

Вопросы ихтиологии, 2019, T. 59, № 2, стр. 242-247
О малоизвестных акулах Etmopterus villosus (Etmopteridae) и Scymnodalatias sherwoodi (Somniosidae) из Тихого океана
В. Н. Долганов *
Национальный научный центр морской биологии Дальневосточного отделения РАН – ННЦМБ ДВО РАН
Владивосток, Россия
* E-mail: vndolganov@mail.ru
Поступила в редакцию 03.11.2017
После доработки 12.04.2018
Принята к публикации 31.05.2018
Аннотация
Приводится описание и распространение редких сквалидных акул Etmopterus villosus (16 экз. пойманы летом 1979 г. на подводных горах Маркус-Неккер) и Scymnodalatias sherwoodi (1 экз. пойман летом 1982 г. в эпипелагиали юго-восточной части Тихого океана). Первый вид известен только по голотипу от Гавайских о-вов, второй – по 3 экз. из Австрало-Новозеландского района.
Во время российских научно-исследовательских экспедиций в Тихом океане были собраны обширные материалы о хрящевых рыбах, в том числе о редких и малоизвестных. К примеру, глубоководная колючая акула Etmopterus villosus к настоящему времени известна только по описанию голотипа – ювенильного самца полной длиной (TL) 170 мм, пойманного у Гавайских о-вов на глубине 406–911 м (Gilbert, 1905; Compagno, 1984). Этот вид внутри рода Etmopterus относится к группе акул “granulosus”, у взрослых особей которых плакоидные чешуи в виде толстых коротких шипиков расположены на теле беспорядочно (Долганов, 1986). К этой группе относятся также обладающие обширными ареалами E. princeps из Северной Атлантики, E. granulosus (включая E. baxteri) из Южного полушария (Южная Африка, Австралия, Новая Зеландия, Южная Америка) и E. virens из Северо-Западной Атлантики, по сравнению с которыми точечный ареал E. villosus выглядит необычно. Летом 1979 г. во время экспедиции НПС “Мыс Юноны” на подводных горах Маркус-Неккер, примыкающих к Гавайскому хребту, были пойманы 16 неполовозрелых и взрослых особей E. villosus TL 368–652 мм.
Другой редкий вид – Scymnodalatias sherwoodi – к настоящему времени известен по трём поимкам у Новой Зеландии и Австралии (Last, Stevens, 2009), из которых только голотип имеет хорошее внешнее описание (Garrick, 1956) и сведения о числе позвонков (Taniuchi, Garrick, 1986). Данные о таких важных таксономических признаках, как строение черепа и птеригоподиев, число оборотов спирального клапана и клапанов сердечного артериального конуса, отсутствуют не только для этого вида, но и для других представителей рода Scymnodalatias (S. albicaudata, S. garricki и S. oligodon). Летом 1982 г. во время экспедиции на НИС “Пулковский меридиан” в эпипелагиали юго-восточной части Тихого океана был выловлен взрослый самец S. sherwoodi TL 730 мм.
В работе приводится описание этих малоизвестных видов и данные о их распространении и биологии.
Etmopterus villosus Gilbert, 1905
(рис. 1)
Рис. 1.
Etmopterus villosus: а – взрослый самец TL 624 мм; б, в – околосимфизные верхние зубы (б – самцов, в – самок); г, д – плакоидные чешуи (г – самец TL 613 мм, д – самка TL 652 мм).
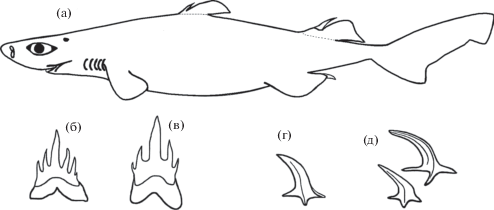
Материал. 7 самок TL 380–652 мм, 9 самцов TL 368–624 мм, 18°20′–24°10′ с.ш. 150°01′–171°55′ в.д., август 1979 г., подводные горы Маркус-Неккер, НПС “Мыс Юноны”, донный трал, глубина 960–1250 м. Коллектор Л.А. Борец.
Описание. Тело массивное. Рыло короткое, широкое, не коническое; его длина до глаз примерно равна горизонтальному диаметру глаза. Ноздри простые, без дополнительных лопастей, расположены вблизи вершины рыла. Расстояние между ноздрями составляет 2.5–3.0 раза в длине рыла до глаз. Ширина рта немного меньше длины рыла до рта. Межглазничное расстояние примерно равно длине рыла до глаз.
Первый спинной плавник (D1) меньше второго (D2). Интердорсальное расстояние примерно в два раза больше расстояния от конца основания D2 до начала верхней лопасти хвоста. Хвостовой плавник короткий, с хорошо выраженной подхвостовой выемкой. Длина его верхней лопасти примерно равна длине головы до 5-й жаберной щели. Задний конец грудного плавника не достигает уровня начала D1. Начало основания брюшных плавников ближе к D2, чем к D1. Антепекторальное расстояние много меньше пектовентрального.
Шипы спинных плавников хорошо развиты. Дермальные чешуи в виде толстых конусообразных шипиков с загнутыми назад вершинами (рис. 1г, 1д) расположены на теле беспорядочно. На рыле, животе и боковых отметинах, образованных скоплением фотофоров, шипики меньших размеров и прямостоящие; в задней части хвостового стебля и на хвосте они образуют продольные ряды, иногда слабовыраженные у взрослых особей.
Зубы верхней челюсти с 3–7 вершинами. Околосимфизные зубы взрослых самок 5-вершинные, самцов с 5–7 вершинами. Центральная вершина наибольшая, прямостоящая. Вторая пара боковых вершин у самок меньше, чем у самцов (рис. 1б, 1в). Коронки примерно в три раза длиннее корней. Зубы нижней челюсти с одной вершиной, сильно наклонены к углам рта, образуя единый режущий ряд.
Измерения 16 экз. TL 368–652 мм, в % TL (в скобках среднее значение признака). Расстояние от конца рыла: до D1 35.5–37.6 (36.4), до D2 60.3–64.3 (61.8), до хвоста 76.8–79.0 (78.0), до грудных плавников 21.3–24.7 (22.8), до брюшных плавников 52.9–56.3 (54.5). Длина рыла: до глаз 5.1–6.8 (6.1), до рта 8.7–10.7 (9.9). Длина головы: до 1-й жаберной щели 16.0–19.1 (17.8), до 5-й жаберной щели 21.5–24.7 (22.8). Горизонтальный диаметр глаза 4.0–5.7 (4.9). Расстояние между глаз 5.5–7.0 (6.5), между ноздрей 3.5–4.2 (3.9). Длина жаберных щелей: первой 1.9–2.4 (2.1), пятой 1.7–2.1 (2.0). Ширина рта 9.3–11.6 (10.5). Длина основания плавников: D1 4.8–6.3 (5.3), D2 6.7–8.1 (7.5), брюшного 5.4–9.0 (7.3). Высота плавников: D1 3.3–4.2 (3.6), D2 3.8–5.2 (4.7), брюшных 5.7–7.8 (6.8). Длина: переднего края грудных плавников 8.6–10.0 (9.1), верхней лопасти хвостового плавника 20.7–23.4 (22.0), нижней лопасти 9.8–13.2 (12.0). Расстояния: междорсальное 18.0–20.6 (19.6), от конца основания D2 до хвоста 8.7–11.1 (9.9), от конца основания брюшного плавника до начала хвоста 12.6–15.8 (14.2), от начала грудного плавника до начала брюшного 28.6–34.9 (31.0).
Туловищных позвонков 41–44 (42.6), позвонков хвостового стебля 13–18 (14.8), предхвостовых 56–59 (57.4), хвостовых 21–25 (23.5), общее число 79–83 (80.9). Число оборотов спирального клапана кишечника 9–11 (9.8). Зубная формула: верхняя челюсть 28–32 (30.9), нижняя – 37–54 (43.9).
Окраска. Тело и плавники коричнево-чёрные. Нижняя поверхность тела немного темнее, но боковые отметины у брюшных плавников и на хвосте почти незаметны.
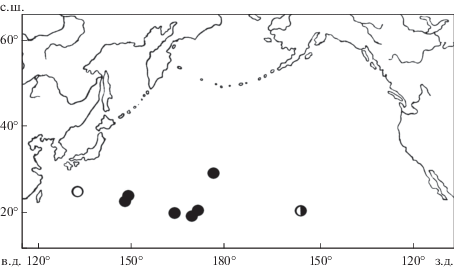
Распространение. Нахождение E. villosus на всём протяжении огромного подводного хребта Маркус-Неккер объединяет его ареал с Гавайским районом и подводными хребтами южнее Японии, где он известен по 1 экз. с подводного хребта Кюсю-Палау как E. princeps (Nakaya, 1982). Летом 2017 г. ярусолов “Палмер” обнаружил этот вид также на подводных горах южной части Северо-Западного подводного хребта. В неплохо изученных водах восточного побережья Японии E. villosus не обнаружен (Nakaya, Shirai, 1992; Shinohara, Matsuura, 1997; Shinohara et al., 2001; Fishes …, 2002; собственные данные). Таким образом, ареал этого вида находится под субтропическими водами (рис. 2), не распространяясь в более высокие широты как у близкородственных ему E. granulosus и E. princeps.
Рис. 2.
Распространение акулы Etmopterus villosus: (◑) – голотип, (⚫) – наши данные, (⚪) – “E. princeps” (Nakaya, 1982).
Биология. Максимальные известные размеры самцов – 624 мм, самок – 652 мм. Среди пойманных акул зрелыми были самцы TL > 600 мм и только одна самка TL 652 мм. Гонады зрелых самцов были длиной около 60 мм и шириной 10 мм, яйца половозрелой самки диаметром до 20 мм. В желудках преобладали миктофиды разных размеров, клювы и хрусталики кальмаров (в том числе и относительно крупных) и остатки ракообразных. Придонная температура воды в западной части ареала вида на глубине 1230–1240 м была 5.4°С. Широкое распространение на подводных горах на глубинах 960–1250 м, разделённых глубинами в несколько километров, и преобладающее питание пелагическими объектами предполагает возможность значительного отрыва от дна и длительное нахождение в толще воды.
Сравнительные замечания. Голотип E. villosus, самец-сеголетка TL 170 мм, не отличаясь от более взрослых особей числом позвонков (Yamakawa et al., 1986), имеет относительно большие размеры головы и глаз, большее расстояние от конца рыла до рта, до 1-й жаберной щели, до грудных плавников и более выраженные ряды дермальных чешуй в задней части тела и на хвосте. Соответственно, число зубов в нижней челюсти, междорсальное и пектовентральное расстояния и расстояние от конца рыла до D2 у него меньше. Это полностью соответствует возрастной изменчивости акул рода Etmopterus (Долганов и др., 2006а, 2006б). В пользу правильной идентификации пойманных экземпляров свидетельствует и тот факт, что заселение Гавайского района возможно только через подводные горы Маркус-Неккер. E. villosus отличается от E. princeps из Северной Атлантики меньшим числом моноспондильных позвонков (41–44 против 44–47) и их общим числом (79–83 против 81–86), меньшими размерами, более густым ошиплением и наличием боковых отметин (хотя и слабовыраженных), которые у E. princeps отсутствуют (Ebert, Stehmann, 2013). От E. granulosus из Южного полушария он хорошо отличается намного меньшим числом позвонков: моноспондильных (41–44 против 46–56), предхвостовых (56–59 против 59–67) и общим (79–83 против 86–89). Кроме этого, E. villosus имеет несколько меньшее число оборотов спирального клапана кишечника (9–11 против 10–13) (Yano, 1997). Следует заметить, что меньшее число позвонков E. villosus по сравнению с E. granulosus и E. princeps соответствует его обитанию в более тёплых низких широтах. E. virens из Карибского бассейна, который также обладает беспорядочно разбросанными по телу низкими крепкими шипиками, относится к карликовым видам. Максимальные размеры его взрослых особей не превышают 23 см (Compagno, 1984). Кроме ареала и размеров E. villosus отличается от него более массивным телом, более коротким хвостовым стеблем, который значительно меньше длины головы, пектовентрального и междорсального расстояний (у E. virens он примерно равен длине головы и пектовентральному расстоянию и в 1.4 раза больше междорсального). Окраска E. villosus намного темнее, и боковые отметины, образованные фотофорами на хвостовом стебле и хвосте, практически не видны, в то время как у E. virens они хорошо выражены (Compagno, 1984).
(рис. 3а, 3б)
Рис. 3.
Scymnodalatias sherwoodi, взрослый самец TL 730 мм из юго-восточной части Тихого океана: а – внешний вид, б – голова, вид снизу; в – неврокраниум; г, д – внешний вид и скелет птеригоподиев. R – рострум, AF – передняя фонтанель, V+VII –отверстия для глазничной ветви V и VII нервов; PPO – заглазничный отросток, POC – затылочный отросток, HP – hypopyle, SR – spur, RH – rhipidion, VM – вентральный маргинальный хрящ, SP – шип, DM – дорсальный маргинальный хрящ, DT – дорсальный терминальный хрящ, VT– вентральный терминальный хрящ.
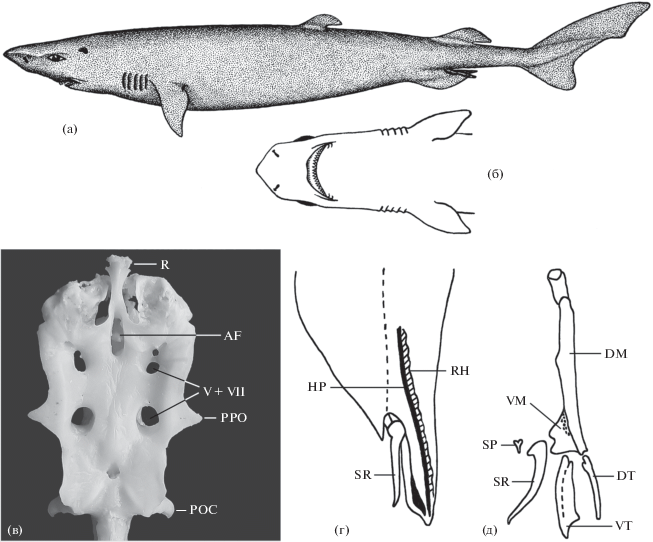
Материал. Половозрелый самец TL 730 мм, 44°52′ ю.ш. 110°00′ з.д., 17.07.1982 г., открытые воды юго-восточной части Тихого океана, НИС “Пулковский меридиан”, пелагический трал, горизонт лова 10 м. Коллектор П.В. Калчугин.
Описание. Рыло длинное, заострённое, его длина почти в два раза больше горизонтального диаметра глаза. Ноздри расположены вблизи вершины рыла. Клапаны ноздрей без дополнительных лопастей. Расстояние между ноздрями около двух раз в длине рыла до рта. Глаза в виде вытянутого овала, их горизонтальный диаметр в три раза меньше межглазничного расстояния. Предротовые щели имеются. Ширина рта примерно равна длине рыла до рта. Начало D1 ближе к началу брюшного плавника, чем к началу грудного. Начало D2 расположено примерно над серединой основания брюшных плавников. Антепекторальное расстояние составляет 57.8% пектовентрального. D1 намного меньше D2. Нет ни верхней, ни нижней предхвостовых ямок, но вдоль верхней и нижней поверхностей хвостового стебля имеются продольные ложбины. Хвостовой плавник с хорошо выраженной нижней лопастью и субтерминальной выемкой. Длина верхней лопасти хвоста немного больше междорсального расстояния, но меньше антепекторального.
Чешуи на теле мелкие, листовидные. Их задний край трёхлопастной, вдоль каждой лопасти имеется продольный гребень. Зубы в обеих челюстях одновершинные. В верхней челюсти они иглообразные, прямостоящие, с ассиметрично закруглёнными вершинами коронок и аркообразными корнями. Зубы нижней челюсти треугольные с острыми боковыми краями коронок, слегка наклонены к углам рта; высота их корней немного превышает высоту коронок. Зубная формула: верхняя челюсть – 29–1–28, нижняя – 15–1–15.
Туловищных позвонков 44, позвонков хвостового стебля 16, хвостовых 23, общее число 83. Число оборотов спирального клапана кишечника 41, рядов клапанов артериального конуса 5. Клапаны развиты слабо, даже в первом ряду. Печень двухлопастная.
Череп платибазальный. Передняя фонтанель маленькая, расположена далеко позади рострума. Рострум небольшой, сплющенный дорсовентрально. Заглазничный и затылочный отростки хорошо развиты. Теменное углубление некрупное. Мощный надглазничный гребень с необычно большими отверстиями для глазничной ветви V и VII нервов (рис. 3в).
Птеригоподии с большим шипом-шпорой и узкой кожной складкой пористой структуры (рипидион), прилегающей к глубокой продольной ложбине, разделяющей дорсальную и вентральную доли птеригоподия (гипопиле). Шпора плавно изогнута по всей длине и лишь её вершина изогнута значительно (рис. 3г). Скелет птеригоподиев (рис. 3д) состоит из дорсального и вентрального терминальных хрящей, прилегающих соответственно к дорсальному и вентральному маргинальным хрящам, крупной шпоры и расположенного за ней мелкого шипа.
Измерения, в % TL. Расстояние от конца рыла: до D1 43.6, до D2 65.5, до хвоста 79.7, до грудных плавников 22.3, до брюшных плавников 61.0. Длина рыла: до глаз 5.5, до ноздрей 2.3, до рта 8.5. Длина головы: до 1-й жаберной щели 17.1, до 5-й жаберной щели 21.6. Ширина головы 13.0. Высота тела у грудного плавника 13.0. Горизонтальный диаметр глаза 3.2. Расстояние между глаз 9.6, между ноздрей 4.0, между предротовыми щелями 8.9. Длина жаберных щелей: первой 2.7, третьей 2.7, пятой 3.0. Ширина рта 7.9. Длина основания плавников: D1 5.2, D2 6.3, брюшного 6.4. Высота плавников: D1 2.7, D2 3.0. Длина: переднего края грудных плавников 10.5, класперов 6.2, верхней лопасти хвостового плавника 20.5, нижней лопасти 10.3. Расстояния: междорсальное 16.4, от конца основания D2 до хвоста 7.9, от конца основания брюшного плавника до начала хвоста 11.0, от начала грудного плавника до начала брюшного 38.6.
Окраска. Тело коричнево-чёрное. На боках имеются небольшие светлые пятна и светлые участки с тёмными пятнами. Края плавников тёмные.
Распространение и биология. S. sherwoodi известен только по 3 экз., пойманным у Новой Зеландии и Южной Австралии на глубине 400–500 м (Last, Stevens, 2009). Описываемый экземпляр выловлен в верхней части эпипелагиали (ход верхней подборы пелагического трала на горизонте 10 м от поверхности) у берегов Южной Америки, что значительно расширяет представление о горизонтальном и вертикальном распространении этого вида. В желудке пойманного экземпляра находили клювы и хрусталики глаз крупных кальмаров.
Список литературы
Долганов В.Н. 1986. Описание новых видов акул семейства Squalidae (Squaliformes) из северо-западной части Тихого океана с замечаниями о валидности Etmopterus frontimaculatus // Зоол. журн. Т. 65. Вып. 1. С. 149–153.
Долганов В.Н., Королев М.Р., Гинанова Т.Т. 2006а. Морфологическое изучение акулы Etmopterus lucifer Jordan et Snyder, 1902 (Squalidae) из северо-западной части Тихого океана // Изв. ТИНРО. Т. 147. С. 261–264.
Долганов В.Н., Королев М.Р., Гинанова Т.Т. 2006б. Морфологическая характеристика акулы Etmopterus bigelowi Shirai et Tachikawa, 1993 (Squalidae) из северо-западной части Тихого океана // Там же. Т. 147. С. 265–268.
Compagno L.J.V. 1984. FAO species catalogue. Sharks of the world. An annotated and illustrated catalogue of shark species known to date. Pt. 1. Hexanchiformes to Lamniformes // FAO Fish. Synop. № 125. V. 4. P. 1–249.
Ebert D.A., Stehmann M. 2013. FAO species catalogue. Sharks, Batoids and Chimaeras of the North Atlantic. Rome: FAO, 523 p.
Fishes of Japan with pictorial keys to the species. 2002 / Ed. Nakabo T. Tokio: Tokai Univ. Press, 1749 p.
Garrick J.A.F. 1956. Studies on New Zealand Elasmobranchii. Pt. V. Scymnodalatias n. g. based on Scymnodon sherwoodi Archey, 1921 (Selachii) // Trans. Roy. Soc. N. Z. V. 83. № 3. P. 555–571.
Gilbert C.H. 1905. The deep-sea fishes of the Hawaiian Islands. Pt. II. Section II // Bull. US Fish. Comm. V. 23. № 2. P. 577–713.
Last P.R., Stevens J.D. 2009. Sharks and rays of Australia. Melbourne: CSIRO, 656 p.
Nakaya K. 1982. Squalidae // Fishes of the Kyushu-Palau Ridge and Tosa Bay / Eds. Okamura O. et al. Tokio: Jpn. Fish. Res. Conserv. Ass. P. 47–53, 310–315.
Nakaya K., Shirai S. 1992. Fauna and zoogeography of deep-benthic chondrichthyean fishes around the Japanese Archipelago // Jpn. J. Ichthyol. V. 39. № 1. P. 37–48.
Shinohara G., Matsuura K. 1997. Annotated checklist of deep-water fishes from Suruga Bay, Japan // Monogr. Nat. Sci. Mus. V. 12. P. 269–318.
Shinohara G., Endo H., Matsuura K. et al. 2001. Annotated checklist of the deepwater fishes from Tosa Bay, Japan // Ibid. V. 20. P. 283–343.
Taniuchi T., Garrick J.A.F. 1986. A new species of Scymnodalatias from the Southern Oceans, and comments on other squaliform sharks // Jpn. J. Ichthyol. V. 33. № 2. P. 119–134.
Yamakawa T., Taniuchi T., Nose Y. 1986. Review of the Etmopterus lucifer group (Squalidae) in Japan // Proc. 2-nd Int. conf. “Indo-Pacific fish biology”. Tokyo: Ichth. Soc. Jpn. P. 197–297.
Yano K. 1997. First record of the brown lanternshark, Etmopterus unicolor, from the waters around New Zealand, and comparison with the southern lanterhshark, E. granulosus // Ichthyol. Res. V. 44. № 1. P. 61–72.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии