

Вопросы ихтиологии, 2019, T. 59, № 1, стр. 105-109
Морфологическая дифференциация алтайского османа Oreoleuciscus humilis (Cyprinidae) в реке Туин, Долина Озёр, Монголия
А. Н. Мироновский 1, 2, *, Ю. Ю. Дгебуадзе 1, 3, Б. Мэндсайхан 4, Ю. В. Слынько 5
1 Институт проблем экологии и эволюции РАН − ИПЭЭ
Москва, Россия
2 Институт биологии внутренних вод РАН – ИБВВ
Ярославская обл., пос. Борок, Россия
3 Московский государственный университет
Москва, Россия
4 Институт географии и геологии Монгольской академии наук
Улан-Батор, Монголия
5 Институт морских биологических исследований РАН − ИМБИ
Республика Крым, Севастополь, Россия
* E-mail: adissa@mail.ru
Поступила в редакцию 04.12.2017
После доработки 28.02.2018
Принята к публикации 22.01.2018
Аннотация
Изучена изменчивость остеологических признаков алтайских османов рода Oreoleuciscus в р. Туин. На основе многомерного анализа 13 индексов, характеризующих пропорции осевого и висцерального черепов, в популяции выявлены две морфологически обособленные формы. Одну из них можно рассматривать как исходную при образовании карликовой формы оз. Орог в ходе циклической диверсификации. Полученный результат является ещё одним свидетельством возможности симпатрической диверсификации рыб в речных экосистемах.
Рыбы рода Oreoleuciscus (алтайские османы, или горные ельцы) обитают в западной части Монголии и сопредельных областях Российской Федерации на сравнительно небольшой территории Центрально-Азиатского бессточного бассейна, ограниченной горами Хангая и Алтая. Алтайские османы населяют пресноводные и солоноватоводные озёра и реки на высоте от 700 до 2200 м над уровнем моря. Нативная ихтиофауна этих водоёмов отличается видовой бедностью: кроме двух видов алтайских османов здесь обитали рыбы лишь родов Thymallus, Orthrias и Triplophysa (Dgebuadze et al., 2012). Это, видимо, и создало условия для возникновения ряда внутривидовых морфоэкологических форм алтайских османов в озёрных популяциях рода (Дгебуадзе, 1982; Баасанжав и др., 1983). В силу ряда причин речные популяции Oreoleuciscus были изучены слабее, и анализ небольших материалов по признакам внешней морфологии не давал оснований усомниться в их мономорфности (Баасанжав и др., 1983; Борисовец и др., 1985). Лишь недавно показано, что в популяции р. Завхан (котловина Больших Озёр) алтайский осман Потанина O. potanini образует две морфологически обособленные формы (Дгебуадзе и др., 2017).
Наша работа посвящена исследованию морфологической изменчивости алтайских османов вида Oreoleuciscus humilis, обитающих в р. Туин (Долина Озёр), на основе многомерного анализа остеологических признаков.
МАТЕРИАЛ И МЕТОДИКА
Исследование выполнено на материале, собранном на разных участках р. Туин: в среднем течении – в районе г. Баян-Хонгор, 46°03′16′′ с.ш. 100°45′59′′ в.д., 03.08.2006 г. (13 экз.); в нижнем течении – южнее сомона Богда-Ула, 45°06′10′′ с.ш. 100°46′33′′ в.д., 04.07.2008 г. (16 экз.) и 45°12′96′′ с.ш. 100°46′35′′ в.д., 31.08.2013 г. (9 экз.). В 2006 и 2013 гг. для лова использовали жаберные сети с ячеёй 12–60 мм (большинство рыб попали в сети с ячеёй 12–18 мм) и электролов; в 2008 г. рыбы пойманы накидной сетью.
Визуально группы морфологически различных особей в уловах не отмечены. Для камеральной обработки головы рыб вместе с костью плечевого пояса cleithrum фиксировали поваренной солью. После препарирования в лаборатории измеряли 13 остеологических параметров (рис. 1). Этот набор признаков характеризуется хорошей сопоставимостью данных при повторных измерениях как одним, так и разными операторами (Мироновский, 2006), что позволило получить ряд интересных результатов при изучении фенетического разнообразия крупных африканских усачей рода Barbus sensu lato (Mina et al., 1996) и озёрных форм алтайского османа Потанина O. potanini (Дгебуадзе и др., 2008; Мироновский и др., 2014).
Рис. 1.
Схема остеологических промеров: BL − базальная длина черепа; B1, B2, B3 − расстояние между внешними краями соответственно frontalia, pterotica и sphenotica; B4 − ширина черепа на уровне соединения frontale и pteroticum; HS1,HS2 − высота черепа на уровне соответственно изгиба parasphenoideum и заднего края parasphenoideum; Hm − высота hyomandibulare; dff − расстояние между крайними точками ветвей глоточного зуба; Op − высота operculum; De − длина dentale, Pop − длина по диагонали praeoperculum; Cltr − длина по диагонали кости плечевого пояса cleithrum.
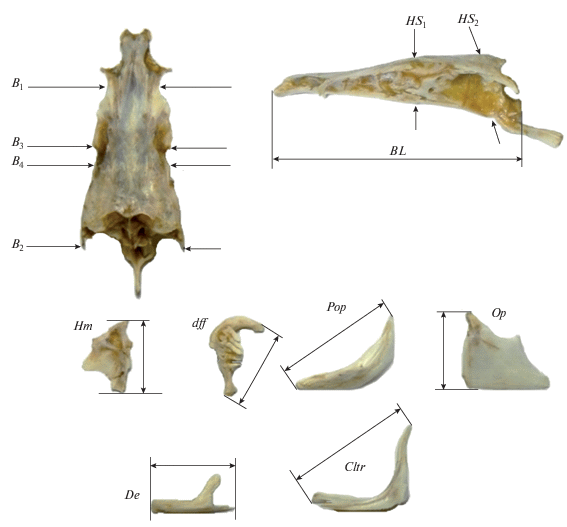
Данные промеров обрабатывали методами многомерной статистики с помощью программ пакета NTSYS 2.0 (Rohlf, 1998). В расчётах использовали индексы, рассчитанные как отношение абсолютных значений измерений к длине черепа (BL). Для оценки морфологических дистанций в многомерном пространстве индексов использовали обобщённое Евклидово расстояние, возведённое в квадрат. Кластерный анализ матриц сходства проводили невзвешенным парно-групповым методом. Результаты кластерного анализа представлены в виде дендрограммы. При анализе главных компонент (ГК) собственные векторы рассчитывали по корреляционной матрице. Длину вектора принимали равной 1. Наряду с традиционным методом анализа ГК использовали подход, основанный на построении многомерных онтогенетических каналов, хорошо зарекомендовавший себя при изучении фенетического разнообразия ряда видов карповых рыб (Mina et al., 1996; Дгебуадзе и др., 2008; Мироновский и др., 2014).
Изучаемые выборки разных лет в расчётах объединены, численность совокупной выборки составила 37 особей. Стандартная длина (SL) особей объединённой выборки варьирует от 78 до 136 мм. В качестве реперной использовали группу 39 сопоставимых по размерам (SL 105–152 мм) особей карликовой формы O. humilis, пойманных летом 2000 г. в оз. Орог, куда впадает р. Туин. Рыб в озере добывали жаберными сетями (ячея 10−60 мм), мальковым неводом (длина 10 м, ячея 3 мм) и сачком.
РЕЗУЛЬТАТЫ
В пространстве рассматриваемых признаков объекты анализируемого множества образуют отчётливо разобщённые совокупности, обозначенные Оk, Т1 и Т2 (рис. 2). На дендрограмме (рис. 2а) в состав кластера Оk вошли все особи карликовой формы Орога и одна особь Туина; кластеры Т1 и Т2 состоят только из особей, пойманных в реке. Состав одноимённых полигонов на плоскости двух первых ГК (рис. 2б) почти полностью совпадает с составом кластеров дендрограммы. Отличие лишь в том, что на рис. 2б в полигон Т1 попадает одна особь карликовой формы из оз. Орог.
Рис. 2.
Результаты многомерного анализа изменчивости рассматриваемых признаков алтайского османа Oreoleuciscus humilis популяции р. Туин и карликовой формы алтайского османа O. humilis оз. Орог: а − дендрограмма сходства особей, “жирные” ветви дендрограммы соответствуют особям формы Т1; б – распределение особей на плоскости первых двух главных компонент (ГК); особи формы: (⚫) − Т1 (р. Туин), (△) − Т2 (р. Туин), (⚪) − карликовой Оk (оз. Орог).
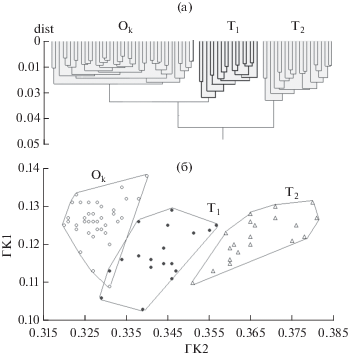
В кластер Т1 вошли 3 особи выборки 2008 г., 8 особей выборки 2006 г. и 7 особей выборки 2013 г. В кластере Т2 – 4 особи выборки 2006 г., 13 особей выборки 2008 г. и 2 особи выборки 2013 г. Следовательно, наблюдаемую морфологическую дифференциацию нельзя объяснить тем, что выборки взяты в разные годы и на разных участках реки. Изменение пропорций по мере роста рыб также не может быть причиной дифференциации: размерный ряд особей кластера Т1 (SL 83−136 мм) почти полностью включает в себя размерный ряд особей кластера Т2 (SL 78−101 мм). Таким образом, распределение особей на рис. 2 можно рассматривать как свидетельство того, что в р. Туин симпатрически обитают две морфологически обособленные формы вида O. humilis. При этом ни одна из форм не идентична карликовой форме оз. Орог. Между формами Оk и Т2 есть различия со значительным хиатусом на плоскости ГК (рис. 2б). В сочетании Оk−Т1 хиатуса нет, полигоны перекрываются, однако в зоне перекрывания находятся лишь по одной особи каждой формы. Между тем известно, что время от времени оз. Орог полностью высыхает в силу климатических колебаний и обитающие в нём рыбы гибнут или уходят в р. Туин. С началом влажного периода, когда озеро заполняется водой, озёрная популяция восстанавливается за счёт рыб р. Туин (Dgebuadze, 1995). Последний раз оз. Орог высыхало в 2004−2010 гг. (Dgebuadze et al., 2012; Dgebuadze, 2015). С 2010 г. озеро начало заполняться водой, к 2013 г. восстановилось почти полностью, произошла дифференциация на карликовую и озёрную формы, и часть особей из пробы 2013 г. в низовьях р. Туин вполне может представлять собой рыб, зашедших из озера. Этим, возможно, и объясняется близость совокупностей Оk и Т1. Однако принципиально важным результатом нашего анализа является обнаружение морфологических различий того или иного уровня между карликовой формой O. humilis из озера и обеими формами, обитающими в реке.
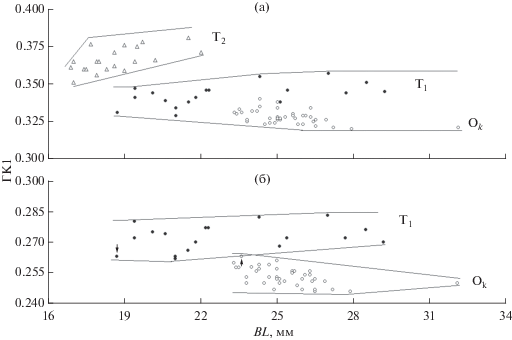
Для выявления структуры остеологических различий был использован метод построения онтогенетических каналов. На рис. 3а, где по оси ординат отложены значения ГК1-индексов рассматриваемых признаков, а по оси абсцисс – абсолютная длина черепа (BL), алтайские османы формы Т2 заметно мельче рыб двух других форм. Тем не менее, распределение точек на координатной плоскости не оставляет сомнений в обособленности онтогенетического канала этой формы от онтогенетических каналов форм Т1 и Оk. Уверенно говорить о разном онтогенетическом развитии форм Т1 и Оk данные рис. 3а не позволяют. Распределение особей на рис. 3а можно трактовать как упорядоченность положения разных форм в общем канале, где в верхней части находятся особи речной формы Т1, а в нижней – особи карликовой формы оз. Орог. Исключение из рассмотрения формы Т2, пересчёт ГК-признаков уже только для форм Т1 и Оk и, далее, повторное построение графика в координатах ГК1 и BL показывают разобщённость траекторий их развития (рис. 3б). Здесь в общем онтогенетическом канале особи формы Т1 и формы Оk распределены только при BL < 24 мм. С увеличением размеров происходит дивергенция. На рис. 3б отмечены (стрелки) две особи, попавшие в зону пересечения полигонов на рис. 2б. Как видим, это самая маленькая особь формы Т1 и одна из самых маленьких особей формы Оk, т. е. особи размеров, при которых дивергенция не началась и развитие шло в общем канале. Таким образом, онтогенетические каналы, представленные на рис. 3б, можно рассматривать как ещё одно подтверждение описанного ранее (Dgebuadze, 1995; Dgebuadze et al., 2012) происхождения форм O. humilis оз. Орог из речной популяции р. Туин.
Рис. 3.
Распределение особей форм Т1 и Т2 р. Туин и карликовой формы оз. Орог алтайского османа Oreoleuciscus humilis на плоскости многомерных онтогенетических каналов: а – все три формы, б – речная форма Т1 и карликовая форма озера; (↑) − особи, попавшие в зону пересечения полигонов форм Т1 и Оk на рис. 2б; остальные обозначения см. на рис. 2.
ВЫВОДЫ
1. Согласно результатам многомерного статистического анализа изменчивости 13 остеологических параметров популяция алтайских османов O. humilis р. Туин подразделена на две морфологически обособленные формы.
2. Взаимное положение многомерных онтогенетических каналов двух выявленных форм и карликовой формы O. humilis, образовывающейся в оз. Орог после его заполнения во влажные периоды климатических колебаний, хорошо согласуется с установленным ранее фактом происхождения популяции оз. Орог от особей р. Туин.
3. Анализ распределения изучаемых особей в пространстве рассматриваемых признаков не позволяет говорить о полной идентичности карликовой формы озера какой-либо из выявленных форм р. Туин.
БЛАГОДАРНОСТИ
Авторы искренне благодарны руководству Российско-монгольской комплексной биологической экспедиции РАН и АНМ за содействие в организации работ в Монголии; М.В. Мине (ИБР РАН) – за ценные советы и замечания к рукописи статьи.
Работа выполнена при частичной поддержке гранта РФФИ № 14-04-00022 А, Программы Президиума РАН № 41 “Биоразнообразие природных систем и биологические ресурсы России” и в рамках разделов государственного задания ИПЭЭ РАН № 0109-2018-0076 (А.Н. Мироновский, Ю.Ю. Дге-буадзе), ИБВВ РАН № АААА-А18-118012690222-4 (А.Н. Мироновский) и ИМБИ РАН № АААА-А18-118020890074-2 (Ю.В. Слынько).
Список литературы
Баасанжав Г., Дгебуадзе Ю.Ю., Демин А.Н. и др. 1983. Обзор видов ихтиофауны МНР // Рыбы Монгольской Народной Республики. М.: Наука. С. 102−224.
Борисовец Е.Э., Дгебуадзе Ю.Ю., Ермохин В.Я. 1985. Морфометрический анализ алтайских османов (Oreoleuciscus; Pisces, Cyprinidae) водоемов МНР: многомерный подход // Зоол. журн. Т. 64. № 8. С. 1199−1212.
Дгебуадзе Ю.Ю. 1982. Механизмы формообразования и систематика рыб рода Oreoleuciscus (Cyprinidae, Pisces) // Зоологические исследования в МНР. М.: Наука. С. 81−92.
Дгебуадзе Ю.Ю., Мина М.В., Мироновский А.Н. 2008. К оценке фенетических отношений алтайских османов (Oreoleuciscus, Cyprinidae) из трех озер Монголии по признакам черепа // Вопр. ихтиологии. Т. 48. № 3. С. 315−323.
Дгебуадзе Ю.Ю., Мироновский А.Н., Мендсайхан Б., Слынько Ю.В. 2017. Первый случай морфологической дифференциации алтайского османа Потанина Oreoleuciscus potanini (Cyprinidae, Actinopterigii) в реке // ДАН. Т. 473. № 2. С. 250−253.
Мироновский А.Н. 2006. Факторы, обуславливающие сопоставимость данных, полученных путем оценки пластических признаков рыб // Вопр. ихтиологии. Т. 46. № 2. С. 240−251.
Мироновский А.Н., Касьянов А.Н., Слынько Ю.В., Дгебуадзе Ю.Ю. 2014. Фенетические отношения и многомерные онтогенетические каналы экологических форм Алтайского османа Oreoleuciscus potanini (Cyprinidae) озера Ногон (Котловина Больших Озёр, Монголия) // Там же. Т. 54. № 1. С. 25–31.
Dgebuadze Yu.Yu. 1995. The land/inland-water ecotones and fish population of Lake Valley (West Mongolia) // Hydrobiologia. V. 303. P. 235−245.
Dgebuadze Yu.Yu. 2015. Central Asian Closed Basin: unique place of cyclic diversification of fish // Proc. Int. Conf. “Ecosystems of Central Asia under current conditions of socio-economic development”. V 2. Ulanbaatar. P. 29−33.
Dgebuadze Yu.Yu., Mendsaikhan B., Dulmaa A. 2012. Diversity and distribution of Mongolian fish: recent state, trends and studies // Erforsh. Biol. Ress. Mongolei (Halle/Saale). V. 12. P. 219–203.
Mina M.V., Mironovsky A.N., Dgebuadze Yu.Yu. 1996. Lake Tana large barbs: phenetics, growth and diversification // J. Fish Biol. V. 48. P. 383–404.
Rohlf F.J. 1998. NTSYS-pc: numerical taxonomy and multivariate analysis system (Version 2.0). N.Y.: Exeter software Publ., 31 p.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии