива широко развиты многочисленные оползни, то возможной причиной выявленного несогласия может быть оползневое смещение чаудинских отложений вниз по склону (рис. 2, в).
Другие проявления молодого складкообразования на Таманском п-ове. В общем виде о четвертичной складчатости свидетельствует прямой молодой тектонический рельеф п-ова: зоны антиклиналей выражены низкими возвышенностями, а зоны синклиналей – заливами, лиманами и седловинами между ними [32]. Один из примеров антиклиналей, длительно развивавшихся в течение четвертичного периода – Зародинская антиклинальная складка, протягивающаяся вдоль восточной части азовского побережья п-ова к северу от пос. За Родину [19, 33]. Антиклиналь сложена тремя толщами пород: нижней – преимущественно морскими мелководными глинами и песками верхнего куяльника (гелазий и нижний калабрий международной хроностратиграфической шкалы); средней – прибрежными песками с конгломератами и брекчиями в основании (средний и верхний калабрий); верхней – наземными суглинками и более грубообломочными отложениями (средний и поздний плейстоцен). Наиболее деформирована нижняя толща. Ее слои наклонены под углами до 70° и нарушены разломами амплитудой до десятков метров. Средняя толща заполнила понижения частично эродированного рельефа, возникшего при деформации нижней толщи и, в свою очередь, была смещена разломами и частично эродирована. В результате изгиба верхней толщи окончательно оформилась выраженная в рельефе антиклиналь, юго-западное крыло которой плавно погружается к Ахтанизовскому лиману, а северо-восточное крыло разрушается оползнями и морской абразией [19].
Важные данные об интенсивности неоплейстоценовых и голоценовых вертикальных движений и складчатых деформаций получены путем определения современного высотного положения береговых линий раннего и начала позднего неоплейстоцена, маркируемых соответственно чаудинскими и карангатскими морскими отложениями. На азовском побережье, воздымавшимся в послечаудинское время особенно интенсивно, подошва чаудинских слоев поднялась до 65–70 м, тогда как юго-восточнее и юго-западнее она находится на высотах 20–23 м [32]. В Таманском заливе чаудинские слои погружены на глубину до 45 м, а в Бугаз-Кизилташском и Витязевском лиманах – до 110 м [34, 35]. Таким образом, современное залегание чаудинских слоев изменяется в пределах 190 м в результате послечаудинских вертикальных движений [32].
Самым северным выходом карангатских отложений на азовском побережье п-ова является разрез между основанием косы Чушка и мысом Ахиллеон, где прибрежно-морской ожелезненный карангатский ракушняк мощностью 1.5 м залегает на цоколе высотой 6–7 м [8]. В присводовых частях плиоценовых антиклиналей, выраженных в четвертичной структуре пологими поднятиями, поверхность цоколя карангатских отложений повышается на 7 м, а кровля – на 8–12 м [32]. В Керченском проливе кровля карангатских отложений сейчас погружена на разную глубину [34, 36, 37]. Размах колебаний современного высотного положения карангатских отложений на Таманском и Керченском п-овах и в соседних акваториях составляет 50–60 м.
ЗАКЛЮЧЕНИЕ
1. Динская антиклинальная складка на северо-западе Таманского п-ова возникла в результате интенсивных тектонических движений, деформировавших морские отложения вплоть до древнеэвксинских (начало среднего неоплейстоцена); на северном крыле складки углы наклона слоев достигли 40–42°, а на юго-восточном – 15–16°.
2. Формирование антиклинали, начавшееся во время и сразу после древнеэвксинской трансгрессии (~0.4 млн л. н.), продолжалось до конца плейстоцена и голоцена в виде сводообразующей деформации покровного комплекса и террасовидной поверхности, которую он слагает.
3. Динская антиклиналь является выразительным примером позднечетвертичной складчатости и связанных с ней вертикальных смещений земной поверхности, характерных и для других районов Таманского п-ова.
Список литературы
Благоволин Н.С. Геоморфология Керченско-Таманской области. М.: Изд-во АН СССР, 1962. 201 с.
Попов Г.И. Плейстоцен Черноморско-Каспийских проливов. М.: Наука, 1983. 216 с.
www.stratigraphy.org [Электронный рecypc].
Cohen K.M. and Gibbard P.L. Global chronostratigraphical correlation table for the last 2.7 million years, version 2019 QI-500 // Quaternary International. 2019. Vol. 500. P. 20–31; https://doi.org/10.1016/j.quaint.2019.03.009
Измайлов Я.А. Строение и биостратиграфия отложений Азово-Черноморских неоплейстоценовых террас как свидетельства последовательного эрозионного углубления Черноморско-Средиземноморских проливов // Теория и методы современной геоморфологии / Мат-лы ХХХV Пленума Геоморфологической комиссии РАН. 2016. С. 221–225.
Свиточ А.А., Селиванов А.О., Янина Т.А. Палеогеографические события плейстоцена Понто-Каспия и Средиземноморья (материалы по реконструкции и корреляции). М.: Изд-во МГУ, 1998. 292 с.
Янина Т.А. Неоплейстоцен Понто-Каспия: биостратиграфия, палеогеография, корреляция. М: Геогр. факультет МГУ, 2012. 264 с.
Арсланов Х.А., Гей Н.А., Измайлов Я.А., Локшин Н.В., Герасимова С.А., Тертычный Н.И. О возрасте и климатических условиях формирования позднеплейстоценовых морских террас побережья Керченского пролива // Вестн. ЛГУ. Геология. География. 1983. № 12. Вып. 2. С. 69–79.
Государственная геологическая карта СССР. Сер. Крымская. Лист L-37-XIX, XXV. Масштаб 1:200000. 1971 г. / Ред. М.В. Муратов. Днепрогеология, Крымнефтеразведка.
Государственная геологическая карта РФ. Изд. второе. Сер. Кавказская. Лист L-37-XIX, XXV. Масштаб 1:200000. 2000 г. / Ред. К.О. Ростовцев. ФГУГП Кавказгеолсъемка.
Государственная геологическая карта РФ. Изд. второе. Сер. Кавказская. Карта четвертичных образований. Лист L-37-XIX, XXV. Масштаб 1:200000. 2000 г. / Ред. К.О. Ростовцев. ФГУГП Кавказгеолсъемка.
Цифровая модель рельефа https://search.earthdata.nasa.gov/search.
Бабак Е.В. Плиоценовые и четвертичные дрейссениды (Dreissenidae, Bivalvia) Эвксинского бассейна // Труды ПИН. 1983. Т. 204. 105 с.
Невесская Л.А., Парамонова Н.П., Бабак Е.В. Определитель плиоценовых двустворчатых моллюсков Юго-Западной Евразии // Труды ПИН. 1997. Т. 269. 267 с.
Nevesskaya L.A. History of the genus Didacna (Bivalvia: Cardiidae) // Paleontological Journal. 2007. Vol. 41. No. 9. P. 861–949.
Янина Т.А. Дидакны Понто-Каспия. М.–Смоленск: Маджента, 2005. 300 с.
Neubauer T.A., van de Velde S., Yanina T., and Wesselingh F.P. A late Pleistocene gastropod fauna from the northern Caspian Sea with implications for Pontocaspian gastropod taxonomy. ZooKeys 770. 2018. P. 43–103. https://doi.org/10.3897/zookeys.770.25365
Додонов А.Е., Тесаков А.С., Симакова А.Н. Таманское местонахождение фауны млекопитающих Синяя балка: новые данные по геологии и биостратиграфии // Ранний палеолит Евразии: новые открытия / С.А. Васильев и др. (ред.). Ростов-н/Д: ЮНЦ РАН, 2008. С. 53–57.
Тесаков А.С., Гайдаленок О.В., Соколов С.А., Фролов П.Д., Трифонов В.Г., Симакова А.Н., Латышев А.В., Титов В.В., Щелинский В.Е. Новые данные о новейшей структуре и возрасте нижнеплейстоценовых отложений северо-востока Таманского п-ова, Южное Приазовье // Геотектоника. 2019. № 5. С. 11–34.
Croitor R. Early Pleistocene small-sized deer of Europe // Hellenic Journal of Geosciences. 2006. Vol. 41. P. 89–117.
Алексеева Л.И. Териофауна раннего антропогена Восточной Европы // Тр. ГИН АН СССР. 1977. Вып. 300. 214 с.
Титов В.В. Крупные млекопитающие позднего плиоцена Северо-Восточного Приазовья. Ростов-н/Д: Изд. ЮНЦ РАН, 2008. 262 с.
Эберзин А.Г. Плиоценовые отложения Западного Предкавказья // Геология СССР. Т. IХ. Северный Кавказ. М.–Л.: Гос. изд-во геол. лит., 1947. С. 321–329.
Лебедева Н.А. Антропоген Приазовья // Тр. ГИН АН СССР. 1972. Вып. 215. 106 с.
van Kolfschoten T. The evolution of the mammal fauna in the Netherlands and the middle Rhine Area (Western Germany) during the late Middle Pleistocene // Mededelingen Rijks Geologische Dienst. 1990. Vol. 43–3. P. 1–69.
Tesakov A.S. Early stage of Allophaiomys evolution in eastern Europe // Paludicola. 1998. Vol. 2. No. 1. P. 98–105.
Heinrich W.-D. Neue Ergebnisse zur Evolution und Biostratigraphie von Arvicola (Rodentia, Mammalia) im Quartär Europas // Zeitschrift für Geologische Wissenschaften. 1987. Vol. 15. No. 3. P. 389–406.
Heinrich W.-D. Some aspects of the evolution and biostratigraphy of Arvicola (Mammalia, Rodentia) in the Central European Pleistocene // Int. Symp. on Evolution, Phylogeny and Biostratigraphy of Arvicolids (Rodentia, Mammalia). Eds. Fejfar O., Heinrich W.-D. Prague: Pfeil Verlag, 1990. P. 165–182.
Maul L.-Ch., Rekovets L., Heinrich W.-D., Keller Th., and Storch G. Arvicola mosbachensis (Schmidtgen 1911) of Mosbach 2: a basic sample for the early evolution of the genus and a reference for further biostratigraphical studies // Senckenbergiana Lethaea. 2000. Vol. 80 (1). P. 129–147.
Kalthoff D.C., Mörs Th., and Tesakov A.S. Late Pleistocene small mammals from the Wannenköpfevolcanoes (Neuwied Basin, western Germany) with remarks on the stratigraphicrange of Arvicola terrestris // Geobios. 2007. Vol. 40. P. 609–623.
van Kolfschoten T. The Palaeolithic locality Schöningen (Germany): a review of the mammalian record // Quaternary International. 2014. Vol. 326–327. P. 469–480.
Трифонов В.Г., Трифонов Р.В. Происхождение и экологические последствия фанагорийской регрессии Черного моря // Геоэкология. Инженерная геология. Гидрогеология. Геокриология. 2006. № 6. С. 509–521.
Измайлов Я.А. Щелинский В.Е. Геологическая ситуация раннепалеолитических местонахождений в Южном Приазовье на Таманском полуострове // Древнейший Кавказ: перекресток Европы и Азии. СПб.: ИИМК РАН. 2013. С. 20–39.
Щербаков Ф.А., Куприн П.Н., Забелина Э.К. Палеогеография Азово-Черноморья в позднем плейстоцене и голоцене // Палеогеография и отложения плейстоцена южных морей СССР. М.: Наука, 1977. С. 51–60.
Островский А.Б., Измайлов Я.А., Щеглов А.Н., Арсланов Х.А., Тертычный Н.И., Гей Н.А., Пиотровская Т., Муратов В.М., Щелинский В.Е., Балабанов И.П., Скиба С.И. Новые данные о стратиграфии и геохронологии плейстоценовых морских террас Черноморского побережья Кавказа и Керченско-Таманской области // Палеогеография и отложения плейстоцена южных морей СССР. М.: Наука, 1977. С. 61–68.
Никонов А.А. Признаки молодой тектонической активности в зонах Южно-Азовского и Керченского разломов // Геотектоника. 1994. № 5. С. 16–27.
Федоров П.В. Позднеплейстоценовая история Черного моря и развитие южных морей Европы // Палеогеография и отложения плейстоцена южных морей СССР. М.: Наука, 1977. С. 25–32.
Дополнительные материалы отсутствуют.
Инструменты
Геоморфология и палеогеография