

Вопросы ихтиологии, 2019, T. 59, № 3, стр. 251-256
Описание нового вида пелагической бельдюги Melanostigma japonicum sp. nova (Zoarcidae) от тихоокеанского побережья юга Японии с ключом для определения видов рода из Тихого океана и сопредельных акваторий Индийского и Южного океанов
А. В. Балушкин *
Зоологический институт РАН − ЗИН РАН
Санкт-Петербург, Россия
* E-mail: ichthlab@zin.ru
Поступила в редакцию 08.10.2018
После доработки 08.10.2018
Принята к публикации 11.10.2018
Аннотация
Описан новый вид Melanostigma japonicum sp. n. из зал. Тоса (Южная Япония). Голотип нового вида, идентифицированный ранее как M. orientale, отличается от этого и других видов меланостигм уникальным строением челюстей с очень крупными выступающими вперёд передними зубами, оригинальной комбинацией ряда счётных признаков, окраской и пропорциями тела. Восстановлен M. flaccidum, ранее сводимый в синонимию M. gelatinosum. Обсуждаются вопросы происхождения и исторического расселения меланостигм. Составлен ключ для определения видов рода в Тихом океане и на сопряжённых акваториях Индийского и Южного океанов.
К настоящему времени в Тихом океане и сопредельных водах Индийского и Южного океанов насчитывается восемь валидных видов космополитического рода пелагических бельдюг Melanostigma Gȕnther: M. bathium Bussing, 1965 (тихоокеанское побережье Южной Америки), M. flaccidum Waite, 1914 (восточное побережье Новой Зеландии), M. gelatinosum Günther, 1881 (s. str.: тихоокеанское побережье Южной Америки, Магелланов пролив), M. kharini Balushkin et Moganova, 2018 (хребет Геракл, Антарктическо-Южнотихоокеанское поднятие), M. inexpectatum Parin, 1977 (Западная экваториальная Пацифика), M. orientale Tominaga, 1971 (тихоокеанское побережье Японии), M. pammelas Gilbert, 1896 (Северо-Восточная Пацифика) и M. vitiazi Parin, 1979 (море Банда, Индо-Вест-Пацифика).
В водах Японии восточная меланостигма M. orientale была впервые описана по трём экземплярам, пойманным на мезобатиальных глубинах в заливах Сагами (голотип) и Суруга (паратипы) (Tominaga, 1971). Это были первые находки представителей рода на северо-западе Тихого океана. Позднее Машида и Ли (Machida, Lee, 1997) сообщили о трёх новых находках меланостигм у берегов Японии, которых они идентифицировали как M. orientale. К сожалению, для двух экземпляров отсутствовали данные о времени и месте поимок, третий же экземпляр был выловлен в зал. Тоса (Южная Япония). Этот последний экземпляр заметно отличался от типовых экземпляров вида. Прежде всего, это касалось общего числа позвонков (110 против 98−100 у типовых экземпляров), но авторы расценили это лишь как проявление внутривидовой изменчивости. Позднее широкие пределы варьирования числа позвонков (93−110) вошли в диагнозы M. orientale в некоторых обзорных работах по рыбам Японии (Hatooka, 2002, 2013). Вместе с тем необычность этой рыбы заключалась даже не в самом факте значительного увеличения у неё числа позвонков, а в том, что такое изменение не привело к соответствующему увеличению других метамерных структур осевого скелета − лучей в спинном (D) и анальном (A) плавниках. По данным Машида и Ли (Machida, Lee, 1997), у этого экземпляра было D 95 и A 80, что лишь на один луч превышает максимальные значения у типовых экземпляров M. orientale (D 94 и A 79).
Возможность изучить экземпляр меланостигмы из зал. Тоса я получил благодаря визиту в ЗИН РАН японского ихтиолога Рио Мисава (Ryo Misawa), который по моей просьбе привёз его с собой из музейной коллекции биологического департамента Университета города Коти. Изучение показало, что он резко отличен от всех известных видов Melanostigma и описан здесь как новый вид рода − Melanostigma japonicum sp. n. Вместе с тем при пересчёте позвонков оказалось, что данные Машида и Ли (Machida, Lee, 1997) были ошибочными, поскольку у рыбы было не 110, а только 100 позвонков (20 туловищных + 80 хвостовых).
МАТЕРИАЛ И МЕТОДИКА
В сравнительных целях были изучены представители других видов рода, хранящиеся в коллекциях ЗИН РАН (Balushkin et al., 2011; Балушкин и др., 2012). В их числе типовые экземпляры следующих видов, обитающих в исследуемых районах: M. inexpectatum Parin (ЗИН № 42640), M. vitiazi Parin (ЗИН № 44000) и M. kharini Balushkin et Moganova (ЗИН №№ 56163, 56164). В моём распоряжении была также рентгенограмма голотипа M. orientale. Считая вид M. gelatinosum, в понимании ряда авторов (McAllister, Ress, 1964; Anderson, 1988, 1990), сборным таксоном, в данной работе я отношу к нему только рыб, обитающих на юге Южной Америки (типовое обитание – Магелланов пролив). Ранее сводимая в синонимию к этому виду новозеландская меланостигма M. flaccidum рассматривается здесь в качестве валидного вида. Для этих двух видов в определительную таблицу были включены только данные типовых экземпляров (McAllister, Ress, 1964; Bussing, 1965) и собственные данные по экземпляру M. gelatinosum из вод Чили (ЗИН № 49064 − TL 73 мм, “Академик Книпович”, 42°24′ ю.ш. 74°45′ з.д., глубина 470−440 м, 26.02.1973 г., коллекторы В.Н. Ефременко, В.П. Природина).
В статье использованы следующие сокращения естественнонаучных музеев: NSMT − Национальный музей науки, Токио, Япония; BSKU − Музей биологического департамента Университета города Коти, Япония. В данной работе сохранена последовательность описания меланостигм, принятая в предшествующих работах (Балушкин, Моганова, 2017, 2018).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
(рис. 1, 2)
Рис. 1.
Голотип Melanostigma japonicum Balushkin, sp. n. − SL 98 мм, Южная Япония, зал. Тоса, 32°59′19″ с.ш. 133°35′52″ в.д., глубина 700−720 м. Фото Рио Мисава (Ryo Misawa).

Рис. 2.
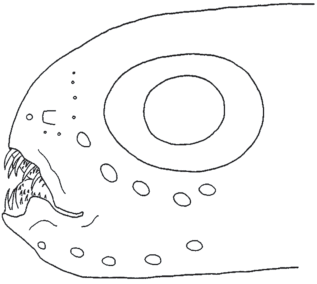
Голова Melanostigma japonicum Balushkin, sp. n. (по: Machida, Lee, 1997. Fig.2). Масштаб: 5 мм.
Melanostigma orientale (non Tominaga, 1971): Machida, Lee, 1997. Р. 1−5 (1 экз. из зал. Тоса, описание, сравнение с типовыми экземплярами M. orientale); Shinohara et al., 2001. Р. 328 (в списке видов зал. Тоса).
Голотип: BSKU 44840 − самка TL 102 мм, SL 98 мм со зрелой икрой, R/T Kotaka Maru, оттер-трал, зал. Тоса, 32°59′19″ с.ш. 133°35′52″ в.д., 700−720 м, 21.06.1988 г.
Диагноз. Челюсти укороченные, нижняя челюсть с изогнутым верхним краем в средней части и с латеральным утолщением. Часть крупных челюстных зубов наружного ряда направлены вперёд и не скрыты за губами. Супратемпоральная коммиссура неполная (нет медиальных сегментов); нет пор в темпоральном канале; нет боковой линии на теле; жаберных тычинок на 1-й жаберной дуге в наружном ряду 16; в грудном плавнике 7 лучей; общее число позвонков 100, из них туловищных 20, хвостовых 80; позвонки удлинённые ассиметричные, длина передней части центра туловищных позвонков (в районе 5−8-го позвонков) составляет 67−77% длины его задней части; окраска передней части головы тёмно-коричневая, хвост светлый.
Основные счётные признаки: D 95, A 80, P 7, vert. 20 + 80 = 100, C 10, жаберных тычинок на 1-й жаберной дуге в наружном ряду 16, до трёх неправильных рядов конических подвижных зубов у симфиза челюстей.
Тело низкое, сильно удлинённое и утончённое к концу, его максимальная высота располагается в передней части туловища. Кожа нежная полупрозрачная подвижная, с развитым желеобразным слоем, без чешуи. Голова небольшая, содержится 8.4 раза в TL, рыльный подъём очень крутой. Рот конечный, разрез рта слегка косой, не достигает вертикали переднего края глаза. Одна пара ноздрей с низкими трубками, окрашенными в чёрный цвет. Жаберное отверстие небольшое, расположено выше верхнего края грудного плавника примерно на расстояние своего вертикального диаметра. Жаберных лучей шесть. Жаберные тычинки простые, без ветвлений на концах.
Челюстные зубы конические подвижные, зубы внешнего ряда клыковидные, очень крупные, часть из них направлена вперёд и не скрыта за губами. Есть зубы на сошнике и нёбных костях.
Рентгенограмма. Позвонков до 1-го птеригиофора D 4. Птеригиофор, поддерживающий 1-й луч анального плавника, расположен на границе туловищного и хвостового отделов. Позвонки амфицельные, с высокими невральными дугами, снабжёнными крупными сочленовными отростками (zygapophyses). Презигапофизы позвонка налегают сверху на постзигапофизы предшествующего позвонка. Парапофизы видны обычно с 3-го позвонка. Epipleuralia и pleuralia на рентгенограмме не видны (скорее всего, они есть, поскольку присутствуют у близкого вида M. inexpectatum). С уростилярным позвонком (pu1 + u1) слиты две гипуральные пластинки: эпаксиальная и гипаксиальная, обе несут по четыре луча хвостового плавника. Epurale одно, слабо окостеневающее, кость не нависает над задней частью предуростилярнего позвонка (preurale 2). Сзади еpurale поддерживает два верхних луча С, которые в отличие от остальных лучей хвостового плавника значительно смещены вперёд примерно до вертикали середины уростиля. Нет верхнего остистого отростка на предуростилярном позвонке.
Сейсмосенсорная система. В инфраорбитальном и преоперкуло-мандибулярном каналах по пять пор, в обоих каналах первые поры немного мельче остальных. В супраорбитальном канале одна пора, расположенная немного впереди и медиальнее ноздри. Супратемпоральная коммиссура, скорее всего, прерванная посередине: через полупрозрачную кожу заметны лишь короткие слепые трубочки, отходящие с каждой стороны медиально от темпорального канала.
Измерения. В % SL: высота тела у начала анального плавника (включая спинной плавник) 7.2, антеанальное расстояние 34.1, постанальное расстояние (до конца лучей C) 69.9, расстояние от заднего края жаберного отверстия до середины ануса 21.8, длина грудного плавника 3.7, расстояние от нижнего края жаберного отверстия до основания 1-го луча грудного плавника 1.1, горизонтальный диаметр жаберного отверстия 0.8, длина головы (c) 12.3, ширина головы (wc) 6.2, высота головы через середину глаза (сh) 6.8; длина верхней челюсти (lmx) 2.1, длина рыла (ao) 2.3, ширина межглазничного расстояния (io) 1.1, продольный диаметр орбиты (o) 4.6. В % с: wc 51.3, сh 56.2, lmx 17.7, ao 19.0, io 9.1, продольный диаметр орбиты (o) 38.0.
Окраска. Тело светлое с полупрозрачной кожей; редко разбросанные меланофоры есть на верху головы; грудной и хвостовой плавники светлые; передняя часть головы тёмно-коричневая; трубочки ноздрей чёрные; жаберная, ротовая полости и перитонеум чёрные, жаберные лепестки белые.
Этимология. Название вида указывает на место находки голотипа у берегов Японии. Вместе с тем этим названием мне бы хотелось отметить также вклад японских ихтиологов в изучение меланостигм и выразить свою благодарность тем из них, кто оказал содействие в выполнении этой работы.
Распространение, биология, историческое расселение. Кроме голотипа, пойманного на глубине 700−720 м, к этому виду может относиться другой экземпляр, пойманный позднее в той же бух. Тоса в экспедиции на судне “Kotaka-maru” (33°05.2′−33°04.8′ с.ш. 133°43.0′−133°42.6′ в.д., глубина 820 м, 22.05.1997 г. (Shinohara et al., 2001)). Как мне любезно сообщил доктор Г. Шинохара, этот экземпляр TL 124 мм хранится в Национальном музее науки под номером NSMT-P55382. Голотип нового вида – самка со зрелой икрой, что установили ранее Машида и Ли (Machida, Lee, 1997). По моим измерениям, диаметр икры составляет 0.8−0.9 мм. Икринки эллипсовидной формы.
Новый вид относится к группе видов меланостигм, характеризующихся малыми размерами и ранним наступлением половой зрелости. Эти особенности Андерсон (Anderson, 1988, 1994) использовал в качестве особого (не морфологического) признака при построении кладограммы родственных отношений видов Melanostigma. По его представлениям, наиболее примитивными видами являются M. vitiazi и M. gelatinosum, а группа карликовых видов (M. bathium, M. atlanticum, M. inexpectatum, M. orientale и M. pammelas − эти виды приведены здесь в порядке ответвлений на кладограмме), полная длина которых не превышает 140 мм11, венчает филогенетическое древо рода (Anderson, 1994. Fig. 54 ). В соответствии с филогенетической схемой автор предложил сценарий происхождения и исторического расселения меланостигм, согласно которому род возник от гимнелинового предка (от названия подсемейства Gymnelinae, куда относится род Melanostigma), проникшего в Южное полушарие из Северной Пацифики. Потомками его здесь стали как более крупные и примитивные M. vitiazi и M. gelatinosum22, так и мелкая форма, ближе всего к которой из современных видов стоит M. bathium, обитающий в водах Перуанского течения. Эта карликовая форма расселялась обратно в Северное полушарие, где в ходе дальнейшей редуктивной эволюции дала виды в Атлантике и Пацифике. По мнению Андерсона (Anderson, 1988, 1994), путь меланостигм из Северной Пацифики в Северную Атлантику можно объяснить в рамках трансокеанской гипотезы Андрияшева (Andriashev, 1986), обоснованной ранее на липаровых видах рода Paraliparis. Суть гипотезы заключалась в том, что расселение этих рыб происходило из Северной Пацифики вдоль тихоокеанских побережий американских континентов в обход Южной Америки с юга, а уже из Магелланова региона либо вновь в Северное полушарие вдоль Срединно-Атлантического хребта, либо на северо-восток к берегам Африки или к Кергеленскому архипелагу вдоль океанических поднятий талассобатиального типа. Неоднократно возвращаясь к своей гипотезе, Андрияшев (1990, 2003) допускал, что с её помощью можно объяснить исторические миграции не только паралипарисов, но и вторично глубоководных представителей других семейств и родов, в том числе вторично пелагических родов Psednos (Liparidae) и Melanostigma. По моим представлениям (Балушкин, 2012), более простым и коротким путём попадания липаровых из Северной Пацифики в Северную Атлантику мог быть всё же Панамский морской пролив, существенное обмеление которого и образование наземного моста между американскими континентами произошло лишь в плиоцене (3−4 млн лет назад). Отрицать такой же путь расселения для меланостигм нет каких-либо серьёзных оснований. Проникнув в Северную Атлантику, меланостигмы могли расселяться на юг не только американским маршрутом, который хорошо обоснован в литературе на примере бельдюговых и морских слизней (Regan, 1914; Andriashev, 1986; Anderson, 1988, 1994; Андрияшев, 1990, 1993, 2003; Andriashev, Stein, 1998; Балушкин, Воскобойникова, 2008; Балушкин, 2012). Подобно североатлантическим по своему происхождению таксонам, в частности родам Gaidropsarus и Merluсcius (Gadiformes), они могли расселяться также европейско-западноафриканским путём, достигая южной оконечности Африки. Подробно эти два маршрута для хеков были обсуждены, например, в недавних обзорах с привлечением молекулярных данных (Quinteiro et al., 2000; Grant, Leslie, 2001). Что же касается причин и места происхождения рода Melanostigma, то, по моему мнению, последовательность дивергенции была прямо обратной тому, что предполагал Андерсон (Anderson, 1988, 1994). Вероятнее всего, предком меланостигм была не крупная форма из Южного полушария, а мелкая форма, которая возникла в результате педоморфоза от предка, обитавшего на родине бельдюговых рыб − в Северной Пацифике. Из современных видов к предку ближе всего стоит M. pammelas, сохранивший в наибольшей степени следы пумилистической дегенерации (термин, предложенный Городковым (1984)). У этого вида не только отсутствуют боковые линии тела и редуцирована супратемпоральная коммиссура, что свойственно также другим северным карликовым видам, но и исчез postcleithrum. Расселение меланостигм на юг, проходившее по обеим сторонам Тихого океана и, вероятно, только по восточной стороне Атлантического океана, сопровождалось в некоторых филогенетических ветвях укрупнением размеров видов, т.е. развитие шло путём геронтоморфоза. Этот путь филогенеза, как отмечал ещё Де Бир (DeBeer, 1930), характеризуется небольшими морфологическими изменениями видового уровня, что мы и наблюдаем у меланостигм. Необходимо подчеркнуть, что возникновение и последующая диверсификация рода Melanostigma от предковой формы с малыми размерами тела полностью согласуется с общей закономерностью эволюции позвоночных, выраженной в законе (правиле) Копа (подробнее о современной трактовке закона см. обзор: Stanley, 1973). Удлинение онтогенеза могло стать причиной возврата (инволюции, реверсии) признаков, которые были присущи предку меланостигм. В частности, это могло привести как к восстановлению боковых линий туловища, которые отсутствуют у карликовых видов, но найдены у крупных видов (M. meteori, M. kharini, M. olgae и M. vitiazi) (Парин, 1979; Балушкин, Орловская, 2019), так и обретению вновь плотной (а не желеобразной) кожи на теле (M. vitiazi).
Сравнительные замечания. Новый вид пополняет группу многопозвонковых пелагических бельдюг подрода Melanostigma, у которых 93 позвонка являются нижним пределом видовой изменчивости (M. atlanticum, M. inexpectatum, M. kharini и M. orientale). Пока трудно судить, насколько эта группа является естественной. Приводимая ниже таблица призвана помочь в идентификации нового вида в тихоокеанских водах.
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА ВИДОВ MELANOSTIGMA ТИХОГО ОКЕАНА И СОПРЕДЕЛЬНЫХ АКВАТОРИЙ ИНДИЙСКОГО И ЮЖНОГО ОКЕАНОВ
1 (2) Кожа на теле плотная, неподвижная и непрозрачная (подрод Bandichthys). На рыле и у симфиза нижней челюсти имеются крупные невромасты. Туловищная боковая линия, составленная из трёх серий поверхностных невромастов (предорсальной, дорсолатеральной и медиолатеральной), внешне хорошо различима (море Банда, Индо-Вестпацифика) …………… M. vitiazi Parin
2 (1) Кожа тонкая подвижная и полупрозрачная (подрод Melanostigma). Нет крупных невромастов на рыле и у симфиза нижней челюсти. Боковая линия отсутствует или поверхностные невромасты туловищных серий плохо различимы (M. kharini).
3 (10) Позвонков менее 92.
4 (7) Темпоральная пора имеется. Верх головы и передней части тела покрыт мелкими тёмными пятнами. Хвостовых позвонков 62−64 (точное число неизвестно для M. flaccidum).
5 (6) Хвост заметно утончается к концу. Длина головы 4.9−6.1, высота тела 9.5−10.0 раз в TL. Длина грудного плавника составляет более 1/3 длины головы (тихоокеанское побережье Южной Америки, Магелланов пролив). …….... M. gelatinosum Günther
6 (5) Хвост высокий и округлый на конце. Длина головы и высота тела 6.6 раза в TL. Длина грудного плавника составляет 1/3 длины головы (восточное побережье Южного острова, Новая Зеландия). .…………………...............M. flaccidum Waite
7 (4) Темпоральная пора отсутствует. Окраска головы и тела, если и имеется, то монотонная, без пятен. Хвостовых позвонков 66−72.
8 (9) Голова и конец хвоста светлые. Позвонков 84−88. Супратемпоральная коммиссура имеется (тихоокеанское побережье Южной Америки, на север до Галапагосских островов) ………..…..…..…..…..…..…..……M. bathium Bussing
9 (8) Голова и конец хвоста тёмные, пигментированные. Позвонков 86−91. Супратемпоральная коммиссура отсутствует (тихоокеанское побережье Северной Америки от Британской Колумбии на юг до зал. Техуантепек) …..…..…….…………. …………….……..…….……............M. pammelas Gilbert
10 (3) Позвонков более 92.
11 (16) Супратемпоральная коммиссура с разрывом посередине или вообще отсутствует.
12 (13) Жаберных тычинок внешнего ряда на 1-й жаберной дуге 12 (возможно, до 14). Антеанальное расстояние 28.5−30.3% TL. Диаметр глаза 20.0−29.4% длины головы (тихоокеанское побережье центральной Японии) ….….….… M. orientale Tominaga
13 (12) Жаберных тычинок наружного ряда 1-й жаберной дуги 16 (возможно, до 18−19). Антеанальное расстояние 32.0−34.5% TL. Диаметр глаза 35.8−38.0% длины головы.
14 (15) Тело низкое, прогонистое, его максимальная высота 6.9% SL. Передние челюстные зубы очень крупные (до 5.8% длины головы). Нижняя челюсть с резким изгибом верхнего края в центре и с латеральным утолщением (тихоокеанское побережье юга Японии) …………………………M. japonicum Balushkin sp. n.
15 (14) Тело сравнительно высокое, его максимальная высота 13.0% SL. Длина увеличенных челюстных зубов внешнего ряда до 2.3% длины головы. Нижняя челюсть без резкого изгиба верхнего края и без латерального утолщения (Западная экваториальная Пацифика) ………………… M. inexpectatum Parin
16 (11) Супратемпоральная коммиссура законченная (полная), без разрывов (хребет Геракл, тихоокеанский сектор Южного океана) ………………..…..…M. kharini Balushkin et Moganova
БЛАГОДАРНОСТИ
Автор благодарен японским ихтиологам Хиромицу Эндо (Dr. Hiromitsu Endo, Kochi University) и Рио Мисава (Ryo Misawa, Kyoto University) за предоставление голотипа нового вида и его рентгено- и фотосъёмку; Генто Шинохара (Dr. Gento Shinohara, National Museum of Nature and Science) и Кацуо Сакамото (Dr. Kazuo Sakamoto, University of Tokyo) − за присланный рентгеноснимок голотипа M. orientale.
Автор признателен М.В. Орловской (ЗИН РАН) за обсуждение и помощь в оформлении работы, С.А. Евсеенко (ИО РАН) − за ценные замечания и советы при чтении рукописи статьи.
Список литературы
Андрияшев А.П. 1990. О вероятности трансокеанского (неарктического) расселения вторичноглубоководных видов рыб бореально-тихоокеанского происхождения до глубин северной Атлантики и Арктики (на примере семейства Liparididae) // Зоол. журн. Т. 69. Вып. 1. С. 61–67.
Андрияшев А. П. 1993. О валидности рода Psednos Barnard (Scorpaeniformes, Liparidae) и его антитропическом ареале // Вопр. ихтиологии. Т. 33. № 1. С. 5−15.
Андрияшев А.П. 2003. Липаровые рыбы (Liparidae, Scorpaeniformes) Южного океана и сопредельных вод // Исследования фауны морей Т. 53 (61). СПб.: Изд-во ЗИН РАН, 474 с.
Балушкин А.В. 2012. Volodichthys gen. nov. − новый род примитивных липаровых рыб (Liparidae: Scorpaeniformes) южного полушария с описанием нового вида V. solovjevae sp. nov. из моря Содружества (Антарктика) // Вопр. ихтиологии. Т. 52. № 1. С. 1–10.
Балушкин А.В., Воскобойникова О.С. 2008. Ревизия рода Genioliparis Andriashev et Neelov (Liparidae, Scorpaeniformes) c описанием нового вида G. kafanovi sp. n. из моря Росса (Антарктика) // Там же. Т. 48. № 2. С. 149–157.
Балушкин А.В., Моганова М.В. 2017. Новый вид пелагической бельдюги рода Melanostigma (Zoarcidae) из западной Антарктики // Там же. Т. 57. № 2. С. 131–136.
Балушкин А.В., Моганова М.В. 2018. Melanostigma kharini sp. nov. (Zoarcidae) − новый вид пелагической бельдюги с хребта Геракл (Антарктическо-Южнотихоокеанское поднятие) // Там же. Т. 58. № 2. С. 139–143.
Балушкин А.В., Орловская М.В. 2019. Melanostigma meteori sp. n. (Zoarcidae) – новый вид пелагической бельдюги с банки Метеор (юго-восточная Атлантика) с замечаниями о полимеризации боковых линий тела у бельдюговых рыб // Там же. Т. 59. № 2. С. 127–136.
Балушкин А.В., Шейко Б.А., Природина В.П. 2012. Каталог фондовой коллекции Зоологического института РАН. Класс костистые рыбы (Osteichthyes). Отряд Окунеобразные (Рerciformes). Подотряд Zoarcoidei, семейства: Bathymasteridae, Zoarcidae, Cryptacanthodidae, Ptilichthyidae и Zaproridae. Подотряд Icosteoidei, семейство Icosteidae // Исследования фауны морей. Т. 71 (79). СПб.: Изд-во ЗИН РАН, 195 с.
Городков К.Б. 1984. Олигомеризация и эволюция систем морфологических структур. 2. Олигомеризация и уменьшение размеров тела // Зоол. журн. Т. LXIII. Вып. 12. С. 1765−1778.
Парин Н.В. 1979. Melanostigma (Bandichthys) vitiazi – новая глубоководная рыба (Melanostigmatidae, Osteichthyes) из моря Банда // Вопр. ихтиологии. Т. 19. Вып. 1 (114). С. 167–170.
Anderson M.E. 1988. Studies on the Zoarcidae (Teleostei: Perciformes) of the southern hemisphere. I. The Antarctic and subantarctis regions // Antarct. Res. Ser. 47. P. 59–113.
Anderson M.E. 1990. Zoarcidae // Fishes of the Southern Ocean / Eds. Gon O., Heemstra P.C. Grahamstown, S. Africa: J.L.B. Smith Inst. Ichthyol. P. 257−276.
Anderson M.E. 1994. Systematics and osteology of the Zoarcidae (Teleostei: Perciformes) // Ichthyol. Bull. J.L.B. Smith Inst. Ichth. V. 60. P. 1–120.
Andriashev A.P. 1986. Review of the snailfish genus Paraliparis (Scorpaeniformes: Liparididae) of the Southern Ocean // Theses Zool. V. 7. Koenigstein: Koeltz Sci. Books, 204 p.
Andriashev A.P., Stein D.L. 1998. Review of the snailfish genus Careproctus (Liparidae, Scorpaeniformes) in Antarctic and adjacent waters // Contrib. Sci. Los Angeles County Mus. № 470. P. 1–63.
Balushkin A.V., Sheiko B.A., Fedorov V.V. 2011. Catalog of the archival collection of the Zoological Institute, Russian Academy of Sciences: Class Osteichthyes (Bony fishes), Order Perciformes, Family Zoarcidae // J. Ichthyol. V. 51. № 10. P. 950–1034.
Bussing W.A. 1965. Studies of the midwater fishes of the Peru-Chile Trench // Antarct. Res. Ser. V. 5. P. 219–222.
DeBeer G.R. 1930. Embryology and evolution. Oxford: Crarendon, 116 p.
Grant W.S., Leslie R.W. 2001. Inter-ocean dispersal is an important mechanism in the zoogeography of hakes (Pisces: Merluccius spp.) // J. Biogeography. V. 28. P. 699−721.
Hatooka K. 2002. Zoarcidae // Fishes of Japan with pictorial keys to the species. V. 2 / Ed. Nakabo T. Second edition. Tokyo: Tokai Univ. Press. P. 1026−1044.
Hatooka K. 2013. Zoarcidae // Fishes of Japan with pictorial keys to the species. V. 2 / Ed. Nakabo T. Third edition. Tokyo: Tokai Univ. Press. P. 1220−1251.
Machida Y., Lee Ch.-L. 1997. Notes on a rare deep-see eelpout, Melanostigma orientale, from Tosa Bay, southern Japan (Perciformes, Zoarcidae) // Bull. Mar. Sci. Fish. Kochi Univ. № 17. P. 1−5.
McAllister D.E., Ress E.J.S. 1964. A revision of the eelpout genus Melanostigma with a new genus and with comments on Maynea // Bull. Nat. Mus. Can. V. 199. P. 85–110.
Quinteiro J., Vidal R., Rey-Méndez M. 2000. Phylogeny and biogeographic history of hake (genus Merluccius), inferred from DNA control-region sequences // Mar. Biol. V. 136. P. 163−174.
Regan C.T. 1914. Fishes // Nat. Hist. Rept. Brit. Antarct. (“Terra Nova”) Exp. 1910. Zool. V. 1. № 1. P. 1−54.
Robin C.R., Ray G.C. 1986. A field guide to Atlantic coast fishes of North America. Boston: Houghton Mifflin Comp., 354 p.
Shinohara G., Endo H., Matsuura K. et al. 2001. Annotated checklist of the deepwater fishes from Tosa Bay, Japan // Nat. Sci. Mus. Monographs. Tokyo. № 20. P. 283−343.
Stanley S.M. 1973. An explanation for Cope’s Rule // Evolution. V. 27. № 1. P. 1−26.
Tominaga Y. 1971. Melanostigma orientale, a new species of zoarcid fish from Sagami Bay and Suruga Bay, Japan // Jpn. J. Ichthyol. V. 18. № 4. P. 151–156.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии