

Вопросы ихтиологии, 2019, T. 59, № 2, стр. 137-152
Каспийская щиповка Sabanejewia caspia – давно известный, но практически не изученный вид семейства вьюновых (Cobitidae): особенности морфологии, кариотип, распространение и предполагаемые филогенетические связи
Е. Д. Васильева 1, *, В. П. Васильев 2
1 Зоологический музей Московского государственного университета
Москва, Россия
2 Институт проблем экологии и эволюции РАН − ИПЭЭ
Москва, Россия
* E-mail: vas_katerina@mail.ru
Поступила в редакцию 25.04.2018
После доработки 31.05.2018
Принята к публикации 19.06.2018
Аннотация
Дано иллюстрированное описание особенностей внешней морфологии, окраски (с учётом изменчивости), строения черепа и кариотипа каспийской щиповки Sabanejewia caspia. В результате сравнительного анализа на основе исследования коллекций по близким видам и данных литературы составлен новый видовой диагноз S. caspia, позволяющий избегать ошибок при идентификации. Представлены подтверждённые данные по распространению вида. Обсуждается современное состояние проблемы оценки филогенетических связей S. caspia и видов Sabanejewia. Обоснованы сомнения в правильности отнесения к виду S. caucasica гаплотипов Генбанка АУ059338 и АУ059339, широко используемых в филогенетике Cobitidae.
Среди видов щиповок (Cobitidae), включаемых в настоящее время в род Sabanejewia Vladykov, 1929, ранее всех была описана каспийская щиповка S. caspia (Eichwald, 1838). В отличие от многих других номинальных видов щиповок она всегда и всеми признавалась валидным видом и лишь перенесена из рода Cobitis Linnaeus, 1758, в котором была описана, в новый род Sabanejewia. Общепризнанность и/или обособленность ареала привели к тому, что S. caspia в наши дни является самым плохо изученным видом рода. До сих пор не изучен геном каспийской щиповки, хотя одним из результатов первой попытки оценить филогенетические связи в роде Sabanejewia на основе данных о изменчивости митохондриального генома (Perdices et al., 2003. Р. 295) является заключение о том, что полученные результаты “отрицают ранее предполагаемые сестринские связи между S. larvata и S. caspia”. Этот вывод был сделан на том основании, что, как оказалось, вид S. larvata (De Filippi, 1859) “не является филогенетически близкородственным ни к одной из мтДНК линий Sabanejewia” (Р. 295), выявленных в данной работе. Ещё раз надо отметить, что вида S. caspia в составе этих линий нет. Не опубликован кариотип каспийской щиповки, приведена лишь его формула в тезисах конференции (Vasil’ev, Vasil’eva, 1994) и отмечено, что он включает 50 хромосом, соответствующих исходному кариотипу для всех щиповок родов Cobitis и Sabanejewia (Васильев, 1985). Более того, морфологическая изменчивость и диагностическое значение морфологических характеристик каспийской щиповки оказались плохо изученными, что привело к досадным ошибкам в идентификации разных видов щиповок каспийского бассейна и, как следствие, к ошибочным определениям ареалов.
Цель настоящей работы – изучить изменчивость внешних морфологических характеристик и пигментации каспийской щиповки на основе собственных сборов и коллекции Зоологического музея МГУ (ЗММУ), выявить диагностические видовые признаки, впервые дать иллюстрированное описание структуры кариотипа, краниологических особенностей вида и на основе полученных данных определить границы ареала и оценить предполагаемые филогенетические связи S. caspia.
МАТЕРИАЛ И МЕТОДИКА
Исследования внешней морфологии и особенностей окраски проводили на основе выборки (36 экз.), собранной 20–21.04.1982 г. в Малом Кызылагачском заливе близ пос. Нариманабад. Помимо этой выборки изучен материал из коллекции ЗММУ (197 экз.): Р-3717 – р. Хирмандали, впадающая в Кызылагачский зал. (6 экз.); Р-3718 – Джильское озеро, Ленкоранская низменность, юго-западная часть Каспия (8 экз.); Р-3719 – Джильское озеро (5 экз.); Р-3720 – р. Кумбаши (= Гумбаши) (1 экз.); Р-3721 – Иранское побережье у устья р. Новая, низовья Сефидруда (9 экз.); Р-3723 – р. Кумбаши (1 экз.); Р-3724 – Джильское озеро (3 экз.); Р-3744 – Джильское озеро (2 экз.); Р-3777 – р. Хирмандали (1 экз.); Р-3779 – р. Кумбаши (1 экз.); Р-15917 – Малый Кызылагачский залив (36 экз.); Р-16747 – ахмаз (пресноводное озеро) в бассейне Куры у каналов АзПАС Мингечаур (2 экз.); Р-16755 – ахмаз там же (6 экз.); Р-16756 – Малый Кызылагачский залив (111 экз.); Р-16803 – каналы АзПАС Мингечаур (5 экз.).
У всех особей из коллекционных материалов анализировали особенности окраски, форму нижней губы, относительную длину усиков, форму суборбитального шипа, наличие утолщений боков тела впереди спинного плавника (считаются характерными для самцов в роде Sabanejewia) и измеряли общую длину тела (TL). Морфометрические признаки, обычно используемые в исследованиях щиповок (Bănărescu et al., 1972; Васильева, 1984; Васильева, Позняк, 1986), изучены у 36 экз. из пробы из Малого Кызылагачского залива. У 26 экз. из этой же пробы изучена морфология черепа по ранее опубликованной методике (Васильева, 1984). Для статистического анализа характеристик использовали стандартные унивариантные методы и коэффициент различия CD.
Структуру кариотипа исследовали в клетках передней части почек на воздушно-сухих препаратах, приготовленных по ранее опубликованной методике (Васильев, 1978). В общей сложности было изучено 55 метафазных пластинок от 12 особей, отловленных в 1985 г. в Малом Кызылагачском заливе и в протоке, которая соединяет залив с морем. Хромосомы классифицировали по системе, предложенной Леваном с соавторами (Levan et al., 1964).
Из коллекции ЗММУ были также изучены пробы из сборов Я. Гинзбурга, исходно идентифицированные как S. caspia, но в результате данного исследования отнесённые к другому описанному из бассейна Каспия виду – хвалынской щиповке Cobitis amphilekta Vasil’eva et Vasil’ev, 201211: Р-3730 – Малый Кызылагачский залив, против устья р. Кумбаши (2 самца); Р-3780 – о. Сара (= Сары), остров у западного берега Каспийского моря, к северу от города Ленкорань (1 самец).
Ряд морфологических признаков и окраска были изучены также у других видов рода Sabanejewia из коллекции ЗММУ.
S. romanica (Băcescu, 1943): P-9429 – р. Арджеш, бассейн Дуная, Румыния (4 экз., собраны и определены П. Бэнэреску); Р-9431 – р. Ведя, бассейн Дуная, Румыния (4 экз., собраны и определены П. Бэнэреску); Р-10291 – р. Гилонт (= Галбенэ), бассейн Дуная, Румыния (2 экз., собраны и определены П. Баческу как Cobitis caspia).
S. larvata: P-17289 – Италия, ров у д. Попола, в окрестностях Казале-Монферрато (3 экз., получены от Дж. Бальма); Р-17063 – Италия, Рио-делла-Оче близ Касальграссо (7 экз., получены от Дж. Дельмастро). Для сравнительного анализа использованы также результаты предшествующих наших исследований по видам рода Sabanejewia (Васильева, Позняк, 1986; Васильева, Васильев, 1988; Васильева, Раб, 1992) и другие данные литературы.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
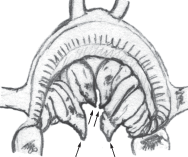
Внешняя морфология и окраска каспийской щиповки. У изученных нами особей D II (5½) 6½ (7, 7½), A II 5½ (6), P I 7–8, V I (4) 4½ (5, 5½), C I 12 I. С учётом данных по особям из р. Кумбаши (Абдурахманов, 1962) и из вод Ирана (Державин, 1934; Coad, 2017) изменчивость меристических признаков шире: D II–III (5½) 6½ (7, 7½), A I–II 5½ (6), P I 6–9, V I (4) 4½ (5, 5½, 6½), C I (7–11) 12 (13) I. Тело сжато с боков; спинной плавник начинается практически на уровне начала брюшных плавников; хвостовой стебель короткий, по нижнему краю несёт хорошо развитый кожистый гребень, по верхнему краю гребень хорошо развит лишь в задней части хвостового стебля. Голова небольшая, с коротким рылом и небольшими глазами (рис. 1). Суборбитальный шип хорошо развит, двуветвистый, передняя ветвь шипа существенно короче задней (рис. 2). Все три пары усиков длинные. В выборке из Малого Кызылагачского залива у большинства особей мандибулярный усик заходил за задний край глаза (25 из 36 экз., или 69.5%), реже доходил до заднего края (11.1%) или чуть не достигал заднего края глаза (19.4%). Только в пробе Р-16756, где преобладали мелкие экземпляры TL < 6 см, встречались отдельные особи с относительно короткими усиками: мандибулярные доходили не далее переднего края глаза. Для щиповок Ирана указывается, что задний усик у них заходит за задний край глаза (Coad, 2017). Чешуя у каспийской щиповки более или менее округлая, с большим центром и малым числом поперечных бороздок (Nalbant, Bianco, 1998; настоящая работа). Нижняя губа двулопастная, на внутренней стороне каждой лопасти по два усиковидных выроста: нижний представляет собой удлинение окончания внутренней стороны губы, а верхний образован удлинением верхней складки (рис. 1в, 3). Эти “усики” обычно заметны даже невооружённым глазом (рис. 1в). В выборке из Малого Кызылагачского залива только у 2 из 41 экз. (4.9%) верхние выросты-усики не были чётко заметны на одной из губных лопастей. В то же время у всех рыб из коллекции ЗММУ, даже у молоди, губы были типичной для вида формы; выросты-усики хорошо заметны и на рисунке строения рта у каспийской щиповки в публикации Налбанта и Бьянко (Nalbant, Bianco, 1998. Fig. 23b ).
Рис. 1.
Взрослая самка Sabanejewia caspia из Малого Кызылагачского залива (Р-15917), TL 62 мм: а, б – вид сбоку и сверху; в – голова снизу; (↑) – усиковидные выросты на правой лопасти нижней губы.

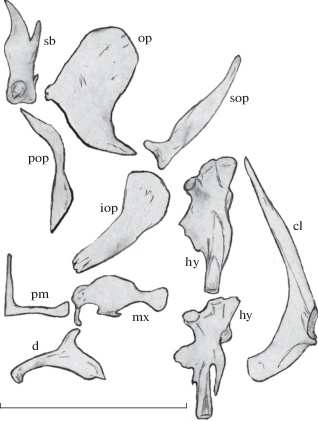
Рис. 2.
Строение костей черепа Sabanejewia caspia из Малого Кызылагачского залива: cl – cleithrum, d – dentale, hy – hyomandibulare (представлены два варианта формы), iop – interoperculum, mx – maxillare, op – operculum, pm – praemaxillare, pop – praeoperculum, sb – суборбитальный шип, sop – suboperculum. Масштаб: 0.5 см.
Морфометрические признаки щиповок из Малого Кызылагачского залива представлены в табл. 1. В выборке пол по данным вскрытия был определён для четырёх самцов и 14 самок; по большинству признаков различия между полученными для них средними значениями не превышают величину ошибки среднего значения у самок и только в шести случаях можно предполагать наличие достоверных различий (pD, lpc, h, lA, lB1, lB3). Однако эти различия между средними значениями у самцов и самок невелики и могут трактоваться не как половые, а как межвыборочные, тем более что выборка самцов не является репрезентативной (вследствие чего статистическое сравнение было бы некорректным). Не выявлено половых различий по данным признакам и в более многочисленной выборке каспийской щиповки из р. Кумбаши (Абдурахманов, 1962). Поэтому мы объединили всех особей из нашей выборки для сравнения с аналогичной суммарной выборкой из р. Кумбаши. Достоверные различия выявлены по 11 признакам (aD, pD, lpc, H, h, lP, lV, c, ao, o, io). По большинству характеристик пределы изменчивости двух выборок существенно перекрываются, и СD не достигает формально подвидового уровня 1.28. Только по величине межглазничного расстояния различия больше (СD = 1.66). В определённой степени это может быть связано (как, впрочем, и различия по другим пластическим признакам) с размерной изменчивостью (щиповки из р. Кумбаши существенно мельче щиповок из Малого Кызылагачского залива) и с тем, что промеры получены разными операторами.
Таблица 1.
Некоторые морфометрические признаки каспийской щиповки Sabanejewia caspia из Малого Кызылагачского залива и р. Кумбаши
| Признак | Малый Кызылагачский залив (наши данные) | Р. Кумбаши (Абдурахманов, 1962) | ||
|---|---|---|---|---|
| Самки (n = 14) | Самцы (n = 4) | Оба пола (n = 36) | Оба пола (n = 100) | |
| TL, мм | $\frac{{60.0{\kern 1pt} --{\kern 1pt} 79.3}}{{66.1 \pm 1.61}}$ | $\frac{{55.0{\kern 1pt} --{\kern 1pt} 69.0}}{{64.3}}$ | $\frac{{55.0{\kern 1pt} --{\kern 1pt} 79.3}}{{64.9 \pm 0.85}}$ | $\frac{{39.0{\kern 1pt} --{\kern 1pt} 58.0}}{{45.4}}$ |
| SL, мм | $\frac{{51.2{\kern 1pt} --{\kern 1pt} 65.2}}{{56.6 \pm 1.25}}$ | $\frac{{47.5{\kern 1pt} --{\kern 1pt} 60.3}}{{55.3}}$ | $\frac{{47.5{\kern 1pt} --{\kern 1pt} 65.3}}{{55.7 \pm 0.70}}$ | $\frac{{34.6{\kern 1pt} --{\kern 1pt} 50.5}}{{39.3}}$ |
| В % SL | ||||
| aD | $\frac{{48.5{\kern 1pt} --{\kern 1pt} 53.4}}{{50.2 \pm 0.40}}$ | $\frac{{48.7{\kern 1pt} --{\kern 1pt} 52.9}}{{50.3}}$ | $\frac{{46.7{\kern 1pt} --{\kern 1pt} 53.4}}{{50.0 \pm 0.23}}$ | $\frac{{48.2{\kern 1pt} --{\kern 1pt} 55.4}}{{51.5 \pm 0.07}}$ |
| pD | $\frac{{41.2{\kern 1pt} --{\kern 1pt} 45.4}}{{43.8 \pm 0.35}}$ | $\frac{{43.6{\kern 1pt} --{\kern 1pt} 47.2}}{{45.0}}$ | $\frac{{41.2{\kern 1pt} --{\kern 1pt} 47.2}}{{44.3 \pm 0.22}}$ | $\frac{{39.0{\kern 1pt} --{\kern 1pt} 44.8}}{{42.0 \pm 0.09}}$ |
| aV | $\frac{{47.6{\kern 1pt} --{\kern 1pt} 52.8}}{{50.4 \pm 0.43}}$ | $\frac{{48.1{\kern 1pt} --{\kern 1pt} 51.6}}{{49.8}}$ | $\frac{{47.6{\kern 1pt} --{\kern 1pt} 52.8}}{{50.1 \pm 0.21}}$ | – |
| aA | $\frac{{71.9{\kern 1pt} --{\kern 1pt} 80.9}}{{76.6 \pm 0.70}}$ | $\frac{{73.9{\kern 1pt} --{\kern 1pt} 77.9}}{{76.6}}$ | $\frac{{71.9{\kern 1pt} --{\kern 1pt} 80.9}}{{76.4 \pm 0.32}}$ | – |
| lpc | $\frac{{13.5{\kern 1pt} --{\kern 1pt} 17.8}}{{15.7 \pm 0.31}}$ | $\frac{{15.9{\kern 1pt} --{\kern 1pt} 18.0}}{{16.8}}$ | $\frac{{13.5{\kern 1pt} --{\kern 1pt} 18.0}}{{15.8 \pm 0.17}}$ | $\frac{{12.5{\kern 1pt} --{\kern 1pt} 17.9}}{{14.7 \pm 0.06}}$ |
| H | $\frac{{13.1{\kern 1pt} --{\kern 1pt} 20.3}}{{16.6 \pm 0.65}}$ | $\frac{{14.7{\kern 1pt} --\,16.6}}{{15.4}}$ | $\frac{{13.1{\kern 1pt} --{\kern 1pt} 20.3}}{{16.0 \pm 0.29}}$ | $\frac{{14.8{\kern 1pt} --{\kern 1pt} 20.8}}{{17.4 \pm 0.09}}$ |
| h | $\frac{{8.4{\kern 1pt} --{\kern 1pt} 11.4}}{{9.8 \pm 0.22}}$ | $\frac{{9.9{\kern 1pt} --{\kern 1pt} 11.4}}{{10.7}}$ | $\frac{{8.4{\kern 1pt} --{\kern 1pt} 11.5}}{{10.0 \pm 0.13}}$ | $\frac{{7.7{\kern 1pt} --{\kern 1pt} 11.5}}{{9.3 \pm 0.08}}$ |
| Cr | $\frac{{6.2{\kern 1pt} --{\kern 1pt} 10.2}}{{8.3 \pm 0.33}}$ | $\frac{{6.5{\kern 1pt} --{\kern 1pt} 8.3}}{{7.6}}$ | $\frac{{6.2{\kern 1pt} --{\kern 1pt} 10.2}}{{7.9 \pm 0.16}}$ | – |
| Crm | $\frac{{6.0{\kern 1pt} --{\kern 1pt} 8.6}}{{7.1 \pm 0.28}}$ | $\frac{{5.5{\kern 1pt} --{\kern 1pt} 7.1}}{{6.6}}$ | $\frac{{5.5{\kern 1pt} --8.6}}{{6.9 \pm 0.13}}$ | – |
| cr | $\frac{{1.4{\kern 1pt} --{\kern 1pt} 2.9}}{{1.9{\text{ }} \pm 0.09}}$ | $\frac{{1.6{\kern 1pt} --{\kern 1pt} 2.1}}{{1.8}}$ | $\frac{{1.4{\kern 1pt} --{\kern 1pt} 2.9}}{{1.9 \pm 0.04}}$ | – |
| P–V | $\frac{{28.1{\kern 1pt} --{\kern 1pt} 34.0}}{{31.3 \pm 0.53}}$ | $\frac{{28.8{\kern 1pt} --{\kern 1pt} 32.8}}{{31.1}}$ | $\frac{{28.1{\kern 1pt} --{\kern 1pt} 34.0}}{{31.0 \pm 0.26}}$ | $\frac{{27.5{\kern 1pt} --{\kern 1pt} 34.9}}{{30.5 \pm 0.12}}$ |
| lP | $\frac{{15.3{\kern 1pt} --{\kern 1pt} 19.0}}{{17.2 \pm 0.26}}$ | $\frac{{15.4{\kern 1pt} --{\kern 1pt} 18.5}}{{17.0}}$ | $\frac{{15.3{\kern 1pt} --{\kern 1pt} 19.8}}{{17.4 \pm 0.17}}$ | $\frac{{13.3{\kern 1pt} --{\kern 1pt} 18.2}}{{15.6 \pm 0.06}}$ |
| lV | $\frac{{12.1{\kern 1pt} --{\kern 1pt} 14.2}}{{13.0 \pm 0.17}}$ | $\frac{{11.9{\kern 1pt} --{\kern 1pt} 14.1}}{{13.1}}$ | $\frac{{11.7{\kern 1pt} --{\kern 1pt} 14.6}}{{13.1 \pm 0.12}}$ | $\frac{{10.2{\kern 1pt} --{\kern 1pt} 14.3}}{{11.9 \pm 0.08}}$ |
| lD | $\frac{{7.3{\kern 1pt} --{\kern 1pt} 9.8}}{{8.4 \pm 0.22}}$ | $\frac{{6.5{\kern 1pt} --{\kern 1pt} 10.2}}{{8.3}}$ | $\frac{{6.5{\kern 1pt} --{\kern 1pt} 10.2}}{{8.4 \pm 0.13}}$ | $\frac{{7.1{\kern 1pt} --{\kern 1pt} 9.9}}{{8.1 \pm 0.07}}$ |
| lA | $\frac{{6.0{\kern 1pt} --{\kern 1pt} 9.3}}{{7.3 \pm 0.28}}$ | $\frac{{7.8{\kern 1pt} --{\kern 1pt} 9.0}}{{8.5}}$ | $\frac{{6.0{\kern 1pt} --{\kern 1pt} 9.3}}{{7.7 \pm 0.14}}$ | $\frac{{5.8{\kern 1pt} --{\kern 1pt} 9.8}}{{7.4 \pm 0.05}}$ |
| hD | $\frac{{12.7{\kern 1pt} --{\kern 1pt} 15.7}}{{14.0 \pm 0.23}}$ | $\frac{{13.4{\kern 1pt} --{\kern 1pt} 15.2}}{{14.2}}$ | $\frac{{12.7{\kern 1pt} --{\kern 1pt} 15.7}}{{14.3 \pm 0.14}}$ | $\frac{{12.0{\kern 1pt} --{\kern 1pt} 16.7}}{{14.3 \pm 0.14}}$ |
| c | $\frac{{18.3{\kern 1pt} --{\kern 1pt} 20.8}}{{19.4 \pm 0.16}}$ | $\frac{{18.2{\kern 1pt} --{\kern 1pt} 20.6}}{{19.1}}$ | $\frac{{17.8{\kern 1pt} --{\kern 1pt} 20.8}}{{19.2 \pm 0.11}}$ | $\frac{{18.0{\kern 1pt} --{\kern 1pt} 22.2}}{{20.3 \pm 0.05}}$ |
| В % с | ||||
| lB1 | $\frac{{16.5{\kern 1pt} --{\kern 1pt} 31.4}}{{20.7{\text{ }} \pm 1.07}}$ | $\frac{{20.4{\kern 1pt} --{\kern 1pt} 28.6}}{{24.5}}$ | $\frac{{16.5{\kern 1pt} --{\kern 1pt} 31.4}}{{23.2 \pm 0.69}}$ | – |
| lB3 | $\frac{{22.3{\kern 1pt} --{\kern 1pt} 40.6}}{{29.8 \pm 1.48}}$ | $\frac{{32.4{\kern 1pt} --{\kern 1pt} 39.6}}{{35.4}}$ | $\frac{{22.3{\kern 1pt} --{\kern 1pt} 40.6}}{{32.1 \pm 0.87}}$ | – |
| ao | $\frac{{28.8{\kern 1pt} --{\kern 1pt} 39.2}}{{35.1 \pm 0.80}}$ | $\frac{{32.7{\kern 1pt} --{\kern 1pt} 37.6}}{{35.5}}$ | $\frac{{28.8{\kern 1pt} --{\kern 1pt} 39.2}}{{35.6 \pm 0.39}}$ | $\frac{{34.4{\kern 1pt} --{\kern 1pt} 44.5}}{{39.3 \pm 0.11}}$ |
| o | $\frac{{15.8{\kern 1pt} --{\kern 1pt} 20.6}}{{18.0 \pm 0.45}}$ | $\frac{{16.8{\kern 1pt} --{\kern 1pt} 18.9}}{{18.0}}$ | $\frac{{15.8{\kern 1pt} --{\kern 1pt} 20.6}}{{18.5 \pm 0.22}}$ | $\frac{{13.4{\kern 1pt} --{\kern 1pt} 20.8}}{{16.3 \pm 0.12}}$ |
| po | $\frac{{48.0{\kern 1pt} --{\kern 1pt} 55.7}}{{52.9 \pm 0.65}}$ | $\frac{{51.5{\kern 1pt} --{\kern 1pt} 55.9}}{{53.3}}$ | $\frac{{48.0{\kern 1pt} --{\kern 1pt} 57.8}}{{53.5 \pm 0.39}}$ | $\frac{{48.8{\kern 1pt} --{\kern 1pt} 60.3}}{{54.3 \pm 0.22}}$ |
| hc | $\frac{{47.8{\kern 1pt} --{\kern 1pt} 60.6}}{{55.1 \pm 1.01}}$ | $\frac{{51.0{\kern 1pt} --{\kern 1pt} 63.1}}{{54.8}}$ | $\frac{{47.8{\kern 1pt} --{\kern 1pt} 63.6}}{{56.0 \pm 0.67}}$ | $\frac{{66.0{\kern 1pt} --{\kern 1pt} 83.0}}{{73.2 \pm 0.22}}$ |
| b | $\frac{{5.6{\kern 1pt} --{\kern 1pt} 9.9}}{{8.4 \pm 0.36}}$ | $\frac{{7.1{\kern 1pt} --{\kern 1pt} 8.9}}{{8.3}}$ | $\frac{{5.6{\kern 1pt} --{\kern 1pt} 10.8}}{{8.4 \pm 0.21}}$ | – |
| io | $\frac{{11.7{\kern 1pt} --{\kern 1pt} 19.6}}{{16.7 \pm 0.57}}$ | $\frac{{15.3{\kern 1pt} --{\kern 1pt} 19.8}}{{17.1}}$ | $\frac{{11.7{\kern 1pt} --{\kern 1pt} 20.8}}{{17.2 \pm 0.34}}$ | $\frac{{18.8{\kern 1pt} --{\kern 1pt} 26.2}}{{22.4 \pm 0.11}}$ |
Примечание. Здесь и в табл. 2: TL – общая длина тела, SL – стандартная длина тела; aD, pD, aV, aA – антедорсальное, постдорсальное, антевентральное и антеанальное расстояния; lpc – длина хвостового стебля, H – наибольшая высота тела перед спинным плавником, h – высота хвостового стебля, Cr – толщина тела перед спинным плавником, Crm – толщина тела на уровне спинного плавника, cr – толщина хвостового стебля, P–V – пектовентральное расстояние, lP – длина грудного плавника, lV – длина брюшного плавника, lD – длина основания спинного плавника, lA – длина основания анального плавника, hD – высота спинного плавника, с – длина головы, lB1 – длина усиков 1-й пары, lB3 – длина мандибулярных усиков (3-я пара), ao – длина рыла, о – горизонтальный диаметр глаза, po – заглазничное расстояние, hс – высота головы на уровне середины глаза, b – расстояние между ростральными усиками, io – межглазничное расстояние. Над чертой − пределы изменчивости, под чертой – среднее значение и его ошибка, n – число исследованных особей, экз.
По всему телу каспийской щиповки разбросаны мелкие пятнышки тёмно-бурого цвета, которые сливаются в непрерывную продольную тёмную полосу, идущую вдоль средней линии бока, тёмную полосу вдоль середины спины и пятна неправильной формы, хаотично разбросанные между серединой бока и спиной и ниже середины бока (рис. 1а, 1б). У многих особей ниже полосы, идущей посредине бока, имеется вторая менее чёткая полоса; ниже этой полосы пятнышки тёмного пигмента обычно более редкие, хотя могут доходить и до средней линии брюха; чаще нижняя часть брюха не пигментирована. У основания хвостового плавника пигментные крапинки сгущаются в два нечётких краевых пятна, соединённых более узкой неровной полоской. Голова густо усыпана тёмными пятнышками, на фоне которых слабо проглядывает тёмная полоса от конца рыла до переднего края глаза. На непарных плавниках разбросанные пятнышки тёмного пигмента объединяются в неправильные поперечные ряды.
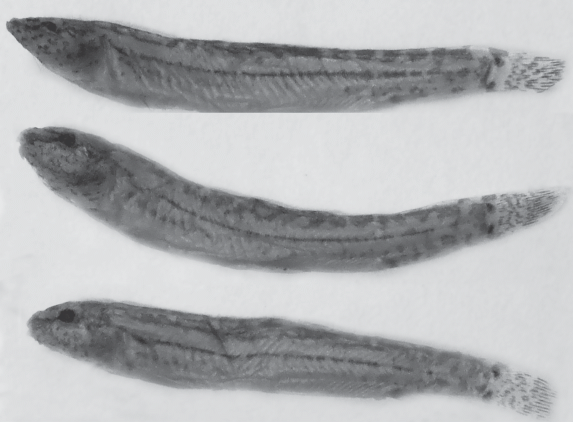
В отличие от взрослых рыб у молоди каспийской щиповки (TL < 35 мм из проб Р-3717, Р-3721, Р-3777, Р-3779, Р-16755, Р-16803) тёмные пятна у основания хвостового плавника обыкновенно разобщены и вдоль спины вместо единой тёмной полосы хорошо заметны разделенные светлыми участками пятна (рис. 4).
Сравнительные замечания. Ранее Державин (1934. С. 113) указывал, что данное Эйхвальдом (Eichwald, 1838) описание каспийской щиповки “недостаточно ясное и не снабжено рисунком”, а более позднее описание Кесслера (Кесслер, 1877) основано на смешанном материале. На самом деле описание Кесслера также не снабжено иллюстрацией и фактически содержит лишь одну характеристику, которую можно считать диагностической: “тёмные пятнышки, расположенные по длине боковой линии или непосредственно под нею, всегда более или менее сливаются между собой в одну неразрывную полоску” (Кесслер, 1877. С. 183). При этом приведённое им число лучей в хвостовом плавнике (16–17) ясно показывает, что лучи считали у какого-то вида рода Cobitis, поскольку для всех видов рода Sabanejewia (как и для каспийской щиповки) характерно наличие не более 12 ветвистых (или 14 развитых) лучей в хвостовом плавнике. В отличие от публикации Кесслера данная Эйхвальдом (Eichwald, 1838. Р. 133) характеристика не позволяет сомневаться в том, что им была изучена именно каспийская щиповка, поскольку для вида указано несколько важных признаков: 1) наличие чёткой непрерывной тёмно-коричневой продольной полосы “taenia longitudinalis fusca cohaerens, distincta, nec interrupta”, 2) чёрная окраска спины “dorsum e fusco nigroque nebulosum”, 3) чёрное основание хвостового плавника “caudalis basi nigra”, 4) покрытая чёрными пятнышками голова “caput nigromaculatum”. В Ленкорани, откуда была описана каспийская щиповка, нет других видов семейства Cobitidae с такими характеристиками. Приведённые Эйхвальдом 15 лучей в хвостовом плавнике (т.е. 13 ветвистых) могут быть не только результатом опечатки, как считал Державин (1934), но и результатом внутривидовой изменчивости признака, диапазон которой приведён нами выше.
Впервые рисунок каспийской щиповки был дан Державиным (1934. Abb. 10 ). На нём, в отличие от более поздних иллюстраций Берга (1949), основанных на разных видах (Васильева, 1995; Васильева, Васильев, 2012), прекрасно переданы особенности видовой окраски, а в характеристике вида отмечено наличие сплошной тёмной полосы не только по бокам тела, но и на спине. Однако, к сожалению, работа Державина не получила широкого признания.
Начиная с работы Кесслера (1877) наличие тёмной непрерывной полосы вдоль середины бока стало единственным или ведущим (наряду с формой суборбитального шипа) ключевым признаком для идентификации каспийской щиповки (Берг, 1916, 1949; Абдурахманов, 1962; Казанчеев, 1981; Kottelat, Freyhof, 2007; Богуцкая и др., 2013). При этом в характеристике вида Берг (1916, 1949) указывал, что на этой полосе иногда располагаются бурые пятна. На практике это приводило к тому, что за каспийскую щиповку ошибочно, как было показано нами (Васильева, 1995; Васильева, Васильев, 2012), принимали музейные экземпляры щиповок рода Cobitis: C. amphilekta и C. melanoleuca Nichols, 1925. У таких особей в результате хранения были утрачены пятна вдоль середины бока, но зато была видна просвечивающая тёмная прослойка соединительной ткани между спинными и брюшными мышцами. Кроме этого в статусе подвида каспийской щиповки – Cobitis caspia romanica Băcescu, 1943 – были описаны щиповки из р. Братья (система р. Арджеш, бассейн Дуная) в Румынии. У этих щиповок имеется тёмная полоса вдоль середины бока, но в отличие от полосы у каспийской щиповки она, как уже указывал ранее Берг (1949. С. 899), также “представляет собой не окраску покровов, а пигментацию, свойственную соединительной ткани горизонтальной перегородки”. Помимо этой полосы у румынской щиповки вдоль середины бока проходит продольный ряд из небольшого числа (8–13) узких, вертикально удлинённых пятен. Таким образом, румынская щиповка существенно отличается по окраске от каспийской, однако она формально вполне соответствует характеристикам S. caspia в трактовке Берга (1949).
На самом деле по общей пигментации тела на каспийскую щиповку более всего похож североитальянский вид S. larvata, у которого мелкие округлые многочисленные (15–27) пятна вдоль середины бока часто сливаются в одну непрерывную полосу, выше и ниже которой разбросаны небольшие пятна неправильной формы, часто образующие дополнительную продольную полосу ниже полосы, идущей вдоль середины бока (Kottelat, Freyhof, 2007; настоящая работа). На основе сходства в окраске Баческу (Bǎcescu, 1961) предположил близкую связь двух видов и даже в своей работе, посвящённой щиповкам Северной Италии, при характеристике S. larvata привёл на рисунке вид сбоку зрелой самки S. caspia (Fig. 2a) и вид сверху самца S. larvata (Fig. 2b). Однако в отличие от каспийской щиповки у многих особей S. larvata можно различить отдельные пятна вдоль середины бока (и нередко они не сливаются) и вдоль середины спины обыкновенно не сплошная полоса, а крупные пятна, разделённые светлыми промежутками, хорошо заметные, по меньшей мере после спинного плавника. Это видно и на иллюстрациях в ряде публикаций (Bǎcescu, 1961; Cavicchioli, Guarnieri, 1969; Lodi, 1979; Kottelat, Freyhof, 2007). (Следует отметить, что обособленные пятна вдоль середины спины имеются и у S. romanica.) Кроме этого у S. larvata в верхней части нижней губы имеется тёмное пятно (у некоторых особей из музейных коллекций сохраняется в виде затемнения), не свойственное другим видам рода Sabanejewia.
Помимо наличия полосы вдоль средней линии бока для диагностики каспийской щиповки в литературе используется ещё один признак – соотношение длины передней и задней ветвей суборбитального шипа (Берг, 1949; Казанчеев, 1981; Kottelat, Freyhof, 2007; Богуцкая и др., 2013). Действительно, у каспийской щиповки передняя ветвь шипа заметно короче задней (рис. 4), однако похожую форму шипа имеют и другие виды родов Cobitis и Sabanejewia, например, S. kubanica Vasil’eva et Vasil’ev, 1988, S. balcanica (Karaman, 1922), S. baltica Witkowski, 1994 и симпатричная каспийской золотистая щиповка S. aurata (De Filippi, 1863) (Берг, 1949; Васильева, Васильев, 1988; Witkowski, 1994; Nalbant, Bianco, 1998; Ludwig et al., 2001; настоящая работа).
В то же время, согласно полученным в настоящей работе данным, важным диагностическим признаком каспийской щиповки следует считать уникальное строение нижней губы с двумя усиковидными выростами на каждой губной лопасти. У других видов рода Sabanejewia, включая S. romanica и S. larvata, (а также у изученных видов рода Cobitis) нижняя губа иной формы: внутренний край каждой губной лопасти варьирует от гладкого или слабо складчатого до более или менее глубоко вырезанного; однако в верхней части лопасти никогда не бывает усиковидного выроста, и только нижний конец у некоторых видов может быть удлинён в виде очень короткого усика (Buşnitza, Bǎcesco, 1946; Nalbant, 1957; Bănărescu et al., 1972; Васильева, Позняк, 1986; Васильева, Васильев, 1988; Witkowski, 1994; Economidis, Nalbant, 1996; Nalbant, Bianco, 1998; настоящая работа).
В качестве дополнительных характеристик для идентификации каспийской щиповки можно использовать также относительную длину усиков, относительную длину хвостового стебля и рыла. Наиболее длинные усики среди видов рода Sabanejewia имеют S. caspia, S. kubanica, S. caucasica (Berg, 1906): у взрослых рыб мандибулярные усики доходят до заднего края глаза или далее. У S. romanica, S. aurata aralensis (Kessler, 1877), S. baltica, во многих популяциях S. balcanica усики более короткие, достигающие обычно не далее уровня середины глаза. Ещё короче усики – достигают обыкновенно не далее переднего края глаза – у S. aurata и S. larvata (Берг, 1949; Васильева, Позняк, 1986; Васильева, Васильев, 1988; Васильева, Раб, 1992; Witkowski, 1994; настоящая работа). Соответственно наблюдаются различия по длине усиков в % длины головы (табл. 2). У пяти изученных нами особей S. larvata TL 55.0–65.5 мм lB1 9.9–12.3 (в среднем 11.0)%, lB3 15.2–21.1 (18.5)%, тогда как S. romanica по этим характеристикам близка к некоторым выборкам S. balcanica: у 6 экз. TL 54.0–93.0 мм – lB1 13.1–19.3 (17.2)%, lB3 18.9–27.6 (23.0)%. Следует, однако, отметить, что длина усиков, по-видимому, в какой-то степени связана с условиями обитания видов. Ранее мы уже отмечали, что, например, у S. aurata самые короткоусые особи встречались в местах с быстрым течением и каменистым дном (Васильева, Васильев, 1988).
Таблица 2.
Некоторые морфометрические признаки разных видов рода Sabanejewia по данным литературы
| Вид | SL, мм | В % с | Источник информации | |||
|---|---|---|---|---|---|---|
| lpc | ao | lB1 | lB3 | |||
| S. balcanica: | ||||||
| – р. Лаборец (n = 13) | $\frac{{62.0{\kern 1pt} --{\kern 1pt} 84.5}}{{72.4}}$ | $\frac{{18.0{\kern 1pt} --22.1}}{{20.1 \pm 0.33}}$ | $\frac{{40.6{\kern 1pt} --{\kern 1pt} 47.4}}{{44.4 \pm 0.61}}$ | $\frac{{9.8{\kern 1pt} --{\kern 1pt} 17.4}}{{13.3 \pm 0.60}}$ | $\frac{{16.4{\kern 1pt} --{\kern 1pt} 22.4}}{{19.6 \pm 0.55}}$ | Васильева, Раб, 1992 |
| – р. Тиса (n = 30) | $\frac{{50.0{\kern 1pt} --{\kern 1pt} 76.0}}{{68.5 \pm 0.14}}$ | $\frac{{19.1{\kern 1pt} --{\kern 1pt} 23.0}}{{21.1 \pm 0.18}}$ | $\frac{{33.0{\kern 1pt} --{\kern 1pt} 46.5}}{{39.7 \pm 0.56}}$ | $\frac{{11.5{\kern 1pt} --{\kern 1pt} 18.5}}{{14.8 \pm 0.16}}$ | $\frac{{20.0{\kern 1pt} --{\kern 1pt} 28.6}}{{24.0 \pm 0.39}}$ | Мовчан, 1988 |
| – р. Днестр (n = 27) | $\frac{{51.0{\kern 1pt} --{\kern 1pt} 75.0}}{{67.0 \pm 0.14}}$ | $\frac{{18.4{\kern 1pt} --{\kern 1pt} 22.1}}{{20.2 \pm 0.18}}$ | $\frac{{37.0{\kern 1pt} --{\kern 1pt} 44.8}}{{41.4 \pm 0.43}}$ | $\frac{{10.0{\kern 1pt} --{\kern 1pt} 18.7}}{{15.5 \pm 0.44}}$ | $\frac{{13.1{\kern 1pt} --{\kern 1pt} 25.2}}{{20.5 \pm 0.57}}$ | То же |
| – р. Быстрица (n =30) | $\frac{{49.0{\kern 1pt} --{\kern 1pt} 82.0}}{{69.2 \pm 0.15}}$ | $\frac{{17.7{\kern 1pt} --{\kern 1pt} 21.3}}{{19.7 \pm 0.19}}$ | $\frac{{35.5{\kern 1pt} --{\kern 1pt} 46.7}}{{42.4 \pm 0.54}}$ | $\frac{{8.2{\kern 1pt} --{\kern 1pt} 18.4}}{{13.7 \pm 0.38}}$ | $\frac{{9.1{\kern 1pt} --{\kern 1pt} 22.1}}{{17.3 \pm 0.54}}$ | » |
| – р. Луква (n = 30) | $\frac{{58.0{\kern 1pt} --{\kern 1pt} 77.0}}{{69.7 \pm 0.06}}$ | $\frac{{17.1{\kern 1pt} --{\kern 1pt} 24.3}}{{20.2 \pm 0.20}}$ | $\frac{{42.8{\kern 1pt} --{\kern 1pt} 53.8}}{{48.4 \pm 0.28}}$ | $\frac{{14.3{\kern 1pt} --{\kern 1pt} 25.0}}{{20.3 \pm 0.50}}$ | $\frac{{22.0{\kern 1pt} --{\kern 1pt} 35.7}}{{28.4 \pm 0.16}}$ | Опалатенко, 1974 |
| S.baltica, р. Видава (n = 36) | $\frac{{50.4{\kern 1pt} --{\kern 1pt} 75.6}}{{64.5 \pm 0.64}}$ | $\frac{{19.7{\kern 1pt} --{\kern 1pt} 24.5}}{{22.2 \pm 0.20}}$ | $\frac{{35.7{\kern 1pt} --{\kern 1pt} 48.9}}{{44.1 \pm 0.53}}$ | – | – | Witkowski, 1994* |
| S. aurata, р. Кура (n = 100) | $\frac{{38.0{\kern 1pt} --{\kern 1pt} 72.0}}{{54.1 \pm 0.32}}$ | $\frac{{14.7{\kern 1pt} --{\kern 1pt} 21.1}}{{17.9 \pm 0.10}}$ | $\frac{{36.0{\kern 1pt} --{\kern 1pt} 52.8}}{{43.4 \pm 0.27}}$ | – | – | Абдурахманов, 1962 |
| S. kubanica, р. Невинка (n = 17) | $\frac{{48.3{\kern 1pt} --{\kern 1pt} 65.2}}{{57.2}}$ | $\frac{{18.3{\kern 1pt} --{\kern 1pt} 23.1}}{{20.1 \pm 0.28}}$ | $\frac{{41.2{\kern 1pt} --{\kern 1pt} 51.1}}{{45.0 \pm 0.66}}$ | $\frac{{13.9{\kern 1pt} --{\kern 1pt} 27.1}}{{19.9 \pm 0.71}}$ | $\frac{{24.2{\kern 1pt} --{\kern 1pt} 36.3}}{{29.2 \pm 0.73}}$ | Васильева, Васильев, 1988 |
| S. caucasica: | ||||||
| – р. Сунжа (n = 15) | $\frac{{58.5{\kern 1pt} --{\kern 1pt} 77.6}}{{67.7}}$ | $\frac{{16.8{\kern 1pt} --{\kern 1pt} 22.02}}{{19.5 \pm 0.4}}$ | $\frac{{42.0{\kern 1pt} --{\kern 1pt} 51.3}}{{48.6 \pm 0.61}}$ | $\frac{{18.8{\kern 1pt} --{\kern 1pt} 29.0}}{{23.4 \pm 0.80}}$ | $\frac{{25.5{\kern 1pt} --{\kern 1pt} 34.3}}{{29.5 \pm 0.73}}$ | Васильева, Позняк, 1986 |
| – р. Хайдон (n = 34) | $\frac{{44.0{\kern 1pt} --{\kern 1pt} 91.2}}{{76.4}}$ | $\frac{{16.9{\kern 1pt} --{\kern 1pt} 28.0}}{{19.0 \pm 0.21}}$ | $\frac{{43.9{\kern 1pt} --{\kern 1pt} 51.8}}{{47.0 \pm 0.29}}$ | $\frac{{16.7{\kern 1pt} --{\kern 1pt} 28.9}}{{24.3 \pm 0.80}}$ | $\frac{{21.7{\kern 1pt} --{\kern 1pt} 35.7}}{{29.6 \pm 1.39}}$ | То же |
По относительной длине хвостового стебля все виды рода Sabanejewia достаточно чётко подразделяются на две группы. К видам с коротким хвостовым стеблем относятся всего два: каспийская щиповка (табл. 1) и S. larvata: у пяти изученных особей TL 55.0–65.5 мм lpc 14.5–16.7 (15.5)%; а рассчитанные нами значения lpc по данным литературы (Cavicchioli, Guarnieri, 1969) составили 14.7 и 15.4%. По lpc наблюдается хиатус между выборками этих двух видов и выборками S. baltica, S. kubanica Vasil’eva et Vasil’ev, 1988 и большинством выборок S. balcanica (табл. 2). В остальных случаях диапазоны варьирования выборочных значений каспийской щиповки и S. larvata лишь незначительно перекрываются с диапазонами значений других видов рода, при этом средние значения видов с коротким хвостовым стеблем, как правило, не заходят за нижнюю границу пределов варьирования признака у остальных видов, а оценки показателя СD превышают формально подвидовой уровень. Помимо видов, указанных в табл. 2, к группе щиповок с длинным хвостовым стеблем относятся S. romanica (у 6 экз. TL 54.0–93.0 мм lpc 17.6–19.5 (18.5)%), S. vallachica (Nalbant, 1957) (по данным Nalbant, 1957 – 16.75–21.50 (18.50)%) и S. bulgarica (Drensky, 1928) (по данным Nalbant, 1957 – 16.30–20.25 (17.65)%).
В меньшей степени выражены межвидовые различия у щиповок рода Sabanejewia по длине рыла. Тем не менее наиболее короткорылая каспийская щиповка из Малого Кызылагачского залива отличается от большинства длиннорылых видов с хиатусом или при незначительном перекрывании диапазонов значений признака ao (табл. 1, 2). У щиповок из р. Кумбаши рыло длиннее, как у S. balcanica р. Тиса, близкие значения признака получены нами для S. larvata – 35.9–42.9 (38.8)%. Все они достоверно отличаются от длиннорылых видов со средними значениями ао 41.4–48.6% (табл. 2), к которым относится и S. romanica – 41.4–44.7 (42.3)%.
Таким образом, на основе проведённого сравнительного анализа видовой диагноз каспийской щиповки должен быть расширен и представлен в следующей редакции, не допускающей неоднозначную трактовку: вдоль средней линии бока проходит непрерывная продольная тёмная полоса, образованная мелкими пигментными крапинками, сплошная тёмная полоса проходит вдоль середины спины, часто имеется менее чёткая тёмная продольная полоса из мелких крапинок ниже середины бока; крупных пятен на спине и на боках тела нет (на спине могут быть только у молоди); на внутренней стороне каждой лопасти нижней губы по два усиковидных выроста: нижний образован удлинённым окончанием внутренней стороны губы, а верхний – удлинением её верхней складки; передняя ветвь суборбитального шипа заметно короче задней; усики длинные: мандибулярные у взрослых особей, как правило, заходят за задний край глаза; хвостовой стебель короткий: содержится обычно более 6 раз в SL (<18% SL); рыло короткое, обычно составляет <40% с.
Половой диморфизм по внешним признакам. Для взрослых самцов многих видов рода Sabanejewia отмечается вздутие боков тела перед спинным и брюшными плавниками, такой тип полового диморфизма рассматривается в качестве одной из важных отличительных характеристик рода (Vladykov, 1929; Берг, 1949). Однако у разных видов половые различия выражены в разной степени. Так, по наличию вздутий зрелые самцы хорошо отличаются от самок у S. larvata, S. bulgarica, S. vallachica, S. kubanica, S. romanica, S. caucasica, в некоторых популяциях S. balcanica и S. aurata (Buşnitza, Bǎcesco, 1946; Nalbant, 1957; Bǎcescu, 1961; Васильева, Позняк, 1986; Васильева, Васильев, 1988; настоящая работа). При этом соотношение полов в выборках при определении по внешнему виду особей оказывается близким к 1 : 1. В то же время в изученной ранее популяции S. balcanica из р. Лаборец вздутия у самцов были выражены очень нечётко: примерно в половине случаев идентификация самцов по этому признаку не совпала с определением пола на основе вскрытия (Васильева, Раб, 1992).
У каспийской щиповки половой диморфизм выражен очень слабо. В пробе, собранной 20–21.04.1982 г. в Малом Кызылагачском заливе близ пос. Нариманабад и в протоке, которая соединяет залив с морем, включающей 51 экз., только у одного самца (TL 69 мм) наблюдались хорошо выраженные боковые вздутия тела впереди спинного плавника (рис. 5). Другие самцы, пол которых был выявлен по результатам вскрытия, по внешним признакам визуально от самок не отличались. При этом длина тела самцов (TL) варьировала от 55 до 74 мм (самок – от 54 до 78 мм), более 30% имели гонады IV стадии зрелости (среди самок преобладали рыбы с яичниками IV–V стадий, у единичных особей (8.6%) наблюдалась остаточная икра). В музейной пробе Р-15917 (собрана в заливе в те же сроки) у 31 экз. (из 36) TL 56.5–72.5 мм явственных вздутий на боках тела не обнаружено. У одной особи TL 62 мм хорошо заметны вздутия в нижней части боков, сразу под тёмной полосой вдоль средней линии тела; эти вздутия начинаются чуть впереди окончания грудных плавников и заканчиваются на уровне начала спинного и брюшных плавников, вздутия плотные. У трёх особей TL 57, 61 и 69 мм нижняя часть боков перед началом спинного и брюшных плавников слабо выпуклая, вздутия мягкие; ещё у 1 экз. TL 62 мм хорошо заметные плотные вздутия в нижней части боков начинаются примерно от одной трети грудного плавника и заканчиваются чуть далее конца грудного плавника. В пробе Р-16756 (собрана в заливе в октябре) у 103 экз. (из 111) TL 47–81 мм (преобладали рыбы TL < 6 см) никаких следов вздутий на боках тела не обнаружено. У 5 экз. TL 55–62 мм перед спинным плавником чуть просматривается слабое расширение тела с боков по сравнению с шириной на уровне плавника; расширение боков перед спинным плавником заметно со спины еще у 2 экз. TL 62 и 64 мм. У 1 экз. TL 58 мм заметны вздутия за головой (доходят до конца грудных плавников), у 1 экз. TL 63 мм вздуто все брюхо. В пробе Р-3718 (Джильское озеро, апрель) у 6 экз. TL 43.0–59.5 мм нет следов вздутий на боках, но у некоторых утолщено брюхо (с икрой?), очень слабые вздутия боков у двух рыб TL 57 и 58 мм. В пробе Р-16755 (ахмаз в бассейне Куры, 28 мая) у 30 взрослых рыб TL 49–57 мм вздутий нет, у 9 экз. TL 52–60 мм очень слабые вздутия на боках тела. В пробе Р-16803 (каналы в районе Мингечаура, 16 мая) слабые расширения верхней части боков перед спинным плавником обнаружены у двух взрослых особей TL 49–53 мм, у других двух TL 49 и 52 мм вздутия не выражены. В пробе Р-3717 (р. Хирмандали, июнь) у двух взрослых особей TL 56.5 и 61.0 мм нет вздутий, у 3 экз. TL 57–62 мм очень слабые вздутия. В пробе Р-3724 (Джильское озеро, июнь) у самого крупного из 3 экз. (TL 58–68 мм) намечается очень слабое расширение перед спинным плавником. Не обнаружено вздутий на боках тела у всех шести взрослых особей TL 57–63 мм из пробы Р-3721 (низовья Сефидруда, июнь), у всех 5 экз. TL 67–75 мм в пробе Р-3719 (Джильское озеро, сентябрь), в пробе Р-16747 (ахмаз в бассейне Куры, 16 мая, 2 экз. TL 52 и 71 мм), в пробе Р-3744 (Джильское озеро, июнь, 2 экз. TL 68 и 74 мм, у мелкой особи вздутое брюхо – с икрой?), в пробах из р. Кумбаши Р-3720 (июнь, 1 экз. TL 64 мм) и Р-3723 (апрель, 1 экз. TL 58 мм – самка с икрой?).
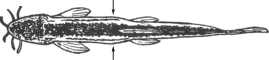
Рис. 5.
Самец Sabanejewia caspia TL 69 мм из Малого Кызылагачского залива с выраженными вздутиями на боках тела (↑), вид сверху.
При анализе полового диморфизма мы руководствовались данными Абдурахманова (1962), согласно которым самцы и самки каспийской щиповки становятся половозрелыми после достижения длины (SL) 35 мм (соответственно TL 39 мм) и нерестятся, по-видимому, в апреле. Во всяком случае 18.04.1953 г. все изученные им рыбы SL 35–65 мм (преобладали 35–40 мм) были с развитыми половыми продуктами (была посчитана плодовитость). Среди самок щиповок, отловленных нами в Малом Кызылагачском заливе 20–21.04.1982 г. (SL > 45 мм), преобладали текучие особи с гонадами V стадии зрелости, встречались самки с яичниками VI–II стадии, более 30% самцов также были текучими, что подтверждает апрельский нерест щиповки в Ленкоранском районе Азербайджана. В разных музейных пробах, собранных в апреле, с относительно высокой численностью рыб соотношение полов у взрослых особей при определении по наличию вздутий на боках варьирует от 1 : 3 (Р-3718) до 1 : 6.2 (Р-15917) в пользу самок, тогда как у вскрытых рыб, отловленных 20–21.04.1982 г., соотношение полов было 23 : 28 в пользу самцов. Среди рыб, собранных 09.05.1985 г. в Малом Кызылагачском заливе для кариологического анализа (110 экз.), соотношение полов у вскрытых особей было 1 : 2.2 в пользу самок, в это время среди самок преобладали особи с остаточной икрой (VI стадия). В то же время в музейной пробе, собранной в конце мая (Р-16755), соотношение полов по наличию вздутий – 1 : 3.3 в пользу самок, а в пробе, собранной в октябре (Р-16756), – 1 : 13. Эти результаты свидетельствуют не в пользу реального соотношения полов и разных биотопических предпочтений самцов и самок, а в пользу слабо выраженного полового диморфизма по внешним признакам даже в нерестовый период.
Рассматривая экологические особенности каспийской щиповки, следует также отметить, что у этого вида икра визуально заметно мельче, чем у симпатричных представителей рода Cobitis, что согласуется с данными Абдурахманова (1962), который приводит для мелких икринок самок р. Кумбашинка средний диаметр 0.33 мм, для средних – 0.45, а для крупных – 0.85 мм. Тогда как у изученных им самок рода Cobitis из р. Союгбулаг (приток Куры) средний диаметр мелких икринок достигал 0.78 мм, а крупных – 1.50 мм. У всех изученных нами особей каспийской щиповки из Кызылагачского залива икра была не желтоватого, как у других щиповок, а зелёного цвета.
Краниологическая характеристика каспийской щиповки. По общему строению черепа и числу костей каспийская щиповка принципиально не отличается от изученных ранее видов родов Sabanejewia и Cobitis (Васильева, 1984; Васильева, Позняк, 1986; Васильева, Васильев, 1988), однако обнаруживает заметную дивергенцию в пропорциях ряда костных структур. Череп у каспийской щиповки невысокий и относительно узкий, теменное отверстие и отверстия на боковых затылочных костях маленькие; operculum и praeoperculum относительно широкие, suboperculum сужается к заднему концу; высота praemaxillare, как правило, ниже его длины; dentale очень низкое, его восходящий отросток несколько сдвинут в переднюю часть кости; hyomandibulare и cleithrum узкие, окончание передней ветви суборбитального шипа далеко не доходит до конца задней ветви (рис. 2, табл. 3). На левой глоточной кости обнаружено от 6 до 12 зубов, модальное число зубов лежит в интервале 8–10 (44%), на правой кости – от 6 до 13 зубов, чаще всего встречалось 9 (20%) или 12 (25%) зубов. У самцов по сравнению с самками достоверно больше средние значения Lf1, wCl, WCl и меньше hD, однако различия эти невелики.
Таблица 3.
Некоторые краниологические признаки каспийской щиповки Sabanejewia caspia из Малого Кызылагачского залива
| Признак | Самки (n = 14) | Самцы (n = 12) | ||
|---|---|---|---|---|
| lim | M ± m | lim | M ± m | |
| TL, мм | 54.0–78.0 | 67.5 ± 1.95 | 50.0–74.0 | 65.1 ± 1.84 |
| В % длины основания черепа | ||||
| Hmax | 26.8–31.8 | 29.5 ± 0.37* | 24.4–35.4 | 31.1 ± 0.81* |
| Heth | 15.5–21.7 | 17.9 ± 0.58* | 14.4–22.8 | 18.3 ± 0.69* |
| sSph | 25.6–34.1 | 30.2 ± 0.70* | 27.8–35.7 | 31.4 ± 0.83 |
| sPt | 33.3–39.8 | 35.5 ± 0.54* | 33.3–41.4 | 37.4 ± 0.68* |
| sEth | 20.2–28.2 | 23.2 ± 0.56* | 21.7–25.3 | 23.5 ± 0.36 |
| Lf1 | 14.0–22.7 | 18.4 ± 0.74* | 19.5–27.4 | 22.2 ± 0.67 |
| Lf2 | 2.3–8.9 | 6.7 ± 0.52* | 3.3–8.9 | 7.0 ± 0.49 |
| В % длины кости | ||||
| hOp | 43.9–56.7 | 48.8 ± 0.92 | 43.1–52.9 | 48.6 ± 0.82 |
| lOp | 75.4–90.1 | 83.8 ± 1.20 | 75.8–85.7 | 81.3 ± 0.73 |
| h1/h2Iop | 52.6–73.7 | 63.6 ± 1.87 | 44.4–76.8 | 59.4 ± 2.93 |
| wPop | 10.1–20.1 | 15.6 ± 0.66 | 14.1–19.9 | 16.7 ± 0.54 |
| rPop | 16.2–25.0 | 19.5 ± 0.81* | 18.1–26.1 | 22.4 ± 1.13* |
| hPm | 78.6–115.3 | 90.9 ± 2.75* | 79.8–98.4 | 87.7 ± 1.93 |
| hDent | 41.3–51.8 | 45.4 ± 0.86* | 39.0–46.7 | 43.2 ± 0.61 |
| rDent | 66.7–80.4 | 73.3 ± 1.42* | 63.7–77.1 | 70.7 ± 1.46 |
| wH | 33.7–42.8 | 39.0 ± 0.74* | 30.8–43.1 | 39.3 ± 0.98 |
| lH | 48.0–65.9 | 57.2 ± 1.47* | 52.6–64.4 | 57.7 ± 1.20 |
| h1/h2Mx | 52.6–93.6 | 75.9 ± 2.51 | 58.6–87.9 | 75.0 ± 2.46 |
| b/aSorb | 31.9–68.0 | 54.1 ± 3.40 | 45.0–72.9 | 57.1 ± 2.65 |
| lSorb | 41.0–86.4 | 72.5 ± 2.92* | 53.9–78.7 | 72.8 ± 1.92 |
| sSorb | 14.8–23.9 | 20.5 ± 1.15* | 16.5–24.3 | 20.1 ± 0.81 |
| wCl | 7.3–10.4 | 9.1 ± 0.21 | 8.3–10.8 | 9.8 ± 0.23 |
| WCl | 15.9–20.2 | 18.3 ± 0.40 | 16.7–22.9 | 20.3 ± 0.56 |
Примечание. lim − пределы изменчивости, M ± m – среднее значение и его ошибка, n – число исследованных особей, экз.; TL – общая длина тела; Hmax − максимальная высота черепа, Heth − высота его передней части, sSph – ширина черепа на уровне sphenotica, sPt – то же на уровне pterotica, sEth – то же на уровне ethmoidalia lateralia, Lf1 − длина теменного отверстия, Lf2 − длина отверстия на occipitale laterale; hOp, lOp − высота и длина переднего края operculum, h1/h2Iop − отношение высоты передней части interoperculum к высоте задней части; wPop, rPop − ширина и расстояние до отростка praeoperculum; hPm − высота praemaxillare, hDent, rDent – высота и расстояние до конца отростка dentale; wH − ширина hyomandibulare, lH − длина его передней части, h1/h2Mx − отношение высоты отростка maxillare к высоте кости, b/aSorb − отношение расстояния до бокового выроста к расстоянию от выроста до конца передней ветви суборбитального шипа, lSorb − расстояние до вершины передней ветви, sSorb − глубина вырезки у основания передней ветви суборбитального шипа, wCl − ширина верхней части cleithrum, WCl – его максимальная ширина; *число исследованных особей меньше n.
Среди изученных ранее видов рода Sabanejewia от каспийской щиповки по краниометрическим индексам в наибольшей степени отличается S. caucasica: в общей сложности достоверные различия выявлены по 16 признакам; по шести из них различия достигают значительной величины: CD превышает формально подвидовой уровень 1.28 между самками по индексам Heth (1.34), hDent (1.49), WCl (2.55), между самцами – wPop (1.39), hPm (1.39), hDent (3.80), lSorb (1.90), WCl (2.85). Диапазоны изменчивости двух видов по этим характеристикам перекрываются слабо, а в некоторых случаях наблюдается хиатус (рис. 6). В меньшей степени выражена дивергенция между каспийской щиповкой и S. kubanica. Достоверные различия между видами выявлены по 13 индексам, по пяти из них CD превышает 1.28 (сравнивались только самки, поскольку выборка самцов кубанской щиповки нерепрезентативна): rPop, hDent, wH, wCl, WCl (CD соответственно 1.53, 1.43, 2.35, 1.27, 2.42), хиатус наблюдается по двум признакам. Ещё ниже уровень дивергенции между каспийской щиповкой и S. aurata aralensis: достоверные различия выявлены по 11 индексам, по четырём из них CD превышает уровень 1.28 при сравнении суммарной выборки аральской щиповки с самками каспийской (hDent – 1.44, wH – 1.53, wCl – 1.38, WCl – 1.89) и с самцами (hDent – 2.01, rDent – 1.37, wH – 1.29, WCl – 1.27).
Рис. 6.
Пределы изменчивости (—) и диапазон средних значений (◼) некоторых краниологических индексов у разных видов щиповок рода Sabanejewia: 1 – S. caspia, 2 – S. aurata aralensis, 3 – S. kubanica, 4 – S. caucasica; lSorb − расстояние до вершины передней ветви, wH − ширина hyomandibulare, hDent − высота dentale, hPm − высота praemaxillare, Heth − высота передней части черепа, wCl − ширина верхней части cleithrum, WCl – его максимальная ширина, rDent − расстояние до конца отростка dentale, wPop − ширина praeoperculum.
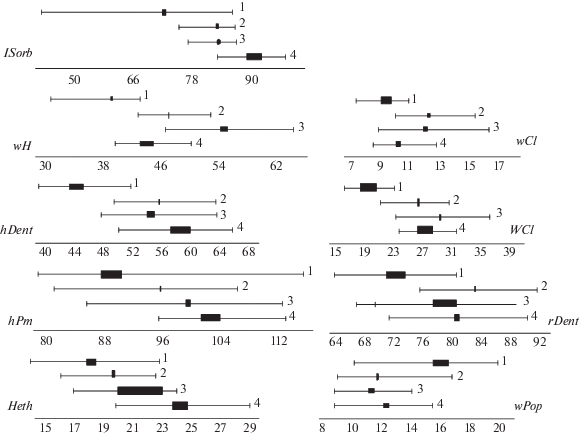
Выявленные краниологические различия свидетельствуют о высоком уровне дивергенции каспийской щиповки от других видов рода. Ранее нами было проведено сравнение по краниологическим индексам аральской и кубанской щиповок (Васильева, Васильев, 1988). Большинство авторов считают аральскую щиповку синонимом S. aurata (Froese, Pauly, 2017; Eschmeyer et al., 2018), самостоятельный видовой статус кубанской щиповки, исходно описанной в качестве подвида S. aurata, подтверждён генетическими исследованиями (Perdices et al., 2003). Между изученными выборками этих двух видов хотя и были выявлены достоверные различия по восьми краниометрическим признакам, однако уровень этих различий невысок: CD < 1.28, поэтому формально их можно рассматривать не как межвидовые, а как межпопуляционные. Слабая дивергенция по краниологическим признакам наблюдается и между некоторыми видами рода Cobitis (Васильева, Васильев, 1988).
Структура кариотипа каспийской щиповки. Кариотип каспийской щиповки включает в диплоидном наборе 50 хромосом, из которых две пары метацентрические, 9 пар субмета- и 15 пар субтело- и акроцентрические (рис. 7а). Характерные особенности кариотипа: 1) длина четырёх метацентрических хромосом относительно небольшая, эти хромосомы заметно меньше или равны длине больших плеч первых субметацентрических хромосом; 2) все хромосомы заметно меньше, чем у других видов родов Sabanejewia и Cobitis. В последнем роде только C. melanoleuca Nichols, 1925 имеет заметно меньшие размеры хромосом, чем другие виды рода. Измерение количества ДНК на клетку показало, что у этого вида содержание ДНК на клетку примерно на 20% меньше, чем у других видов рода Cobitis (Васильев и др., 1999). Важно отметить, это указывалось нами и ранее (Васильева, Васильев, 1988), что среди изученных видов рода Sabanejewia подобная структура кариотипа характерна только для S. larvata (Lodi, Marchionni, 1980). Число и морфологию метацентрических хромосом можно рассматривать в качестве маркерных признаков для S. caspia и S. larvata. Для сравнения на рис. 7б представлен изученный нами кариотип S. aurata из р. Куры (близ Ахалдабо) с тремя парами метацентрических хромосом. Среди других видов рода Sabanejewia две пары метацентрических хромосом имеет только S. balcanica из р. Лаборец (бассейн Тисы, Словакия) (Васильева, Раб, 1992), но это не мелкие, как у S. caspia, а достаточно крупные (особенно первая пара) хромосомы. Более детальный сравнительный анализ кариотипов по раскладкам хромосом S. caspia (рис. 7а) и S. larvata (Lodi, Marchionni, 1980) показывает, что эти виды отличаются по числу субметацентрических хромосом: у первого вида таких хромосом деять пар, у второго – восемь. Таким образом, в кариологическом отношении два вида, S. caspia и S. larvata, образуют компактную и обособленную от других видов группу в роде Sabanejewia.
Рис. 7.
Кариотип каспийской щиповки Sabanejewia caspia из Малого Кызылагачского залива (а) и S. aurata из р. Кура (б): m – мета-, sm – субмета-, sta – субтело- и акроцентрические хромосомы.

Распространение каспийской щиповки. Ранее (Васильева, 1995) уже было показано, что все указания на обитание каспийской щиповки в северной части Каспийского моря (Берг, 1949; Абдурахманов, 1962; Казанчеев, 1981) основаны на ошибочной видовой идентификации. Подтверждённым музейными коллекциями и опубликованными исследованиями следует считать ареал вида в бассейне Юго-Западного и Южного Каспия от низовьев Куры до Баболя (= Бабуля) (Державин, 1934; Абдурахманов, 1962; Васильева, 1995; Coad, 2017; настоящая работа). Кесслер (1877) считал каспийскую щиповку солоноватоводным видом и уверенно отмечал её лишь в Каспийском море, ставя под вопрос наличие в Куре. Абдурахманов (1962), наоборот, считал, что это пресноводная рыбка, обитает в стоячих и медленно текучих водах с водяной растительностью – в придаточных водоёмах нижнего течения Куры (озёра Шильян, Аджикабул, Ахчала), в р. Кумбашинка, впадающей в Малый Кызылагачский залив. К пресным водоёмам относятся все пробы каспийкой щиповки из Ирана с конкретными локалитетами (Coad, 2017) и все пробы в коллекции ЗММУ (Малый Кызылагачский залив, хотя и имеет морское происхождение, давно отделён от моря искусственной дамбой и фактически превращён в замкнутый пресноводный водоём).
Филогенетические связи каспийской щиповки впервые на основе классических морфологических исследований попытался оценить Баческу (Băcescu, 1943), описав, как уже отмечалось выше, в статусе её подвида щиповку из Румынии – ныне самостоятельный вид S. romanica. В то же время Бэнэреску (Bǎnǎrescu, 1989, 1990) считал близкородственным S. caspia североитальянский вид S. larvata, приводя эту пару видов в качестве примера географического викаризма. В качестве сестринских видов рассматривали их также Налбант и Бьянко (Nalbant, Bianco, 1998). Проведённый в настоящей работе сравнительный анализ морфологических характеристик этих трёх видов не даёт оснований для их сближения, в то же время кариологические данные свидетельствуют в пользу филогенетического родства S. caspia и S. larvata.
Первая (и единственная) попытка оценки филогенетических связей каспийской щиповки с использованием молекулярно-генетического анализа была осуществлена, как уже отмечалось выше, при полном отсутствии материалов для исследования этого вида (Perdices et al., 2003). Отсутствие данных по каспийской щиповке, чьё распространение существенно расширяет современный ареал Sabanejewia в азиатском направлении, позволяет сомневаться не только в корректности оценки филетических отношений в пределах рода в целом, но и выделения базальных линий и постулирования “монофилетичности” конгломерата из “балтийского, кавказского и каспийских видов” (Perdices et al., 2016. Р. 11). В состав этой монофилетичной группы вошли признаваемые авторами в статусе самостоятельных видов S. baltica, S. kubanica, S. aurata, а также предкавказская щиповка S. caucasica – ещё один вид рода, с которым связаны досадные ошибки в идентификации, приводящие к неверной оценке видового ареала, на которых мы считаем необходимым остановиться в настоящей работе.
Описание S. caucasica было составлено по экземпляру ЗИН 8560, для которого указан локалитет “Saagdan, Gr. Laba-Fluss, oberer Kuban-FL, Kubansk-Gebiet, N. Kaukasus” (Berg, 1906. Р. 37). Соответственно этой этикетке он был собран в р. Сагдан (= Загедан) – притоке верхнего течения Большой Лабы, системы р. Кубань (бассейн Чёрного моря). Кроме этого в описании упомянуты шесть рыб, полученных из ЗММУ, которых ранее Берг (1899) определил как “C. aurata”. Это щиповки из рек Челдырка, Камбилеевка, Аргун, относящихся к бассейну Терека. Берг (Berg, 1906) отмечает, что эти особи не отличаются от экземпляра ЗИН 8560, по одной из них был сделан рисунок в первоописании. Таким образом, типовой локалитет предкавказской щиповки охватывает бассейны как Чёрного, так и Каспийского морей, а типовая серия должна включать 2 экз., один из которых теоретически может быть опознан по рисунку (сделанному в натуральную величину) из 6 рыб, присланных из ЗММУ22. На упомянутом рисунке чётко представлена характерная для S. caucasica окраска (Берг, 1949. С. 896–897): “тело покрыто многочисленными бурыми пятнышками. Больших чёрных пятен вдоль середины боков нет (выделение наше. – Авторы)”. Такая окраска не обнаружена до сих пор ни у какого другого вида щиповок данного рода и является основным диагностическим признаком предкавказской щиповки, в ареал которой Берг (1949) включал реки Каспийского бассейна (Кума, Терек, Сулак, Шура-озень) и бассейн верхнего течения Кубани. На основании музейных экземпляров и собственных сборов мы подтвердили распространение вида в бассейнах Терека (приток Сунжи, р. Хайдон, р. Челдырка) и Кумы (р. Подкумок и отводные каналы р. Золка) и дали подробную морфологическую характеристику вида, включая особенности строения костей черепа, и ряд сведений по экологии (Васильева, Позняк, 1986). Все заключения работы сделаны на основе конкретных проб ЗММУ: Р-2244, Р-2850, Р-15908–Р-15910. В то же время во всех последующих публикациях (фаунистических или краснокнижных списках) ни одно из упоминаний предкавказской щиповки не обосновывается какими-либо фактическими данными. Более того, в популярной монографии Коттеля и Фрейхофа (Kottelat, Freyhof, 2007), где ареал предкавказской щиповки фантастически расширился, полностью захватив западное и южное побережья Каспийского моря (от Кизлярского залива до ирано-туркменской границы), на фотографии “Sabanejewia caucasica, Самур” (P. 325, вторая фотография сверху, фото А. Насеки) на самом деле изображён вид S. aurata с характерным для него рядом крупных пятен вдоль середины бока и с тёмной полосой мелких крапинок, идущих от рыла через глаз, также не свойственной S. caucasica. Эта же фотография приведена для S. caucasica и Богуцкой с соавторами (2016. Фото 16). И более того, что особенно удручает, она долгое время была размещена на популярном сайте FishBase (Froese, Pauly, 2017), вводя в заблуждение широкие круги пользователей.
До этой фотографии никаких упоминаний о наличии предкавказской щиповки в р. Самур (как и в других водоёмах к югу от Сулака) не было. Берг (Berg, 1906) подчёркивал, что изученные им особи данного вида не относятся к бассейну Куры; на необходимость подтверждения конкретным материалом включения Ирана в ареал вида указывает и Коэд (Coad, 2017). В этой связи следует подчеркнуть, что ранее мы отмечали сходство биотопов, занимаемых S. caucasica и S. aurata, случаи совместного обитания которых неизвестны (Васильева, Позняк, 1986), а в бассейне р. Кура и в водоёмах Ирана подтверждено распространение S. aurata. После публикации фотографии А. Насеки Самур также следует включить в ареал S. aurata.
В связи с изложенным выше и полученными Пердисес с соавторами (Perdices et al., 2003) результатами правильность отнесения к виду S. caucasica двух экземпляров щиповок, чьи гаплотипы обозначены исходно как 257, 260 MO и затем помещены в Генбанк под № АУ059338 и АУ059339, вызывает серьёзные сомнения. На филогенетических деревьях, построенных на основе данных по изменчивости митохондриального генома, в пределах общей клады S. aurata–S. caucasica–S. baltica–S. kubanica гаплотипы предкавказской щиповки объединяются в общую группу с гаплотипами золотистой щиповки из бассейна Каспия и р. Риони, в то время как S. baltica и S. kubanica образуют две самостоятельных подклады с высоким уровнем поддержки. Этот результат свидетельствует в пользу принадлежности изученных проб АУ059338 и АУ059339 к виду S. aurata, к сожалению, ваучерные экземпляры, по-видимому, не сохранились. Поскольку упомянутые гаплотипы достаточно часто используются в филогенетических реконструкцией вьюновых в качестве репера S. caucasica (Perdices et al., 2003, 2016; Bartoňová et al., 2008; Buj et al., 2008), несомненно, следует повторно изучить генотип данного вида на основе фиксированных ваучерных особей, доступных для верификации. Совершенно необходимы также генетические исследования каспийской щиповки с фиксацией ваучерных экземпляров в музейных коллекциях. А пока, до проведения таких исследований, к сожалению, следует констатировать, что филогенетические связи S. caspia, как и филогенетические отношения всех видов рода Sabanejewia, остаются дискуссионными.
Таксономические исследования рыб Понто-Каспия проводятся при финансовой поддержке Российского фонда фундаментальных исследований (проект № 16-04-00130), а изучение биоразнообразия и обработка депонированных музейных коллекций – при поддержке Российского научного фонда (проект № 14-50-00029) в рамках гостемы АААА-А16-116021660077-3.
Список литературы
Абдурахманов Ю.А. 1962. Рыбы пресных вод Азербайджана. Баку: Изд-во АН АзССР, 407 с.
Берг Л.С. 1899. Данные по ихтиофауне Кавказа // Изв. Кавказ. музея. Т. 1. Вып. 3. С. 1–36.
Берг Л.С. 1916. Рыбы пресных вод Российской империи. М.: Тип. т-ва Рябушинских, 563 с.
Берг Л.С. 1949. Рыбы пресных вод СССР и сопредельных стран. Ч. 2. М.; Л.: Изд-во АН СССР. С. 469–926.
Богуцкая Н.Г., Кияшко П.В., Насека А.М., Орлова М.И. 2013. Определитель рыб и беспозвоночных Каспийского моря. Т. 1. Рыбы и моллюски. СПб.; М.: Т-во науч. изд. КМК, 543 с.
Васильев В.П. 1978. Хромосомный полиморфизм у смариды – Spicara smaris (Pisces, Centracanthidae) // Зоол. журн. Т. 57. № 8. С. 1276–1278.
Васильев В.П. 1985. Эволюционная кариология рыб. М.: Наука, 300 с.
Васильев В.П., Виноградов А.Е., Розанов Ю.М., Васильева Е.Д. 1999. Содержание ДНК в клетках форм однополо-двуполого комплекса щиповок рода Cobitis и щиповки Лютера C. lutheri (Cobitidae) // Вопр. ихтиологии. Т. 39. № 3. С. 355–361.
Васильева Е.Д. 1984. Сравнительный морфологический анализ двух популяций щиповок (род Cobitis, Cobitidae), отличающихся числом пятен у основания хвостового плавника // Там же. Т. 24. Вып. 1. С. 43–53.
Васильева Е.Д. 1995. Об отсутствии каспийской щиповки Sabanejewia caspia (Cobitidae) в бассейне северного Каспия // Там же. Т. 34. № 6. С. 839–840.
Васильева Е.Д., Васильев В.П. 1988. Исследование внутривидовой структуры Sabanejewia aurata (Cobitidae) с описанием нового подвида S. aurata kubanica subsp. nov. // Там же. Т. 28. Вып. 2. С. 192–212.
Васильева Е.Д., Васильев В.П. 2012. Cobitis amphilekta sp. nova – новый вид щиповки (Cobitidae, Cypriniformes) из бассейна Каспийского моря // Там же. Т. 52. № 2. С. 177–183.
Васильева Е.Д., Позняк В.Г. 1986. Морфологическая характеристика предкавказской щиповки Sabanejewia caucasica (Berg) (Cobitidae) // Там же. Т. 26. Вып. 3. С. 402–409.
Васильева Е.Д., Раб П. 1992. Золотистая щиповка Sabanejewia aurata (Cobitidae) реки Лаборец // Там же. Т. 32. Вып. 2. С. 176–181.
Державин А.Н. 1934. Пресноводные рыбы южного побережья Каспия // Тр. Аз. отд. Закавказ. филиала АН СССР. Сект. зоология. Т. 7. С. 91–126.
Казанчеев Е.Н. 1981. Рыбы Каспийского моря. М.: Лег. и пищ. пром-сть, 167 с.
Кесслер К. 1877. Рыбы, водящиеся и встречающиеся в Арало-каспийско-понтийской области // Тр. Арало-Касп. эксп. Вып. 4. 360 с.
Мовчан Ю.В. 1988. Вьюновые, сомовые, икталуровые, пресноводные угри, конгеровые, саргановые, треcковые, колюшковые, игловые, гамбузиевые, зеусовые, сфиреновые, кефалевые, атериновые, ошибневые // Фауна Украины. Т. 8. Рыбы. Вып. 3. Киев: Наук. думка, 367 с.
Опалатенко Л.К. 1974. К морфобиологической характеристике вьюновых (Cobitidae) верхнего Днестра // Вестн. зоологии. № 6. С. 56–62.
Băcescu M.C. 1943. Deux poissons nouveaux pour la faune de la Roumanie: Cobitis aurata balcanica Karaman et Cobitis caspia romanica n. spp. // Bull. Secti. Sci. Acad. Roumaine. Bucaresti. V. 26. № 2. P. 133–141.
Băcescu M. 1961. Contributo allo studio dei Cobitis dell’Italia settentrionale // Estrato dall’archivio di oceanografia e limnologia. V. 12. F. 2. P. 185–189.
Bănărescu P.M. 1989. Vicariant patterns and dispersal in European freshwater fishes // Spixiana. V. 12. № 1. P. 91–103.
Bănărescu P. 1990. Zoogeography of fresh waters. V. 1. General distribution and dispersal of freshwater animals. Wiesbaden: Aula-Verlag, 511 p.
Bănărescu P., Nalbant T.T., Chelmu S. 1972. Revision and geographical variation of Sabanejewia aurata in Romania and the origin of S. bulgarica and S. romanica (Pisces, Cobitidae) // Annot. Zool. Botan. Bratislava. V. 75. P. 1–49.
Bartoňová E., Papoušek I., Lusková V. et al. 2008. Genetic diversity and taxonomy of Sabanejewia balcanica (Osteichthyes: Cobitidae) in the waters of the Czech Republic and Slovakia // Folia Zool. V. 57. № 1–2. P. 60–70.
Berg L.S. 1906. Beschreibung einiger kaukasischer Fische // Bull. Acad. Imp. Sci. St. Pétersbourg. Sér. 5. V. 24. № 1–2. P. 35–39.
Buj I., Podnar M., Mrakovčić M. et al. 2008. Morphological and genetic diversity of Sabanejewia balcanica in Croatia // Folia Zool. V. 57. № 1–2. P. 100–110.
Buşnitza Th., Băcesco M. 1946. Cobitis bulgarica Drensky un poissons nouveau pour la faune de la Roumanie // Notationes Biollogicae. V. 4. № 1–3. P. 125–135.
Cavicchioli G., Guarnieri P. 1969. Conferma della presenza in Italia di Cobitis (Sabanejewia) larvata de Filippi (Osteichthyes, Cobitidae) // Natura. V. 60. F. 3. P. 176–184.
Coad B. 2017. Freshwater fishes of Iran. Cobitidae to Cyprinodontidae. (http://www.briancoad.com/Species%20Accounts/ Cobitidae%20to%20Cyprinodontidae.htm)
Economidis P.S., Nalbant T.T. 1996. A study of the loaches of the genera Cobitis and Sabanejewia (Pisces, Cobitidae) of Greece, with description of six new taxa // Trav. Mus. Nat. Hist. Natur. Grigore Antipa. V. 36. P. 295–347.
Eichwald C.E. 1838. Faunae Caspii maris primitiae // Bull. Soc. Imp. Naturalistes de Moscou. V. 11. P. 125–174.
Eschmeyer W.N., Fricke R., van der Laan R. (eds.). 2018. Catalog of fishes: genera, species, references (http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp. Version 03/2018)
Froese R., Pauly D. (eds.). 2017. FishBase. World Wide Web electronic publication. (www.fishbase.org. Version 10/2017).
Kottelat M., Freyhof J. 2007. Handbook of European freshwater fishes. Cornol, Switzerland; Berlin, Germany: Kottelat and Freyhof, 646 p.
Levan A., Fredga A., Sandberg A. 1964. Nomenclature for centromeric position on chromosomes // Hereditas. V. 52. № 2. P. 201–220.
Lodi E. 1979. Osservazioni intorno a Sabanejewia larvata (De Fil.) e alle sue relazioni con gli altri Cobitidi italiani (Pisces, Cypriniformes) // Bull. Mus. Zool. Univ. Torino. № 6. P. 135–144.
Lodi E., Marchionni V. 1980. Chromosome complement of the masked loach Sabanejewia larvata (De Fil.) (Pisces, Osteichthyes) // Caryologia. V. 33. № 4. P. 435–440.
Ludwig A., Bohlen J., Wolter C., Pitra C. 2001. Phylogenetic relationships and historical biogeography of spined loaches (Cobitidae, Cobitis and Sabanejewia) as indicated by variability of mitochondrial DNA // Zool. J. Linnean Soc. V. 131. P. 381–392.
Nalbant T. 1957. Cobitis aurata vallachica, eine neue Unterart des Balkan-Steinpeitzgers (Pisces, Cobitidae) // Senckenbergiana Biologica. V. 38. № 3/4. P. 209–212.
Nalbant T.T., Bianco P.G. 1998. The loaches of Iran and adjacent regions with description of six new species (Cobitoidea) // Ital. J. Zool. V. 65. Suppl. P. 109–123.
Perdices A., Doadrio I., Economidis P.S. et al. 2003. Pleistocene effects on the European freshwater fish fauna: double origin of the cobitid genus Sabanejewia in the Danube basin (Osteichthyes: Cobitidae) // Mol. Phylogen. Evolution. V. 26. P. 289–299.
Perdices A., Bohlen J., Šlechtová V., Doadrio I. 2016. Molecular evidence for multiple origins of the European spined loaches (Teleostei, Cobitidae) // PLoS ONE. V. 11. № 1. P. 1–18. (http://dx.doi.org/10.1371/journal.pone.0144628)
Vasil’ev V.P., Vasil’eva E.D. 1994. The karyological diversity in spined loaches from genera Cobitis and Sabanejewia // VIII Congr. Europ. Ichthyol. “Fishes and their environment”. Oviedo, Spain. P. 67.
Vladykov V.D. 1929. Sur un nouveau genre de Cobitides: Sabanejewia // Bull. Mus. Nat. Hist. Natur. Sér. 2. V. 1. № 1. P. 85–90.
Witkowski A. 1994. Morphological characteristics of Sabanejewia aurata (De Filippi, 1865) from the Odra River basin, with description of a new subspecies (Teleostei: Cypriniformes: Cobitidae) // Zool. Abhandl. Staat. Mus. Tierkunde Dresden. Bd. 48. № 3. P. 23–51.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии