

Вопросы ихтиологии, 2019, T. 59, № 3, стр. 286-291
Зимнее распределение рыб в русловой яме реки Иртыш
А. Д. Мочек 1, *, Э. С. Борисенко 1, Д. С. Павлов 1
1 Институт проблем экологии и эволюции РАН – ИПЭЭ
Москва, Россия
* E-mail: amochek@yandex.ru
Поступила в редакцию 09.10.2018
После доработки 09.10.2018
Принята к публикации 11.10.2018
Аннотация
С помощью компьютеризированных гидроакустических комплексов исследовано зимнее распределение рыб на крупнейшей русловой яме р. Иртыш. Установлено, что подо льдом на русловой яме образуются скопления рыб, местоположение, плотность, численность и размерный состав которых претерпевают в течение зимы существенные изменения. В начале зимы плотность и численность скоплений рыб выше, чем в предвесенний период. Распределение рыб по площади ямы меняется из месяца в месяц, но приуроченность большинства рыб к толще воды и приповерхностным горизонтам остаётся неизменной. На акватории ямы рыбы повсеместно объединены в динамичные агрегации. Бóльшая часть рыб подо льдом представлена малоразмерными особями карповых (Cyprinidae),
Феномен концентрации рыб на глубоководных участках водоёмов неоднократно отмечен в литературе (Вотинов, 1958; Никольский, 1963; Иоганзен, 1972; Поддубный, Малинин, 1988). Наши исследования на нижнем Иртыше, в Горнослинкинской русловой яме, позволили детально изучить распределение рыб, закономерности формирования их скоплений в период открытой воды (Павлов, Мочек, 2005; Pavlov et al., 2006; Экология рыб …, 2006; Павлов и др., 2011). Известно, что с наступлением зимы распределение и поведение рыб во внутренних водоёмах умеренных и высоких широт существенно меняется, однако особенности подлёдного распределения рыб на крупных реках изучены недостаточно. В малых реках льдообразование является критическим фактором для выживания молоди лососёвых (Salmonidae) вследствие образования донного льда (Brown, 1999; Stiсkler et al., 2006; Stiсkler, 2008; Brown et al., 2011). Поэтому зимой рыбы в таких водотоках концентрируются на участках с минимальным течением и слабой степенью промерзания (Brown, 1999; Stiсkler et al., 2006; Huusko et al., 2007; Stiсkler, 2008; Brown et al., 2011). В озёрах в течение подлёдного периода численность и размерный состав рыб в скоплениях и их размещение изменяются.
В лимнических водоёмах после ледостава распределение рыб определяется температурным режимом разных горизонтов, их кислородной обеспеченностью, особенностями оборонительно-пищевых отношений гидробионтов (Щербаков, 1967; Eckmann, 1995; Steinhart, Wurtsbaugh, 1999). Универсальной особенностью подлёдного распределения рыб в озёрах является образование скоплений на фоне сохранения суточной ритмики их вертикальных кочёвок (Павлов и др., 1991; Presnyakov, Borisenko, 1993; Eckmann, 1995; Steinhart, Wurtsbaugh, 1999; Jurvelius, Marjoma, 2008).
По известным нам литературным данным, специальные ихтиологические исследования русловых ям крупных рек в зимнее время ранее не проводились. Материалы, отражающие закономерности освоения рыбами русловых ям подо льдом, необходимы для понимания экологической роли этих участков речной системы и представляют актуальность в практическом отношении (Павлов, Мочек, 2005).
Цель работы – изучить особенности зимнего распределения рыб на крупной русловой яме в подлёдный период с декабря по март: выявить динамику плотности, размерного состава и численности рыб, распределение скоплений рыб по площади ямы и глубинам, изучить их агрегации.
МАТЕРИАЛ И МЕТОДИКА
Горнослинкинская русловая яма р. Иртыш расположена в координатах 58°44′ с.ш. 68°41′ в.д.; образована по руслу реки, имеет сложную форму с двумя поворотами на 90°; её протяжённость 1.5 км, площадь акватории превышает 58 га. Скорость течения на стрежне достигает 1.5 м/с, а перепады глубин − более 40 м. Вследствие сильного течения, сложной формы русла и срединного поднятия дна по стрежню ямы образуются мощные водовороты с вертикальной осью вращения, у берегов и на больших глубинах формируются зоны гидравлической тени.
Среди рыбного населения русловой ямы по численности доминируют Cyprinidae (более 98% – преимущественно молодь): Leuciscus idus, Abramis brama, Rutilus rutilus, Leucaspius delineates, Carrasius carrasius. Рыбы других фоновых семейств – Percidae (Perca fluviatilis, Stizostedion lucioperca, Gymnocephalus cernuus), Acipenseridae (Acipenser ruthenus), Esocidae (Esox lucius), Gadidae (Lota lota) − встречаются значительно реже. Видовая принадлежность рыб установлена на основе сетных обловов, а также оригинального гидроакустического метода – анализа огибающей эхосигнала (Pavlov еt al., 2010).
Изучение особенностей жизнедеятельности рыб на русловых ямах, и тем более подо льдом, требует применения специальных методик. В последние десятилетия для подлёдных ихтиологических исследований широко используется гидроакустическая аппаратура в сочетании с контрольным ловом рыб (Presnyakov, Borisenko, 1993; Eckmann, 1995; Steinhart, Wurtsbaugh, 1999; Jurvelius et al., 2000, 2011; Jurvelius, Marjoma, 2008). Настоящая работа основана на гидроакустическом зондировании акватории как рациональном и предпочтительном на сегодняшний день способе изучения рыбного населения крупных естественных водотоков.
Исследования проводили с декабря 2006 по март 2007 гг., т.е. в период устойчивого ледового покрова на Иртыше. Основная часть работы выполнена с использованием компьютеризированного комплекса вертикального зондирования “Аскор” (ООО “Промгидроакустика”, Россия), который включает эхолот с частотой излучения 50 и 200 кГц, передающие компоненты, систему фиксации, анализа и преобразования эхосигналов. Приёмно-передающие антенны размещали подо льдом. Для этого бурили 18 лунок диаметром 20 см по стрежневой трансекте и 6 лунок на поперечной трансекте в верхней и нижней оконечностях ямы. Отражённый акустической целью сигнал эхолота записывался в цифровой форме на твёрдый диск ноутбука. В итоге дистанционным путём определяли плотность скоплений и размер обнаруженных рыб, их местоположение по горизонтальным и вертикальным векторам. Дальнейшая компьютерная обработка позволяла с помощью специальных программ создавать таблицы, графические отображения и планшеты распределения рыб в водоёме.
Для наблюдения за агрегациями рыб подо льдом, фиксации их перемещений и межгруппового обмена особями применяли гидролокатор кругового обзора FSS-3300 (“Symrad”, Норвегия). Антенну локатора устанавливали на глубине 10 м в срединной части ямы через прорубь размером 0.5 × 0.5 м. С помощью этого прибора зондировали толщу воды до дна и таким образом регистрировали всех рыб в границах диаграммы направленности антенны – 20°. Продолжительность наблюдения составила 1 ч, причём каждые 5 мин изображение останавливали для компьютерной фиксации местоположения и перемещения рыб. Изображение на мониторе гидролокатора записывали на жёсткий диск с обозначениями расположения агрегаций рыб и одиночных особей, береговых отрогов и срединного поднятия дна на русловой яме. По визуализированным перемещениям рыб оператор в дальнейшем строил этограммы состояния скоплений.
Всего проведено шесть серий гидроакустического зондирования по каждой из трансект с помощью эхолота и одно наблюдение на стрежне с помощью гидролокатора. Исследования выполняли в светлое время суток.
РЕЗУЛЬТАТЫ
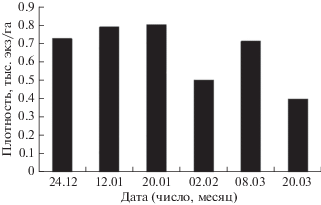
В течение всей зимы на русловой яме подо льдом постоянно находится скопление рыб, но его местоположение и плотность, а также численность и размерный состав рыб меняются. Наибольшая плотность скопления отмечена в разгар зимы (декабрь−январь), а к концу зимы (февраль−март) наблюдается тенденция её уменьшения (рис. 1). Соответственно, численность скопления рыб на яме претерпевала значительные колебания в течение всей зимы при сохранении общего тренда на снижение к началу весны. Основу скопления рыб на яме постоянно составляли особи длиной 8−16 см. Вместе с тем доля особей минимального (5−7 см), а также среднего (17−32 см) и крупного (>32 см) размера менялась от месяца к месяцу (таблица).
Рис. 1.
Средняя плотность скоплений рыб на акватории Горнослинкинской ямы в зимний период с декабря 2006 по март 2007 гг.
Общая численность рыб, плотность скопления и соотношение разных размерных групп на акватории Горнослинкинской русловой ямы в зимний период, %
| Дата | Численность, экз. | Плотность, экз/га | Длина рыб, см | ||||
|---|---|---|---|---|---|---|---|
| средняя | максимальная | 5–7 | 8–16 | 17–32 | >32 | ||
| 24.12.2006 | 22 100 | 727 | 9600 | 43 | 42 | 8 | 7 |
| 12.01.2007 | 25 700 | 789 | 9728 | 37 | 39 | 18 | 6 |
| 20.01.2007 | 25 900 | 795 | 33 000 | 32 | 49 | 14 | 5 |
| 02.02.2007 | 16 200 | 498 | 5500 | 26 | 42 | 23 | 9 |
| 08.03.2007 | 23 200 | 713 | 6600 | 24 | 53 | 15 | 8 |
| 20.03.2007 | 13 200 | 404 | 8400 | 28 | 56 | 11 | 5 |
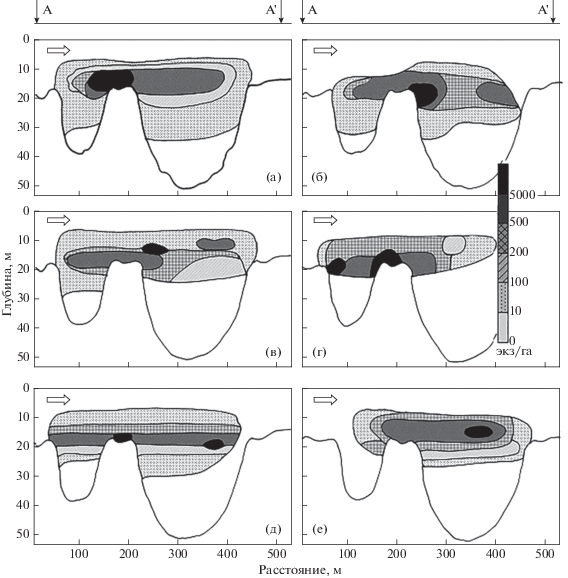
Освоение рыбами пространства русловой ямы отражает динамичность процесса их распределения (рис. 2, 3) В начале зимы (декабрь−январь) скопление рыб занимает наибольшее пространство русловой ямы как по площади акватории (рис. 2а−2в), так и по глубине (рис. 3а−3в). В последующий зимний период горизонтальные и вертикальные пределы массового размещения рыб сужаются. Если в декабре и январе нижняя граница скопления рыб достигала глубины 30−35 м, то в феврале лишь 20 м (рис. 3г). В марте рыбы размещались главным образом до глубины 25−30 м (рис. 3д, 3е). По мере прохождения зимнего периода участок максимальной плотности скопления смещался по направлению к стрежню реки (2д). В наибольшей степени тенденция перехода скопления на стрежень проявилась в конце марта (рис. 2е). Одновременно наблюдался подъём основной массы рыб на глубины не более 25 м (рис. 3е). На протяжении всей зимы максимальная плотность скопления была приурочена к участкам неоднородного донного рельефа – срединному поднятию и прибрежному отрогу ямы.
Рис. 2.
Планшеты размещения рыб на акватории Горнослинкинской русловой ямы в подлёдный период: а − 24.12.2006 г., б − 12.01.2007 г., в − 20.01.2007 г., г − 02.02.2007 г., д − 08.03.2007 г., е − 20.03.2007 г.; (⇒) − направление течения реки; А−А' – стрежневая трансекта.
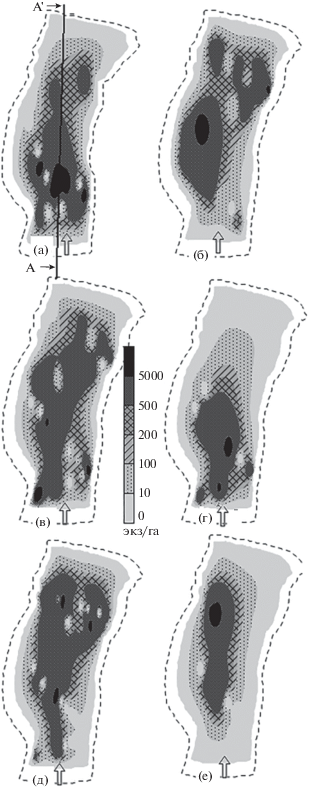
Рис. 3.
Вертикальная структура скоплений рыб на Горнослинкинской русловой яме в зимний период: а – 24.12.2006 г., б – 12.01.2007 г., в – 20.01.2007 г., г – 02.02.2007 г., д – 08.03.2007 г., е – 20.03.2007 г.; обозначения см. на рис. 2.
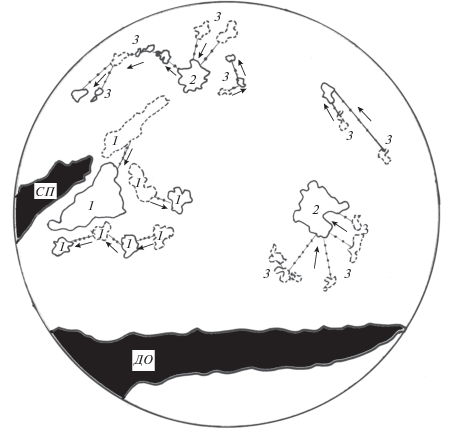
Скопления рыб на русловой яме представлены динамичными агрегациями (рис. 4). Одиночные рыбы, а также группы постоянно меняют своё местоположение, причём между ними наблюдается обмен особями. У береговых отрогов и на срединном поднятии ямы могут располагаться придонные агрегации, а в толще воды − пелагические группы и одиночные особи. Агрегации рыб распадаются и формируются вновь, увеличиваются в размерах за счёт соседних групп.
Рис. 4.
Придонные (1) и пелагические (2) агрегации рыб и одиночные особи (3), по данным регистрации сканирующим гидролокатором кругового обзора FSS-3300 срединного поднятия (СП) и донных отрогов (ДО) Горнослинкинской русловой ямы в марте 2007 г.; (- -) – первоначальное положение скоплений, (—) – итоговое положение скоплений спустя 1 ч, (→) – направление перемещений рыб.
ОБСУЖДЕНИЕ
Зимнее распределение рыб на русловой яме характеризуется динамичными чертами: происходят изменения плотности, численности, размерного состава и местоположения скопления. Максимальная плотность скопления рыб наблюдается в начале зимы, однако на протяжении зимы она заметно снижается, падая до минимума весной. В первую половину зимы на акватории ямы численность рыб в скоплении была максимальной, а в конце зимы она уменьшилась практически в два раза. Вероятно, к весне значительная часть молоди и даже крупных рыб погибают, а часть производителей покидают участки зимовки, мигрируя к местам размножения и нагула. В течение зимних месяцев наблюдаются широкие перемещения рыб по площади русловой ямы, хотя основная масса рыб предпочитает неоднородности донного рельефа. Вертикальное распределение рыб претерпевает лишь незначительные изменения с сохранением очевидной приуроченности основной массы рыб к пелагическим горизонтам.
Молодь в пелагиали образует подвижные агрегации с переменным составом. На протяжении ограниченного времени агрегации распадаются, многие особи их покидают, объединяясь в малые группы, но вскоре рыбы образуют новые агрегации (рис. 4). Этот процесс напоминает известный эффект стохастического движения в кормящихся стаях рыб (Радаков, 1972). Столь интенсивная изменчивость объединений молоди объясняется разными причинами, в том числе воздействием крайне неоднородных гидравлических условий на русловой яме.
Скопления рыб на акватории русловых ям крупных рек наблюдаются всесезонно – при открытой воде и подо льдом. В их составе встречаются все размерно-возрастные группы – от ранней молоди до производителей, хотя весной и в начале лета абсолютно доминирует молодь (Pavlov et al., 2006). Концентрирующий эффект русловых ям для рыб как зимой, так и летом определяется внешними условиями этого своеобразного биотопа. Вследствие мощного течения, перепадов глубины и резких поворотов русла на яме формируется сложная гидравлическая структура потока с разнонаправленными водоворотами. На этот участок течением реки заносится покатная молодь, активно подходят производители. Рыбы, попавшие на русловые ямы, подолгу остаются в таких местообитаниях. С одной стороны, водовороты на длительное время дезориентируют и удерживают рыб в пелагиали (Павлов, 1979). С другой стороны, у отрогов ямы и на больших глубинах формируются затишные зоны, где рыбы могут находиться длительное время, применяя при этом минимум усилий. Кроме того, интенсивное перемешивание водных слоёв благоприятствует улучшению кислородного и термального режимов местообитания. Хищники на русловой яме находят обильную кормовую базу, а их потенциальные жертвы (молодь рыб) приобретают дополнительную возможность защиты. Эти особенности биотопа ямы в целом способствуют концентрации и длительному пребыванию здесь множества рыб.
Таким образом, исследования на Горнослинкинской русловой яме показали, что зимой здесь формируются скопления рыб, основу которых составляют особи малых размеров – от 8 до 16 см. На протяжении зимы меняются местоположение, плотность, численность и размерный состав скоплений. Распределение рыб по площади ямы изменчиво, но приуроченность большинства особей к толще воды и приповерхностным горизонтам остаётся постоянной. Скопления рыб на яме образуются вследствие водоворотной структуры течения на резких перепадах глубин и поворотах русла. Молодь в массе попадает на яму с транзитным потоком и длительное время остаётся дезориентированной в водоворотах либо держится на затишных участках биотопа. На акватории русловой ямы формируются разные по размеру агрегации рыб, между которыми происходит обмен особями.
Список литературы
Вотинов Н.П. 1958. Осетровые рыбы Обского бассейна. Тюмень: Тюмен. книж. изд-во, 42 с.
Иоганзен Б.Г. 1972. Зональное и биотопическое распределение рыб в долине Оби // Биологические ресурсы поймы Оби. Новосибирск: Наука. С. 270−291.
Никольский Г.В. 1963. Экология рыб. М.: Высш. шк., 368 с.
Павлов Д.С. 1979. Биологические основы управления поведением рыб в потоке воды. М.: Наука, 319 с.
Павлов Д.С., Мочек А.Д. 2005. Рыбные ресурсы Обь-Иртышского бассейна и роль зимовальных ям в их сохранении // Фундаментальные основы управления биологическими ресурсами. М.: Т-во науч. изд. КМК. С. 132−138.
Павлов Д.С., Гусар А.Г., Михеев В.Н. и др. 1991. Пространственное распределение и биология плотвы в пелагиали озера Глубокого в подледный период // Бюл. МОИП. Отд. биол. Т. 96. Вып. 2. С. 95−102.
Павлов Д.С., Мочек А.Д., Борисенко Э.С. и др. 2011. Распределение рыб в пойменно-русловом комплексе Нижнего Иртыша // Биология внутр. вод. № 2. С. 71–79.
Поддубный А.Г., Малинин Л.К. 1988. Миграции рыб во внутренних водоемах. М.: Агропромиздат, 223 с.
Радаков Д.В. 1972. Стайность рыб как экологическое явление. М.: Наука, 174 с.
Щербаков А.П. 1967. Озеро Глубокое. М.: Наука, 377 с.
Экология рыб Обь-Иртышского бассейна. 2006 / Под ред. Павлова Д.С., Мочека А.Д. М.: Т-во науч. изд. КМК, 506 с.
Brown R. 1999. Fall and early winter movements of cutthroat trout, Oncorhynchus clarki, in relation to water temperature and ice conditions in Dutch Creek, Alberta // Environ. Biol. Fish. V. 55. P. 359–368.
Brown R., Wayne A., Steven F. 2011. A primer on winter, ice, and fish: what fisheries biologists showed know about winter ice processes and stream–dwelling fish // Fisheries. V. 36. № 1. P. 8−28.
Eckmann R. 1995. Abundance and horizontal distribution of Lake Constance pelagic whitefish (Coregonus lavaretus L.) during winter // Advanc. Limnol. V. 46. P. 249−259.
Huusko A., Greenberg L., Stickler M. et al. 2007. Life in the ice lane: the winter ecology of stream salmonids // River Res. Applicat. V. 23. P. 469−491.
Jurvelius J., Marjoma T. 2008. Night, day, sunrise, sunset: do fish under snow and ice recognize the difference? // Freshwat. Biol. V. 53. P. 2287–2294.
Jurvelius J., Lilja J., Hirvonen E. et al. 2000. Under ice density in fish stocks in lakes: gill netting, seining, trawling and hydroacouistics as sampling methods // Aquat. Liv. Resour. V. 13. P. 403–408.
Jurvelius J., Kolari I., Leskela A. 2011. Quality and status of fish stocks in lakes: gillnetting, seining, trawling and hydroacoustics as sampling methods // Hydrobiologia. V. 660. P. 29–36.
Pavlov D.S., Mochek A.D., Borisenko E.S. et al. 2006. Biological significance of the Gornoslinkinskaya riverbed depression in the Irtysh // J. Ichthyol. V. 46. Suppl. 2. P. S125−S133.
Pavlov D.S., Mochek A.D., Borisenko E.S., Degtev A.I. 2010. Hydroacoustic investigation of taxonomic composition and of vertical distribution of fish in the riverbed depression // Ibid. V 50. №. 11. P. 969−976.
Presnyakov V.V., Borisenko E.S. 1993. The study of fish behaviour under ice of Lake Glubokoe by means of scanning sonar // Fish. Res. V. 15. P. 323–329.
Steinhart G.B., Wurtsbaugh W.A. 1999. Under-ice diel vertical migrations of Oncorhynchus nerka and their zooplankton prey // Can. J. Fish. Aquat. Sci. V. 56. № 1. P. 152–161.
Stickler M. 2008. Anchor ice formation and habitat choice of Atlantic salmon (Salmo salar L.) parr in steep streams: Th. Ph.D. Norw. Univ. Sci. Techn., 273 p
Stickler M., Enders E.C., Pennell C.J. et al. 2006. Alfredsen. Gradient related movement of Atlantic salmon parr during a freeze up event in a small natural river // Proc. 18-th IAHR Int. Symp. Ice. P. 49−61.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии