

Вопросы ихтиологии, 2019, T. 59, № 3, стр. 258-267
Морфологическая изменчивость крупночешуйной краснопёрки Tribolodon hakonensis (Cyprinidae)
Н. С. Романов *
Национальный научный центр морской биологии Дальневосточного отделения РАН – ННЦМБ
Владивосток, Россия
* E-mail: n_romanov@inbox.ru
Поступила в редакцию 30.01.2018
После доработки 17.04.2018
Принята к публикации 02.06.2018
Аннотация
Исследована морфологическая изменчивость семи признаков двух форм крупночешуйной краснопёрки Tribolodon hakonensis из 10 водоёмов Дальнего Востока. Проанализированы три показателя – флуктуирующая асимметрия, географическая изменчивость и изменчивость значений признаков. Различия уровня флуктуирующей асимметрии T. hakonensis из разных выборок обусловлены разными условиями эмбрионального и раннего постэмбрионального развития. Северная форма отличается от южной бóльшим средним значением сумм дисперсий по всем признакам, что указывает на пониженную стабильность развития. Географическая изменчивость у T. hakonensis проявляется по шести признакам. Формы достоверно различаются между собой по средним значениям признаков, кроме числа подглазничных костей. Средние значения коэффициента вариации признаков варьируют в значительных пределах; формы между собой не различаются, и только по числу заглазничных костей у южной формы этот показатель заметно выше. Наибольшее число достоверных различий по дисперсии флуктуирующей асимметрии, средним значениям признаков и коэффициентам вариации наблюдается чаще всего при сравнении между выборками северной формы, а у южной – всегда минимальны или отсутствуют вовсе, что может быть как результатом отбора проб северной формы со значительно большей части ареала, так и более высоким уровнем нуклеотидной изменчивости у северной формы по сравнению с южной.
Крупночешуйная краснопёрка Tribolodon hakonensis была описана Гюнтером в 1877 г. из оз. Хаконе (о. Хонсю, Япония) как Leuciscus hakuensis (Богуцкая, Насека, 2004). Она, как и другие представители рода, ведёт проходной образ жизни: нагуливается в море, а на зимовку и нерест заходит в реки. Её ареал охватывает тихоокеанское побережье Азии от Тайваня на юге до Шантарских о-вов на севере; есть она на южных Курильских о-вах, Сахалине и островах Японского архипелага (Берг, 1949; Линдберг, Легеза, 1965; Шедько, 2002; Атлас …, 2003). Современные генетические исследования показали, что этот вид неоднороден и состоит из двух форм – северной и южной. Ареал южной формы простирается от Северного Приморья на юг, на остальной части ареала этого вида обитает северная форма (Семина и др., 2006; Семина, 2008; Брыков и др., 2011; Рязанова, Полякова, 2012). Проведённый морфологический анализ выявил значительные различия между этими формами (Гудков и др., 2010). Работ по морфологической изменчивости крупночешуйной краснопёрки довольно много (Kurawaka, 1977; Чуриков, Сабитов, 1982; Иванков и др., 1984; Богуцкая, 1988, 1990; Гавренков, 1988; Свиридов, 1999; Свиридов, Дюбанов, 2001; Свиридов, Иванков, 2002; Гриценко, 2002; Гудков и др., 2010), но вопросы флуктуирующей асимметрии в них не рассматриваются. Флуктуирующая асимметрия характеризует проявление случайной изменчивости развития индивидуума и является показателем степени стабильности развития. В некоторых работах (Tebb, Thoday, 1954; Beardmore, 1960; Valentine, Soule, 1973; Захаров, 1987; Romanov, 1995) показано, что благоприятным условиям среды соответствует определённый уровень флуктуирующей асимметрии (как отражение относительной стабильности развития), а при необычных условиях развития он может быть совершенно иным. Это позволяет использовать флуктуирующую асимметрию для контроля состояния природных популяций разных видов (Захаров, 1987; Биотест…, 1993; Захаров и др., 2000), что очень важно, так как от состояния конкретных природных популяций зависит как сохранение отдельных видов, так и нормальное функционирование экосистем в целом.
Цель работы – проанализировать флуктуирующую асимметрию, географическую изменчивость и изменчивость признаков крупночешуйной краснопёрки из разных районов её ареала.
МАТЕРИАЛ И МЕТОДИКА
Материалом послужили сборы крупночешуйной краснопёрки из разных частей ареала в пределах российского Дальнего Востока – Сахалинской области, Хабаровского и Приморского краёв (рис. 1; табл. 1).
Рис. 1.
Карта-схема района исследований и места сбора проб крупночешуйной краснопёрки Tribolodon hakonensis: 1 − оз. Большое, 2 – р. Тумнин, 3 − р. Тымь, 4 − р. Набиль, 5 – р. Фирсовка, 6 – р. Корсаковка, 7 – р. Киевка, 8 – р. Литовка, 9 – р. Артемовка, 10 – р. Раздольная.
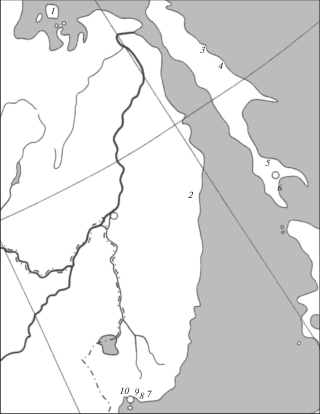
Таблица 1.
Объём собранного материала, доля асимметричных и асимметричных по числу признаков особей двух форм крупночешуйной краснопёрки Tribolodon hakonensis из водоёмов Дальнего Востока
| Водоём | Число рыб, экз. | Асимметричныеособи, % | Асимметричные по числу признаков, % | ||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |||
| Северная форма | |||||||
| Оз. Большое | 35 | 88.6 | 38.7 | 35.5 | 22.6 | 3.2 | – |
| Р. Тумнин | 75 | 92.0 | 43.5 | 31.9 | 21.7 | 2.9 | – |
| Р. Тымь | 53 | 96.2 | 43.1 | 29.4 | 23.5 | 7.8 | 7.8 |
| Р. Набиль | 58 | 94.8 | 27.8 | 47.3 | 21.8 | 3.6 | – |
| Р. Фирсовка | 35 | 97.1 | 32.4 | 35.3 | 23.5 | 5.9 | 2.9 |
| Р. Корсаковка | 64 | 89.1 | 19.3 | 38.6 | 28.1 | 8.8 | 5.3 |
| В среднем | 93.0 | 34.1 | 36.3 | 23.5 | 5.4 | 2.7 | |
| Южная форма | |||||||
| Р. Киевка | 62 | 91.9 | 35.1 | 26.3 | 24.6 | 8.8 | 5.3 |
| Р. Литовка | 96 | 87.5 | 34.5 | 38.1 | 19.0 | 6.0 | 2.4 |
| Р. Артёмовка | 49 | 79.6 | 48.7 | 25.6 | 17.9 | 7.7 | – |
| Р. Раздольная | 100 | 93.0 | 28.0 | 43.0 | 17.2 | 9.7 | 2.2 |
| В среднем | 88.0 | 36.6 | 33.3 | 19.7 | 8.1 | 2.5 | |
| В среднем обе формы | 91.0 | 35.1 | 35.1 | 22.1 | 6.4 | 2.6 | |
Для анализа были выбраны следующие билатеральные признаки: число ветвистых лучей в грудных (P) и брюшных (V) плавниках, число заглазничных (porb) и подглазничных (iorb) костей, число каналов второго порядка на первой заглазничной (porb-I), слёзной (lacr) и предкрышечной (pop) костях.
Флуктуирующую асимметрию оценивали по следующим показателям: 1) доля асимметричных рыб в выборке; 2) доля асимметричных особей по разному числу признаков от числа асимметричных рыб в выборке (число рыб асимметричных по одному признаку делится на число асимметричных рыб; число асимметричных по двум признакам рыб делится на число асимметричных рыб и т.д.); 3) доля асимметричных особей по каждому из признаков от общего числа случаев асимметрии в выборке (число случаев асимметрии в выборке представляет собой сумму случаев асимметрии по всем признакам); 4) дисперсия флуктуирующей асимметрии, которую рассчитывали по формуле, предложенной Палмером и Штробеком (Palmer, Strobeck, 1986):
Изменчивость признаков оценивали по коэффициенту вариации, который рассчитывался по формуле: СV = 100σ/M; его ошибку вычисляли по формуле:
Достоверность отличий средних значений (M) и коэффициентов вариации признаков определяли по величине t-критерия Стьюдента (Плохинский, 1970). Дендрограммы сходства строили методом полного сцепления. В качестве характеристики различий использовали Эвклидово расстояние.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Флуктуирующая асимметрия. Доля асимметричных особей в исследованных выборках крупночешуйной краснопёрки варьирует в довольно широких пределах (табл. 1): наименьшая характерна для рыб из р. Артёмовка (79.6%), а наибольшие – из рек Фирсовка (97.1%) и Тымь (96.2%). Среднее значение этого показателя заметно больше у северной формы, чем у южной (93 против 88%). Бóльшая часть рыб в выборках асимметрична по одному и двум признакам – в среднем по 35.1%, заметно меньше по трём (22.1%) и совсем немного по четырём (6.4%) признакам. Только в шести выборках были особи, асимметричные по пяти признакам.
В среднем у обеих форм крупночешуйной краснопёрки наибольшая частота встречаемости асимметрии (относительно общего числа случаев) характерна для числа каналов сейсмосенсорной системы на предкрышечных и слёзных костях и числа ветвистых лучей в грудных плавниках, а наименьшая – для числа заглазничных и подглазничных костей (табл. 2). У южной формы по сравнению с северной этот показатель намного больше по числу заглазничных костей и заметно меньше по числу каналов на предкрышечных костях.
Таблица 2.
Флуктуирующая асимметрия у двух форм крупночешуйной краснопёрки Tribolodon hakonensis из водоёмов Дальнего Востока
| Водоём | Доля асимметричных рыб, % числа случаев асимметрии | ||||||
|---|---|---|---|---|---|---|---|
| P | V | porb | iorb | porb-I | lacr | pop | |
| Северная форма | |||||||
| Оз. Большое | 25.4 | 3.4 | 5.1 | 5.1 | 11.9 | 23.7 | 25.4 |
| Р. Тумнин | 19.7 | 7.9 | 2.4 | 0.8 | 17.3 | 26.0 | 26.0 |
| Р. Тымь | 22.7 | 12.4 | 2.1 | 5.2 | 16.5 | 20.6 | 20.6 |
| Р. Набиль | 21.6 | 10.8 | 4.5 | 3.6 | 15.3 | 21.6 | 22.5 |
| Р. Фирсовка | 22.2 | 11.1 | 4.2 | 1.4 | 13.9 | 18.1 | 29.2 |
| Р. Корсаковка | 19.6 | 4.3 | 6.5 | 2.9 | 18.1 | 23.9 | 24.6 |
| В среднем | 21.9 | 8.3 | 4.1 | 3.2 | 15.5 | 22.3 | 24.7 |
| Южная форма | |||||||
| Р. Киевка | 22.0 | 8.7 | 6.3 | 3.1 | 20.5 | 20.5 | 18.9 |
| Р. Литовка | 17.5 | 9.4 | 5.3 | 4.7 | 16.4 | 24.6 | 22.2 |
| Р. Артёмовка | 26.4 | 6.9 | 5.6 | 2.8 | 18.1 | 22.2 | 18.1 |
| Р. Раздольная | 21.4 | 7.5 | 7.0 | 3.0 | 16.4 | 21.4 | 23.4 |
| В среднем | 21.8 | 8.1 | 6.1 | 3.4 | 17.9 | 22.2 | 20.7 |
| В среднем обе формы | 21.9 | 8.2 | 4.9 | 3.3 | 16.4 | 22.3 | 23.1 |
Примечание. Здесь и в табл. 3−7: P и V − число ветвистых лучей в грудных и брюшных плавниках, porb, iorb − число заглазничных и подглазничных костей, porb-I, lacr, pop – число каналов сейсмосенсорной системы соответственно на первой заглазничной, слёзной и предкрышечной костях.
Минимальные средние значения дисперсии флуктуирующей асимметрии (табл. 3) характерны для числа лучей в грудных (0.862) и брюшных (1.844) плавниках, а максимальные – для числа каналов на первых заглазничных костях (41.194). Выборки крупночешуйной краснопёрки достоверно различаются по дисперсии флуктуирующей асимметрии в следующем числе случаев: 24 – по iorb, 22 – porb, 21 – V, 14 – porb-I, 3 – pop; по P и lacr различия недостоверны. Формы крупночешуйной краснопёрки несущественно отличаются по средним значениям дисперсии флуктуирующей асимметрии по большинству признаков и только по porb-I у северной формы дисперсия в 1.5 раза больше, чем у южной. У северной формы сумма дисперсий по всем признакам варьирует в значительных пределах – от 59.707 (р. Фирсовка) до 141.713 (р. Тымь), а у южной колебания этого показателя невелики, что можно объяснить как бóльшим разнообразием условий развития из-за взятия проб северной формы со значительно бóльшего ареала, так и более высоким уровнем нуклеотидной изменчивости у северной формы по сравнению с южной – 0.013146 против 0.001130 (Рязанова, Полякова, 2012). Также можно отметить, что среднее значение суммы дисперсий по всем признакам у северной формы намного больше, чем у южной. Высокий уровень флуктуирующей асимметрии, а значит, пониженная стабильность развития у краснопёрки из оз. Большое и р. Тымь, вероятно, являются результатом нахождения этих популяций на северной границе ареала, а из р. Корсаковка – значительным антропогенным воздействием на экосистему этой реки.
Таблица 3.
Дисперсия флуктуирующей асимметрии (×10–3) некоторых признаков у двух форм крупночешуйной краснопёрки Tribolodon hakonensis из водоёмов Дальнего Востока
| Водоём | Признак | |||||||
|---|---|---|---|---|---|---|---|---|
| P | V | porb | iorb | porb-I | lacr | pop | Σ | |
| Северная форма | ||||||||
| Оз. Большое | 0.918 | 0.812 | 10.084 | 11.709 | 50.438 | 14.660 | 4.867 | 93.488 |
| Р. Тумнин | 0.814 | 1.651 | 3.891 | 1.935 | 33.274 | 14.812 | 5.416 | 61.793 |
| Р. Тымь | 0.816 | 2.783 | 5.552 | 12.639 | 99.681 | 15.425 | 4.817 | 141.713 |
| Р. Набиль | 0.988 | 2.444 | 9.629 | 9.483 | 38.131 | 15.841 | 5.680 | 82.196 |
| Р. Фирсовка | 0.871 | 2.550 | 8.065 | 4.147 | 26.645 | 12.564 | 4.865 | 59.707 |
| Р. Корсаковка | 0.879 | 1.211 | 18.417 | 8.645 | 40.149 | 16.086 | 5.901 | 91.288 |
| В среднем | 0.881 | 1.909 | 9.273 | 8.093 | 48.053 | 14.898 | 5.258 | 88.315 |
| Южная форма | ||||||||
| Р. Киевка | 0.870 | 2.247 | 8.515 | 8.901 | 36.631 | 12.240 | 4.383 | 73.787 |
| Р. Литовка | 0.744 | 1.858 | 11.460 | 11.204 | 33.648 | 13.626 | 4.857 | 77.397 |
| Р. Артёмовка | 0.838 | 1.198 | 10.057 | 5.801 | 26.279 | 12.526 | 3.623 | 60.322 |
| Р. Раздольная | 0.880 | 1,689 | 11.897 | 8.268 | 27.064 | 13.014 | 4.365 | 67.177 |
| В среднем | 0.833 | 1.748 | 10.482 | 8.544 | 30.906 | 12.852 | 4.307 | 69.670 |
| В среднем обе формы | 0.862 | 1.844 | 9.757 | 8.273 | 41.194 | 14.079 | 4.877 | 80.887 |
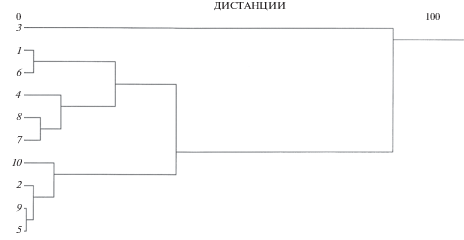
Кластеризация выборок крупночешуйной краснопёрки по сумме дисперсий по всем признакам показывает разбиение на четыре кластера (рис. 2): выборки с наименьшими значениями (Тумнин, Фирсовка, Артемовка, Раздольная) входят в кластер, к которому примыкают выборки со средними (Набиль, Киевка, Литовка) и высокими (Большое, Корсаковка) значениями, обособленно располагается кластер р. Тымь. Таким образом, половину кластеров образуют выборки обеих форм, а другую – северной формы.
Рис. 2.
Дендрограмма сходства выборок крупночешуйной краснопёрки Tribolodon hakonensis по сумме дисперсий флуктуирующей асимметрии исследованных признаков; обозначения см. на рис. 1.
Географическая изменчивость. Значения анализируемых признаков северной и южной формы красночешуйной краснопёрки из разных водоёмов приведены в табл. 4, критерия Стьюдента при их сравнении – в табл. 5. Число ветвистых лучей в грудных плавниках у северной формы варьирует в пределах 13–18, у южной – 15–18. Для северной формы можно отметить увеличение, а для южной – уменьшение средних значений этого признака с севера на юг. Достоверные различия средних значений между выборками отмечаются в следующем числе случаев: у северной – в 12, при сравнении северной с южной – в 14, у южной – в одном. Сравнение средних значений этого признака между формами крупночешуйной краснопёрки показывает значительные достоверные различия.
Таблица 4.
Значения признаков у двух форм крупночешуйной краснопёрки Tribolodon hakonensis из водоёмов Дальнего Востока
| Признак | Северная форма | Южная форма | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Оз. Большое | Р. Тумнин | Р. Тымь | Р. Набиль | Р. Фирсовка | Р. Корсаковка | Р. Киевка | Р. Литовка | Р. Артёмовка | Р. Раздольная | |
| P | $\frac{{15.73 \pm 0.13}}{{15{\kern 1pt} --{\kern 1pt} 18}}$ | $\frac{{15.93 \pm 0.08}}{{15{\kern 1pt} --{\kern 1pt} 18}}$ | $\frac{{16.68 \pm 0.11}}{{15{\kern 1pt} --{\kern 1pt} 19}}$ | $\frac{{15.36 \pm 0.13}}{{13{\kern 1pt} --{\kern 1pt} 18}}$ | $\frac{{16.57 \pm 0.13}}{{15{\kern 1pt} --{\kern 1pt} 18}}$ | $\frac{{16.33 \pm 0.10}}{{15{\kern 1pt} --{\kern 1pt} 18}}$ | $\frac{{16.55 \pm 0.073}}{{15{\kern 1pt} --{\kern 1pt} 18}}$ | $\frac{{16.43 \pm 0.08}}{{15{\kern 1pt} --{\kern 1pt} 18}}$ | $\frac{{16.44 \pm 0.06}}{{15{\kern 1pt} --{\kern 1pt} 18}}$ | $\frac{{16.35 \pm 0.07}}{{15{\kern 1pt} --{\kern 1pt} 18}}$ |
| V | $\frac{{8.03 \pm 0.04}}{{7{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{8.13 \pm 0.05}}{{7{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{7.91 \pm 0.06}}{{7{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{7.93 \pm 0.07}}{{6{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{8.23 \pm 0.10}}{{7{\kern 1pt} --{\kern 1pt} 10}}$ | $\frac{{8.13 \pm 0.05}}{{7{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{8.21 \pm 0.044}}{{6{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{8.17 \pm 0.04}}{{7{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{8.50 \pm 0.05}}{{8{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{8.32 \pm 0.05}}{{7{\kern 1pt} --{\kern 1pt} 9}}$ |
| porb | $\frac{{2.96 \pm 0.045}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{3.07 \pm 0.03}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{2.98 \pm 0.02}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.99 \pm 0.03}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{3.01 \pm 0.035}}{{3{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{2.93 \pm 0.03}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{3.06 \pm 0.022}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{2.97 \pm 0.02}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{3.00 \pm 0.025}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{3.11 \pm 0.04}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ |
| iorb | $\frac{{2.04 \pm 0.03}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.01 \pm 0.01}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.05 \pm 0.03}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.03 \pm 0.024}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.01 \pm 0.02}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.05 \pm 0.03}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.03 \pm 0.016}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.04 \pm 0.02}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{\,2.02 \pm 0.014}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.03 \pm 0.02}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ |
| porb-I | $\frac{{1.99 \pm 0.05}}{{1{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.39 \pm 0.08}}{{1{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{2.13 \pm 0.06}}{{1{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.16 \pm 0.05}}{{1{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.57 \pm 0.16}}{{2{\kern 1pt} --{\kern 1pt} 6}}$ | $\frac{{2.27 \pm 0.06}}{{1{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{2.60 \pm 0.085}}{{1{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{2.34 \pm 0.06}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{2.02 \pm 0.014}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{2.89 \pm 0.10}}{{2{\kern 1pt} --{\kern 1pt} 6}}$ |
| lacr | $\frac{{4.11 \pm 0.09}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{4.13 \pm 0.08}}{{2{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{3.98 \pm 0.08}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{4.07 \pm 0.08}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{4.51 \pm 0.10}}{{3{\kern 1pt} --{\kern 1pt} 6}}$ | $\frac{{4.17 \pm 0.08}}{{2{\kern 1pt} --{\kern 1pt} 6}}$ | $\frac{{4.33 \pm 0.060}}{{3{\kern 1pt} --{\kern 1pt} 6}}$ | $\frac{{4.30 \pm 0.06}}{{3{\kern 1pt} --{\kern 1pt} 6}}$ | $\frac{{4.39 \pm 0.059}}{{3{\kern 1pt} --{\kern 1pt} 5}}$ | $\frac{{4.43 \pm 0.07}}{{2{\kern 1pt} --{\kern 1pt} 6}}$ |
| pop | $\frac{{6.64 \pm 0.12}}{{6{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{6.87 \pm 0.11}}{{5{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{6.92 \pm 0.10}}{{5{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{6.42 \pm 0.08}}{{5{\kern 1pt} --{\kern 1pt} 8}}$ | $\frac{{7.07 \pm 0.25}}{{5{\kern 1pt} --{\kern 1pt} 11}}$ | $\frac{{6.38 \pm 0.10}}{{5{\kern 1pt} --{\kern 1pt} 10}}$ | $\frac{{7.44 \pm 0.108}}{{6{\kern 1pt} --{\kern 1pt} 11}}$ | $\frac{{6.92 \pm 0.08}}{{5{\kern 1pt} --{\kern 1pt} 10}}$ | $\frac{{7.43 \pm 0.09}}{{6{\kern 1pt} --{\kern 1pt} 9}}$ | $\frac{{6.92 \pm 0.08}}{{6{\kern 1pt} --{\kern 1pt} 11}}$ |
Примечание. Здесь и в табл. 5: над чертой − среднее значение и его ошибка, под чертой – пределы варьирования показателя.
Таблица 5.
Значения признаков у двух форм крупночешуйной краснопёрки Tribolodon hakonensis и критерия Стьюдента при их сравнении
| Признак | Северная форма | Южная форма | tst |
|---|---|---|---|
| P | $\frac{{16.08 \pm 0.04}}{{13{\kern 1pt} --{\kern 1pt} 19}}$ | $\frac{{16.43 \pm 0.03}}{{15{\kern 1pt} --{\kern 1pt} 18}}$ | 7.0 |
| V | $\frac{{8.06 \pm 0.02}}{{6{\kern 1pt} --{\kern 1pt} 10}}$ | $\frac{{8.28 \pm 0.02}}{{6{\kern 1pt} --{\kern 1pt} 9}}$ | 7.8 |
| porb | $\frac{{3.00 \pm 0.01}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | $\frac{{3.033 \pm 0.012}}{{2{\kern 1pt} --{\kern 1pt} 4}}$ | 2.1 |
| iorb | $\frac{{2.03 \pm 0.007}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | $\frac{{2.03 \pm 0.007}}{{2{\kern 1pt} --{\kern 1pt} 3}}$ | 0.0 |
| porb-I | $\frac{{2.26 \pm 0.02}}{{1{\kern 1pt} --{\kern 1pt} 6}}$ | $\frac{{2.63 \pm 0.03}}{{1{\kern 1pt} --{\kern 1pt} 6}}$ | 10.3 |
| lacr | $\frac{{4.14 \pm 0.03}}{{2{\kern 1pt} --{\kern 1pt} 6}}$ | $\frac{{4.36 \pm 0.03}}{{2{\kern 1pt} --{\kern 1pt} 6}}$ | 5.2 |
| pop | $\frac{{6.70 \pm 0.04}}{{5{\kern 1pt} --{\kern 1pt} 11}}$ | $\frac{{7.38 \pm 0.04}}{{5{\kern 1pt} --{\kern 1pt} 11}}$ | 12.0 |
Число ветвистых лучей в брюшных плавниках северной формы 6–10, южной – 6–9. У обеих форм наблюдается увеличение средних значений этого признака с севера на юг. Между выборками отмечаются достоверные отличия: у северной – в 7, при сравнении северной с южной – в 17, у южной – в 4 случаях. Южная форма достоверно отличается от северной.
Число заглазничных костей у обеих форм варьирует в пределах двух–четырёх. Для северной формы характерно некоторое уменьшение, а для южной – увеличение значений этого признака с севера на юг. Достоверные различия средних значений между выборками отмечаются в следующем числе случаев: у северной – в трёх, при сравнении северной с южной – в восьми, у южной – в трёх. Различия между формами невелики.
Число подглазничных костей у крупночешуйной краснопёрки колеблется в небольших пределах – две–три. Какой-то стройной картины географической изменчивости этого признака не отмечается, так же как и достоверных различий между выборками и формами.
Варьирование числа каналов на первой заглазничной кости у крупночешуйной краснопёрки отмечается в пределах 1–6. Для обеих форм характерно увеличение средних значений этого признака с севера на юг. Достоверные различия средних значений между выборками отмечаются в следующем числе случаев: у северной – в 10, при сравнении северной с южной – в 17, у южной – в 4. Различия между формами высоко достоверны.
Число каналов на слёзной кости варьирует в широких пределах – 2–6. Для обеих форм можно отметить увеличение средних значений этого признака с севера на юг. Между выборками отмечаются достоверные отличия: у северной – в 5, при сравнении северной с южной – в 16 случаях, а выборки южной формы не различаются. Различия между формами существенны.
Число каналов на предкрышечной кости варьирует в пределах 5–11. Для обеих форм характерно увеличение средних значений этого признака с севера на юг. Достоверные различия средних значений между выборками отмечаются в следующем числе случаев: у северной – в 6, при сравнении северной с южной – в 19, у южной – в 5. Различия между формами по этому признаку высоко достоверны.
Таким образом, географическая изменчивость у крупночешуйной краснопёрки проявляется по шести из семи исследованных признаков: по четырём (V, porb-I, lacr, pop) – у обеих форм увеличение средних значений с севера на юг, а по двум (P, porb) – разнонаправленные изменения у северной и южной формы. Достоверные отличия между выборками по этому показателю отмечаются в 151 случае: 31 – porb-I, 30 – pop, 28 – V, 27 – P, 21 – lacr, 14 – porb. В 60.3% случаев это различия между формами, в 28.5% – между выборками северной и в 11.3% – южной формы. Формы крупночешуйной краснопёрки достоверно различаются между собой по средним значениям признаков, кроме числа подглазничных костей (табл. 5).
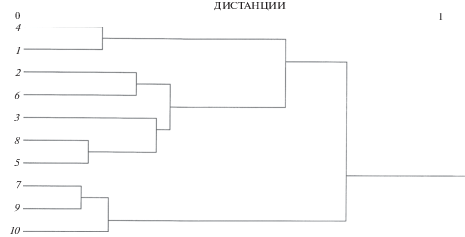
Кластеризация выборок крупночешуйной краснопёрки по средним значениям признаков демонстрирует разбиение на четыре кластера (рис. 3). Один объединяет выборки из рек Тымь, Фирсовка и Литовка, к нему примыкает другой из выборок рек Тумнин и Корсаковка; к этой паре присоединяется кластер оз. Большое и р. Набиль. Обособленно располагается кластер из рек Киевка, Артёмовка, Раздольная. Таким образом, в двух кластерах объединяются выборки северной формы, в одном – южной, а один кластер оказывается смешанным.
Рис. 3.
Дендрограмма сходства выборок крупночешуйной краснопёрки Tribolodon hakonensis по средним значениям исследованных признаков; обозначения см. на рис. 1.
Изменчивость значений признаков. Средние значения коэффициента вариации признаков варьируют в широких пределах – от 4.78 (P) до 24.51 (porb-I); по анализируемым признакам формы не различаются, и только значение porb у южной формы заметно выше (табл. 6). У северной формы разброс значений коэффициента вариации между выборками больше, чем у южной, по большинству признаков, кроме porb-I. Колебания суммы значений коэффициента вариации по всем признакам между выборками южной формы больше, чем у северной, а по средним значениям этого показателя формы между собой не различаются.
Таблица 6.
Коэффициент вариации значений признаков у двух форм крупночешуйной краснопёрки Tribolodon hakonensis из водоёмов Дальнего Востока
| Водоём | Признак | |||||||
|---|---|---|---|---|---|---|---|---|
| P | V | porb | iorb | porb-I | lacr | pop | Σ | |
| Северная форма | ||||||||
| Оз. Большое | 4.83 | 2.98 | 8.99 | 10.00 | 15.98 | 12.80 | 10.59 | 66.17 |
| Р. Тумнин | 4.61 | 5.26 | 9.32 | 4.08 | 27.74 | 16.90 | 14.28 | 82.19 |
| Р. Тымь | 4.65 | 5.39 | 4.60 | 10.39 | 21.55 | 15.10 | 10.87 | 72.55 |
| Р. Набиль | 6.34 | 7.00 | 6.96 | 9.02 | 20.19 | 15.11 | 9.89 | 74.51 |
| Р. Фирсовка | 4.53 | 7.17 | 6.98 | 5.97 | 37.35 | 13.53 | 20.79 | 96.32 |
| Р. Корсаковка | 4.93 | 4.62 | 8.77 | 10.34 | 22.60 | 16.24 | 12.90 | 80.4 |
| В среднем | 4.98 | 5.40 | 7.60 | 8.30 | 24.24 | 14.95 | 13.22 | 78.69 |
| Южная форма | ||||||||
| Р. Киевка | 4.89 | 6.07 | 8.07 | 8.72 | 34.62 | 15.50 | 16.26 | 94.13 |
| Р. Литовка | 4.51 | 5.21 | 7.24 | 9.80 | 24.50 | 14.72 | 11.25 | 77.23 |
| Р. Артёмовка | 3.91 | 5.92 | 8.30 | 7.03 | 7.03 | 13.35 | 11.43 | 56.97 |
| Р. Раздольная | 4.58 | 5.97 | 11.82 | 8.42 | 33.49 | 15.10 | 11.26 | 90.64 |
| В среднем | 4.47 | 5.79 | 8.86 | 8.49 | 24.91 | 14.67 | 12.55 | 79.74 |
| В среднем обе формы | 4.78 | 5.56 | 8.11 | 8.38 | 24.51 | 14.84 | 12.95 | 79.11 |
Достоверные различия коэффициента вариации между выборками крупночешуйной краснопёрки по porb-I наблюдаются в 28 случаях, по iorb – в 21, по porb и pop, – в 20, по V, – 17, по P – в 8, по lacr – в одном случае (табл. 7). Наибольшее число достоверных различий между выборками по коэффициенту вариации наблюдается чаще всего при сравнении форм, реже – у северной; различия между выборками южной формы всегда минимальны или отсутствуют вовсе.
Таблица 7.
Число достоверных различий по коэффициенту вариации между выборками двух форм крупночешуйной краснопёрки Tribolodon hakonensis
| Форма | Признак | |||||||
|---|---|---|---|---|---|---|---|---|
| P | V | porb | iorb | porb-I | pop | lacr | Σ | |
| Северная | 4 | 9 | 7 | 9 | 8 | 9 | 1 | 47 |
| Северная c южной | 4 | 8 | 10 | 10 | 15 | 8 | 0 | 55 |
| Южная | 0 | 0 | 3 | 2 | 5 | 3 | 0 | 13 |
| Обе | 8 | 17 | 20 | 21 | 28 | 20 | 1 | |
Кластеризация выборок крупночешуйной краснопёрки по сумме коэффициентов вариации по всем признакам показывает разбиение на три кластера (рис. 4). В самый большой входят выборки северной формы и одна южной со средними значениями этого показателя; к этому кластеру примыкает другой – с самыми высокими значениями. Обособленно располагается третий кластер с минимальными значениями. Примечательно, что ни один кластер не состоит из выборок одной формы.
Рис. 4.
Дендрограмма сходства выборок крупночешуйной краснопёрки Tribolodon hakonensis по сумме коэффициентов вариации исследованных признаков; обозначения см. на рис. 1.

ЗАКЛЮЧЕНИЕ
Наименьшая дисперсия флуктуирующей асимметрии характерна для числа ветвистых лучей в брюшных и грудных плавниках, а наибольшая – для числа каналов на первых заглазничных костях. Среднее значение последнего признака у северной формы существенно больше, чем у южной. У северной формы сумма дисперсий по всем признакам варьирует в широких пределах, а у южной колебания этого показателя невелики, что можно объяснить как бóльшим разнообразием условий развития из-за взятия проб северной формы со значительно большего ареала, так и более высоким уровнем нуклеотидной изменчивости у северной формы по сравнению с южной (Рязанова, Полякова, 2012). Северная форма отличается от южной бóльшим средним значением сумм дисперсий по всем признакам, что говорит о меньшей стабильности условий развития рыб северной формы. Это может быть обусловлено как расположением некоторых выборок на северной границе ареала, так и значительным антропогенным воздействием.
Географическая изменчивость у крупночешуйной краснопёрки проявляется по шести из семи исследованных признаков: по четырём (V, porb-1, lacr, pop) наблюдается увеличение средних значений с севера на юг у обеих форм, а по двум (P, porb) – изменения разнонаправлены при сравнении форм. Достоверные различия между выборками по этому показателю отмечаются в 151 случае (31 – porb-I, 30 – pop, 28 – V, 27 – P, 21 – lacr, 14 – porb). В 60.3% случаев это различия между формами, в 28.5% – между выборками северной и в 11.3% – южной формы. Формы крупночешуйной краснопёрки достоверно различаются между собой по средним значениям признаков, кроме числа подглазничных костей.
Средние значения коэффициента вариации признаков варьируют в широких пределах; формы между собой не различаются, и только по числу заглазничных костей у южной формы заметно выше. У северной формы разброс значений коэффициента вариации между выборками больше, чем у южной, по большинству признаков, кроме числа каналов на первой заглазничной кости, что может быть обусловлено более высоким уровнем нуклеотидной изменчивости у северной формы (Рязанова, Полякова, 2012). Формы крупночешуйной краснопёрки не различаются средними значениями суммы коэффициентов вариации по всем признакам. Наибольшее число достоверных различий между выборками по коэффициентам вариации наблюдается чаще всего при сравнении форм, реже – между выборками северной, а между выборками южной формы всегда минимальны или отсутствуют вовсе.
Список литературы
Атлас пресноводных рыб России: в 2 томах. 2003. Т. 1 / Под ред. Решетникова Ю.С. М.: Наука, 379 с.
Берг Л.С. 1949. Рыбы пресных вод СССР и сопредельных стран. Ч. 2. М.; Л.: Изд-во АН СССР. С. 469–925.
Биотест: интегральная оценка здоровья экосистем и отдельных видов. 1993 / Под ред. В.М. Захарова, Д.М. Кларка М:. Мос. отд. МФ “Биотест”, 68 с.
Богуцкая Н.Г. 1988. Топография каналов сейсмосенсорной системы карповых рыб подсемейств Leuciscinae, Xenocyprinidae и Cultrinae // Вопр. ихтиологии. Т. 28. Вып. 3. С. 367–382.
Богуцкая Н.Г. 1990. Морфологические основы системы Карповых рыб подсемейства Ельцовых (Leuciscinae, Cyprinidae) // Там же. Т. 30. Вып. 6. С. 920–933.
Богуцкая Н.Г., Насека А.М. 2004. Каталог бесчелюстных и рыб пресных и солоноватых вод России с номенклатурными и таксономическими комментариями. М.: Т-во науч. изд. КМК, 389 с.
Брыков Вл.А., Полякова Н.Е., Семина А.В. 2011. Филогенетический анализ выявляет два периода дивергенции у крупночешуйной красноперки Tribolodon hakonensis (Pisces, Cyprinidae) // Генетика. Т. 47. № 11. С. 1491–1500.
Гавренков Ю.И. 1988. Биология, морфология и состояние запасов дальневосточных красноперок рода Tribolodon в Южном Приморье // Изв. ТИНРО. Т. 123. С. 74–81.
Гриценко О.Ф. 2002. Проходные рыбы острова Сахалин (систематика, экология, промысел). М.: Изд-во ВНИРО, 247 с.
Гудков П.К., Полякова Н.Е., Семина А.В., Назаркин М.В. 2010. Сравнительный морфологический анализ крупночешуйной красноперки Tribolodon hakonensis Günther, 1877 (Cyprinidae, Cpriniformes) Сахалина и южного Приморья // Вопр. ихтиологии. Т. 50. № 6. С. 772–776.
Захаров В.М. 1987. Асимметрия животных. M.: Наука, 216 с.
Захаров В.М., Баранов А.С., Борисов В.И. и др. 2000. Здоровье среды: методика оценки. М.: Центр экол. политики России, 68 с.
Иванков В.Н., Лукьянов П.У., Мостовая Н.В., Рухлова Г.Ф. 1984. Таксономическое значение морфологических признаков молоди двух видов дальневосточных красноперок // Биология моря. № 3. С. 29–33.
Линдберг Г.У., Легеза М.Н. 1965. Рыбы Японского моря и сопредельных частей Охотского и Желтого морей. Ч. 2. М.: Наука, 391 с.
Плохинский Н.А. 1970. Биометрия. М.: Изд-во МГУ, 367 с.
Рязанова И.Н., Полякова Н.Е. 2012. Дифференциация крупночешуйной красноперки Tribolodon hakonensis (Pisces: Cyprinidae) на российской части ареала по данным кариологического анализа и ПЦР-ПДРФ-анализа митохондриальной ДНК // Генетика. Т. 48. № 2. С. 225–234.
Свиридов В.В. 1999. Морфология и структура чешуи популяций дальневосточных красноперок рода Tribolodon Приморья и южного Сахалина // Матер. II регион. конф. «Актуальные проблемы морской биологии, экологии и биотехнологии». Владивосток: Изд-во ДВГУ. С. 135–136.
Свиридов В.В., Дюбанов А.М. 2001. Особенности подвесочного аппарата дальльневосточных красноперок (Tribolodon, Cyprinidae) // Матер. IV регион. конф. “Актуальные проблемы морской биологии, экологии и биотехнологии”. Владивосток: Изд-во ДВГУ. С. 108–110.
Свиридов В.В., Иванков В.Н. 2002. Топография сейсмосенсорных каналов головы молоди дальневосточных красноперок рода Tribolodon (Сyprinidae) и ее значение для видовой диагностики // Вопр. ихтиологии. Т. 42. № 3. С. 418–420.
Семина А.В. 2008. Молекулярная эволюция и филогенетические отношения в двух группах рыб семейств Mugilidae и Cyprinidae: Автореф. дис. … канд. биол. наук. Владивосток: ИБМ ДВО РАН, 23 с.
Семина А.В., Полякова Н.Е., Брыков В.А. 2006. Генетический анализ выявляет криптический вид у дальневосточных красноперок рода Tribolodon // ДАН. Т. 407. № 4. С. 571–573.
Чуриков А.А., Сабитов Э.Х. 1982. Дополнение к диагнозу дальневосточных красноперок // Вопр. ихтиологии. Т. 22. Вып. 5. С. 881–883.
Шедько С.В. 2002. Обзор пресноводной ихтиофауны // Растительный и животный мир Курильских островов. Владивосток: Дальнаука. С. 118–134.
Beardmore J.A. 1960. Developmental stability in constant and fluctuating temperatures // Heredity. V. 14. P. 411–422.
Graham J., Freeman D.C., Emlen J. 1993. Antisymmetry, directional asymmetry and dynamic morphogenesis // Genetica. V. 89. № 1−3. P. 121−137.
Kurawaka K. 1977. Cephalic lateral-line systems and geographical distribution of the genus Tribolodon (Cyprinidae) // Jpn. J. Ichthyol. V. 24. № 3. P. 167–175.
Palmer A.R., Strobeck C. 1986. Fluctuating asymmetry: measurement, analysis, patterns // Ann. Rev. Ecol. Syst. № 17. P. 391−421.
Romanov N.S. 1995. Fluctuating asymmetry in chum salmon, Oncorhynchus keta, from the Maritime Province // J. Ichthyol. V. 35. № 9. P. 171−182.
Sokal R.R., Rohlf F.J. 1981. Biometry. N.Y.: Freeman and Co., 859 p.
Tebb G., Thoday J.M. 1954. Stability in development and relational balance of X-chromosomes in Drosophila melanogaster // Nature. V. 174. P. 1109–1110.
Valentine D.V., Soule M. 1973. Effect of p,p'-DDT on developmental stability of pectoral fin rays in the grunion, Leuresthes tenuis // Nat. Mar. Fish. Serv. Fish. Bull. V. 71. P. 921–925.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии