

Вопросы ихтиологии, 2019, T. 59, № 4, стр. 416-427
Видовой состав, распределение и некоторые аспекты биологии рыб рода Lycodes (Zoarcidae) в море Лаптевых в 2014 году
Е. В. Смирнова 1, *, О. В. Карамушко 1, Н. В. Чернова 2
1 Мурманский морской биологический институт Кольского научного центра РАН – ММБИ КНЦ РАН
Мурманск, Россия
2 Зоологический институт РАН – ЗИН
Санкт-Петербург, Россия
* E-mail: smirnova@mmbi.info
Поступила в редакцию 07.09.2018
После доработки 13.02.2019
Принята к публикации 25.02.2019
Аннотация
По материалам экспедиционных работ в море Лаптевых в 2014 г. представлены данные о встречаемости, пространственно-батиметрическом распределении, температуре и солёности воды, а также типах грунтов в местах обитания шести видов ликодов: югорского Lycodes jugoricus, бледного L. pallidus, полярного L. polaris, сетчатого L. reticulatus, стрельчатого L. sagittarius и L. cf. palearis. Экземпляры, наиболее сходные с гребенчатым ликодом L. cf. palearis, впервые обнаружены в этом море – на мелководье о-ва Котельный (20–43 м). Обычным, относительно многочисленным и наиболее обширно распространённым видом в море Лаптевых является полярный ликод; частота встречаемости 24.7 против 1.2−6.2% остальных пяти видов. Приводятся размеры пойманных особей, соотношение длина–масса для югорского и полярного ликодов. Рассмотрен состав пищи югорского, полярного и сетчатого ликодов.
Исследования разнообразия, структуры и процессов функционирования рыбной части сообществ экосистем арктических морей имеет важное теоретическое и прикладное значение. Особенно актуальными такие исследования становятся в периоды климатических колебаний, когда в полной мере раскрывается истинный потенциал видов и проявляются их физиологические и продукционные возможности в постоянно меняющихся условиях среды. Несмотря на очевидную значимость, подобного рода исследования для высокоширотной Арктики практически отсутствуют. Имеющиеся в настоящее время данные не дают оснований утверждать, что условно негативные процессы, уже наблюдаемые в арктических морях, не приведут к изменению структурно-функциональных характеристик ихтиоценов региона. Это связано с тем, что до настоящего времени нет полных сведений по биологии непромысловых арктических видов, играющих важную роль в экосистемах полярных районов. Для большинства из них нет никаких данных о степени влияния тех или иных факторов на формирование их местообитаний, без чего невозможно прогнозирование экосистемных последствий от различного рода климатических процессов и техногенных воздействий. Целенаправленные исследования причинно-следственных отношений рыб к меняющимся факторам среды обитания являются обязательным условием получения данных о типологии ареалов, их динамике, общих закономерностях формирования рыбной части сообществ холодноводных экосистем.
Одним из элементов таких экосистем являются представители рода Lycodes, которые достаточно широко распространены в Арктике (Андрияшев, 1954; Mecklenburg et al., 2011, 2013). Встречаются ликоды и в море Лаптевых – одной из наиболее суровых по условиям среды обитания акваторий.
Цель настоящей работы – исследовать распределение, экологию и биологию шести видов ликодов, собранных в море Лаптевых в сентябре–октябре 2014 г.
МАТЕРИАЛ И МЕТОДИКА
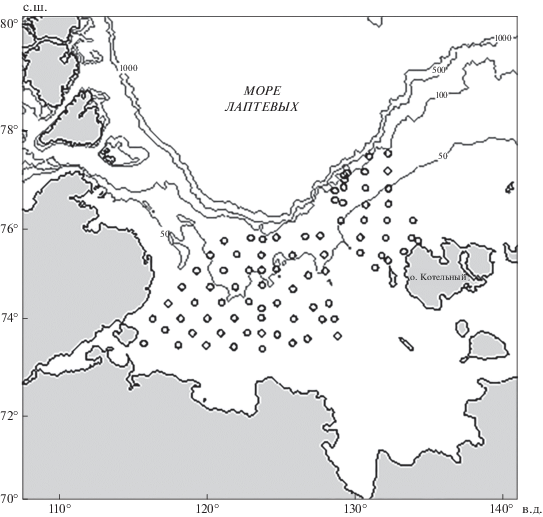
Материал собран в рейсе НИС “Дальние Зеленцы” на достаточно обширной акватории моря Лаптевых 01.09−05.10.2014 г. Траления выполнены на 81 комплексной станции от континентального склона на севере до почти прибрежных районов на юге (рис. 1). Лов проводили конвенционным донным тралом (чертёж 2837-00-000), оснащённым норвежским грунтропом-рокхоппером, который позволяет захватывать с поверхности дна даже самые мелкие объекты. Вертикальное раскрытие трала составляет 10 м, горизонтальное – 22 м, ячея 135 мм с кутовой мелкоячейной вставкой 12 мм, имеющей длину около 4 м. Длительность буксировки составляла 30 мин, скорость траления – 2.5 узла. На станциях с глубинами <20 м использовали трал Сигсби с входным периметром 100 × 30 см (0.3 м2), ячеёй 7 мм; продолжительность траления 10 мин.
Рис. 1.
Карта-схема траловых станций (⚪), выполненных в море Лаптевых в сентябре–октябре 2014 г.; (—) – изобаты.
Всего поймано 75 ликодов, относящихся к шести видам: L. jugoricus, L. pallidus, L. polaris, L. reticulatus, L. sagittarius и L. cf. palearis. У всех пойманных особей измеряли полную длину (TL) с точностью до 1 мм и массу с точностью до 0.1 г; представителей наиболее массовых видов – L. jugoricus и L. polaris – вскрывали для определения пола и качественного состава содержимого пищеварительных трактов. Интенсивность питания оценивали визуально по степени наполнения желудков по пятибалльной шкале (Методическое пособие .., 1974; Инструкции …, 2001).
Для характеристики среды обитания ликодов на каждой станции измеряли температуру и солёность воды с помощью автоматического СТД-зонда (SBE 19 plus V2 S/N19P73910-7324, “Sea-bird Electronics”, США) и отбирали пробы грунта дночерпателем Ван-Вина; тип грунта определяли по наиболее часто используемой их градации (Клёнова, 1960).
Средние значения численности и биомассы ликодов рассчитывали на основании количественных данных по составу уловов и площади облова (Аксютина, 1968), но без учёта коэффициента уловистости, который для данного типа трала неизвестен.
Латинские названия рыб даны в соответствии с аннотированным списком рыб арктического региона (Mecklenburg et al., 2013) и современной систематикой рыб (Eschmeyer et al., 2018); русские названия − по Черешневу и Кириллову (2007).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Видовой состав. Из шести видов ликодов, обнаруженных в наших уловах в 2014 г., пять встречались в море Лаптевых и ранее. Кроме того, в сборах отмечены экземпляры, которых по определителям (Андрияшев, 1954; Mecklenburg et al., 2018) следует отнести к гребенчатому ликоду L. palearis (рис. 2), так как от наиболее сходного L. pallidus они отличаются нечёрным перитонеумом. Однако северотихоокеанский L. palearis в море Лаптевых не был отмечен: его ареал доходит в Арктике лишь до восточной части Чукотского моря (Парин и др., 2014; Mecklenburg et al., 2018). Кроме того, наши экземпляры несколько отличаются от L. palearis менее выраженными лопастями подбородочных гребней. По этим причинам более обосновано обозначить их как L. cf. palearis. В то же время считаем важным включить их в рассмотрение, чтобы обратить внимание последующих исследователей на особенности условий обитания этой формы.

Согласно последнему аннотированному каталогу рыб семейства Zoarcidae род Lycodes включает 60 видов (Anderson, Fedorov, 2004), из которых 25 встречаются в арктическом регионе (Balushkin et al., 2011; Mecklenburg et al., 2013). В целом в северных морях России обитают 20 видов ликодов (Есипов, 1940; Андрияшев, 1954; Черешнев, Кириллов, 2007; Боркин и др., 2008; Карамушко, 2008; Balushkin et al., 2011; Wienerroither et al., 2011; Dolgov, 2013; Mecklenburg, Steinke, 2015), а их число от Баренцева до Чукотского морей варьирует от 3 видов в Белом море до 12 (с учётом последних данных) в море Лаптевых. Таким образом, фауна Lycodes в море Лаптевых наиболее разнообразна из всех северных морей России (табл. 1).
Таблица 1.
Видовой состав и встречаемость рыб рода Lycodes в северных морях России
| Вид | Море | |||||
|---|---|---|---|---|---|---|
| Баренцево | Белое | Карское | Лаптевых | Восточно-Сибирское | Чукотское | |
| L. adolfi Nielsen et Fosså, 1993 – ликод Адольфа | – | – | – | + | – | + |
| L. attenuatus Knipowitsch, 1906 – укороченный ликод | – | – | – | + | – | – |
| L. esmarkii Collett, 1875 – узорчатый ликод | + | – | + | – | – | – |
| L. eudipleurostictus Jensen, 1902 – двупёрый ликод | + | – | + | + | – | – |
| L. frigidus Collett, 1879 – абиссальный ликод | – | – | – | + | – | + |
| L. gracilis Sars, 1867 – тонкий ликод | + | – | – | – | – | – |
| L. jugoricus Knipowitsch, 1906 – югорский ликод | + | + | + | + | + | – |
| L. luetkenii Collett, 1880 – ликод Люткена | + | – | + | – | – | – |
| L. marisalbi Knipowitsch, 1906 – беломорский ликод | – | + | – | – | – | – |
| L. mucosus Richardson, 1855 – слизистый ликод | – | – | – | – | – | + |
| L. palearis Gilbert 1896 – гребенчатый ликод | – | – | – | +* | – | + |
| L. pallidus Collett, 1879 – бледный ликод | + | – | + | + | + | – |
| L. polaris (Sabine, 1824) – полярный ликод | + | + | + | + | + | + |
| L. raridens Taranetz et Andriashev, 1937 – редкозубый ликод | – | – | – | – | + | + |
| L. reticulatus Reinhardt, 1835 – сетчатый ликод | + | – | + | + | – | – |
| L. rossii Malmgren, 1865 – ликод Росса | + | – | + | + | + | – |
| L. sagittarius McAllister 1976 – стрельчатый ликод | – | – | + | + | – | – |
| L. seminudus Reinhardt, 1837 – полуголый ликод | + | – | + | + | – | + |
| L. squamiventer Jensen, 1904 – чешуебрюхий ликод | + | – | + | – | – | – |
| L. turneri Bean, 1879 – ликод Тернера | – | – | – | – | – | + |
| Всего | 11 | 3 | 11 | 12 | 5 | 8 |
Следует отметить, что статус встречающегося в море Лаптевых укороченного ликода L. attenuatus требует ревизии (Mecklenburg et al., 2016), но в настоящее время он рассматривается как валидный вид (Balushkin et al., 2011; Парин и др., 2014; Eschmeyer et al., 2018) и поэтому пока включён в общий список.
Пространственное распределение и частота встречаемости (рис. 3). Среди обнаруженных в море Лаптевых видов наиболее обширной акваторией распространения в 2014 г. характеризуется полярный ликод, который встречался в основном в южной и восточной частях района исследований (рис. 3г). Но даже у данного относительно многочисленного вида частота встречаемости в уловах составила 24.7% (табл. 2). Сведения о том, что полярный ликод встречается на шельфе моря Лаптевых и в эстуарии р. Хатанга, приводились и ранее, но наиболее широко данный вид распространён на юге и юго-западе (Андрияшев, 1948, 1954; Balushkin et al., 2011), хотя отдельные экземпляры встречались и на северо-востоке моря (Chernova, Neyelov, 1995). В настоящее время можно с полной уверенностью сказать, что на северо-востоке моря Лаптевых полярный ликод также не является редким.
Рис. 3.
Места поимок особей шести видов рода Lycodes в море Лаптевых: а – югорский ликод L. jugoricus, б − L. cf. palearis, в – бледный ликод L. pallidus, г – полярный ликод L. polaris, д – сетчатый ликод L. reticulatus, е – стрельчатый ликод L. sagittarius; (⚫) – наши данные, (▲) – данные литературы; ост. обозначения см. на рис. 1.
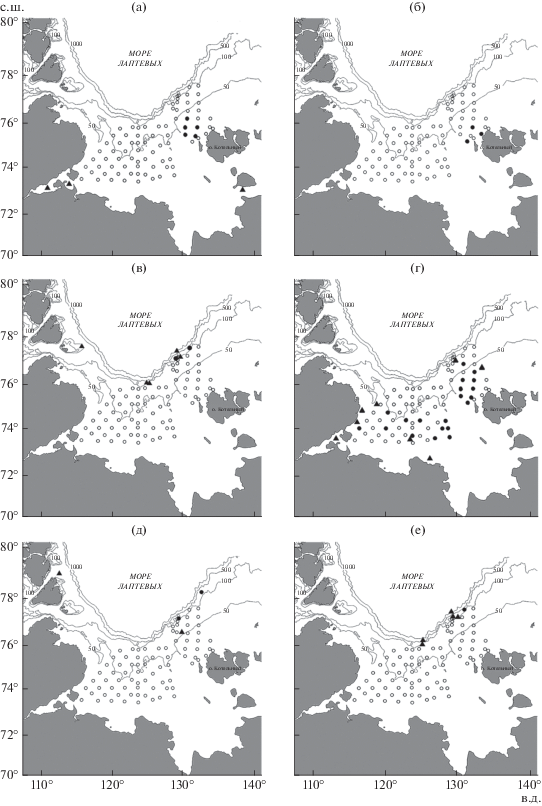
Таблица 2.
Частота встречаемости, плотность распределения и размеры особей шести видов рода Lycodes в море Лаптевых, сентябрь–октябрь 2014 г.
| Вид | Число рыб, экз. | Частота встречаемости, % |
Плотность | Длина (TL), мм | Масса, г | |
|---|---|---|---|---|---|---|
| экз/км2 | кг/км2 | |||||
| L. jugoricus | 11 | 6.2 | 47.5 | 12.25 | $\frac{{190{\kern 1pt} --{\kern 1pt} 460}}{{305.0 \pm 78.1}}$ | $\frac{{32.0{\kern 1pt} --{\kern 1pt} 840.0}}{{257.8 \pm 231.8}}$ |
| L. cf. palearis | 3 | 3.7 | 21.6 | 2.63 | 116, 145, 194 | 4.0, 17.0, 20.0 |
| L. pallidus | 2 | 2.5 | 21.6 | 1.56 | 111, 180 | 5.2, 22.0 |
| L. polaris | 54 | 24.7 | 58.3 | 10.63 | $\frac{{77{\kern 1pt} --{\kern 1pt} 285}}{{194.4 \pm 47.9}}$ | $\frac{{2.1{\kern 1pt} --{\kern 1pt} 123.0}}{{45.2 \pm 29.5}}$ |
| L. reticulatus | 4 | 2.5 | 32.4 | 1.99 | 98, 292, 298, 315 | 4.6, 182.0, 205.0, 265.0 |
| L. sagittarius | 1 | 1.2 | 21.6 | 1.08 | 246 | 50.0 |
Экземпляры L. cf. palearis были пойманы у западного побережья о-ва Котельный архипелага Новосибирские острова в точках с координатами: 75°53.3ʹ с.ш. 134°35.4′ в.д., 76°06.4ʹ с.ш. 137°19.5ʹ в.д. и 76°27.9ʹ с.ш. 135°59.1ʹ в.д. (рис. 3б). Югорский ликод, как и L. cf. palearis, обнаружен только в одном локальном районе к северо-западу от о-ва Котельный (рис. 3а), поэтому и частота встречаемости этих видов была низкой – соответственно 6.2 и 3.7% (табл. 2). Следует отметить, что югорский ликод также обнаружен гораздо севернее тех районов, которые известны по прежним поимкам (Андрияшев, 1948, 1954; Balushkin et al., 2011) и по ареалу вида в Арктике (Mecklenburg et al., 2018).
На континентальном склоне в северной части моря Лаптевых на приграничных станциях обследованного района были зарегистрированы единичные или редкие поимки трёх видов ликодов – бледного, сетчатого и стрельчатого (рис. 3в, 3д, 3е). Координаты нахождений: L. pallidus – 78°23.1ʹ с.ш. 136°11.1ʹ в.д. и 78°03.0ʹ с.ш. 132°56.3ʹ в.д., L. reticulatus – 78°04.3ʹ с.ш. 133°24.4ʹ в.д. и 79°02.8ʹ с.ш. 140°01.9ʹ в.д., L. sagittarius − 78°23.1ʹ с.ш. 136°11.1ʹ в.д. Частота их встречаемости составила 1.2–2.5% (табл. 2). Эти виды встречались в море Лаптевых и ранее, и их численность всегда была очень низкой (Chernova, Neyelov, 1995).
Батиметрическое распределение. Дно моря Лаптевых представляет собой слабо изрезанную равнину с неглубокими желобами, возвышенностями и банками (Добровольский, Залогин, 1982). Около половины площади моря составляют районы с глубинами <50 м, а в южной его части даже <25 м. На севере шельфовой зоны глубины плавно увеличиваются до 100 м, после чего начинается довольно крутой континентальный свал, заканчивающийся глубоководной равниной. Практически все эти глубины доступны для определённых видов ликодов, но в 2014 г. исследования проведены только на шельфе и самой верхней части континентального склона.
Основная доля особей полярного ликода (98.1%) обнаружена на глубине от 21 до 50 м, а общий диапазон глубин для данного вида составил 21–55 м. Эти значения достаточно близки к зарегистрированным ранее – 22–72 м (Андрияшев, 1939; Chernova, Neyelov, 1995), но теперь диапазон глубин, на которых встречался полярный ликод в море Лаптевых, находится в пределах 21–72 м.
Глубина нахождений L. cf. palearis в море Лаптевых – 20–43 м; границы батиметрического распространения гребенчатого ликода в ареале в целом существенно шире и составляют 2−925 м (Mecklenburg et al., 2018).
Югорский ликод в 2014 г. пойман на глубинах 21–37 м, что несколько глубже, чем указывалось ранее – 12–14 м (Андрияшев, 1954; Andriashev, 1986). Поэтому на данный момент батиметрический диапазон обитания югорского ликода в море Лаптевых находится в пределах 12–37 м. Сетчатый ликод в 2014 г. также отмечен заметно глубже – 240–277 м, чем в более ранний период – 54–125 м (Андрияшев, 1954; Chernova, Neyelov, 1995). Таким образом, общий диапазон глубин, на которых встречается сетчатый ликод в море Лаптевых, составляет теперь 54–277 м.
Бледный ликод в период наших исследований был обнаружен на глубинах от 255 до 307 м, но в море Лаптевых отмечался ранее в гораздо более широком диапазоне – 72–2151 м (Андрияшев, 1948; Chernova, Neyelov, 1995; Balushkin et al., 2011). То же касается и стрельчатого ликода, который в 2014 г. был обнаружен на глубине 277 м, хотя в предшествующие годы был отмечен на глубинах 120−2151 м (Chernova, Neyelov, 1995).
Термический диапазон обитания. Температура водных масс в море Лаптевых даже в летние месяцы в его южной части на горизонте 25 м и глубже составляет от – 1.4 до –1.5°С (Добровольский, Залогин, 1982). В северных районах моря на континентальном склоне отрицательные значения температуры прослеживаются приблизительно до глубины 100 м и потом после 300 м, а промежуточный слой из-за влияния тёплых атлантических вод характеризуется повышенным теплосодержанием (0.6–0.8°С). Все представители рода Lycodes, встречающиеся в море Лаптевых, по характеру географического ареала относятся к арктическим видам (Андрияшев, 1954; Парин и др., 2014; Mecklenburg et al., 2018), что предполагает возможность их существования при экстремально низких отрицательных значениях температуры.
По результатам исследований, выполненных в море Лаптевых в 2014 г., в наиболее широком диапазоне температуры обитают виды, которые встречались на мелководье (L. jugoricus, L. cf. palearis, L. polaris), где сезонные колебания температуры, как правило, максимальны. Для этих видов диапазоны температуры составили соответственно 3.17, 4.45 и 4.22°С. При этом доля особей, встречавшихся при самых низких значениях температуры, была существенной − 33.3–47.2% (табл. 3). Для L. cf. palearis предшествующих данных по термическому диапазону обитания в море Лаптевых не было; для гребенчатого ликода в пределах ареала он составляет от –2.1 до 14.6°С (Mecklenburg еt al., 2016). При наиболее низких отрицательных значениях отмечен и бледный ликод, но фактическая температура воды при его нахождении (–1.04°С) оказалась заметно выше, чем у видов, пойманных на мелководье. В целом бледный ликод в море Лаптевых отмечен при температурах от –1.80 (Андрияшев, 1948) до 0.49°С (Chernova, Neyelov, 1995). Сетчатый ликод в 2014 г. обнаружен только при положительных значениях температуры – 0.4−1.1°С, хотя ранее данный вид был пойман в море Лаптевых при температуре –1.6°С (Chernova, Neyelov, 1995). Стрельчатый ликод обнаружен при температуре 1.12°С. В предшествующий период этот вид встречался в море Лаптевых как при отрицательных (–0.76°), так и положительных (0.68°) значениях температуры (Chernova, Neyelov, 1995).
Таблица 3.
Распределение особей шести видов рода Lycodes при разной температуре и температурный диапазон их встречаемости в море Лаптевых, сентябрь–октябрь 2014 г., %
| Вид | Температура, °С | Диапазон температуры, °С | ||||
|---|---|---|---|---|---|---|
| –2.00 … 1.01 | –1.0 … 0 | 0.01–1.00 | 1.01–2.00 | 2.01–3.00 | ||
| L. jugoricus | 45.5 | – | – | 54.5 | – | –1.50 ... 1.67 |
| L. cf. palearis | 33.3 | – | – | 33.3 | 33.3 | –1.72 ... 2.73 |
| L. pallidus | 100 | – | – | – | – | –1.04 |
| L. polaris | 47.2 | 30.6 | – | 16.7 | 5.6 | –1.72 ... 2.5 |
| L. reticulatus | – | – | 66.7 | 33.3 | – | 0.40−1.10 |
| L. sagittarius | – | – | – | 100 | – | 1.12 |
Солёность воды в местах обитания. Пространственно-временное распределение солёности воды в море Лаптевых крайне неоднородно, но её увеличение закономерно происходит с юго-востока на северо-запад и север (Добровольский, Залогин, 1982). Важную роль в формировании солёностного режима моря Лаптевых играет приток континентальных пресных вод, ежегодный объём которого составляет ~ 541 км3 (Bauch et al., 2009), а подавляющая его часть поступает в восточную часть моря. На мелководных южных и юго-восточных участках солёность варьирует в пределах 1–5‰, на северо-западе увеличивается до 34‰, а в среднем колеблется в пределах 20–30‰ (Добровольский, Залогин, 1982). Эти характерные особенности моря Лаптевых могут оказывать влияние на пространственное распределение разных видов ликодов.
В 2014 г. наиболее эвригалинные виды были выловлены на мелководье. Так, в местах поимки югорского ликода солёность вод варьировала в пределах 25.6–33.8‰, L. cf. palearis – 25.25–33.85‰, полярного – 28.55–34.75‰ (табл. 4). Югорский ликод ранее встречался только в прибрежье моря Лаптевых и в более опреснённых водах – 15–23‰ (Андрияшев, 1948, 1954). Полярный ликод и в предшествующие годы был зарегистрирован в районах с солёностью 25.3–34.86‰ (Андрияшев, 1948; Chernova, Neyelov, 1995).
Таблица 4.
Распределение особей шести видов рода Lycodes при разной солёности придонных вод и солёностный диапазон их встречаемости в море Лаптевых, сентябрь–октябрь 2014 г., %
| Вид | Солёность, ‰ | Диапазон солёности, ‰ | ||||
|---|---|---|---|---|---|---|
| 25.00–26.99 | 27.00–28.99 | 29.00–30.99 | 31.00–32.99 | 33.00–34.99 | ||
| L. jugoricus | 9.1 | 9.1 | 36.3 | 9.1 | 36.4 | 25.60–33.80 |
| L. cf. palearis | 33.3 | – | 33.3 | – | 33.4 | 25.25–33.85 |
| L. pallidus | – | – | – | – | 100 | 34.00–34.53 |
| L. polaris | – | 5.6 | 5.6 | 36.1 | 52.8 | 28.55–34.75 |
| L. reticulatus | – | – | – | – | 100 | 34.68–34.80 |
| L. sagittarius | – | – | – | – | 100 | 34.68 |
Бледный, сетчатый и стрельчатый ликоды встречались только в водах с океанической солёностью – соответственно 34.0–34.53, 34.68–34.80 и 34.68‰ (табл. 4). Ранее среди этих видов только бледный ликод был обнаружен при более низком значении солёности – 30.80‰ (Андрияшев, 1948), а L. reticulatus и L. sagittarius отмечены соответственно при 33.03–34.24 и 34.66–34.82‰ (Chernova, Neyelov, 1995).
Следует отметить, что в летний период 1941 г. в б. Кожевникова, которая расположена в средней части Хатангского залива, были пойманы два ликода, видовая принадлежность которых не была определена (Михин, 1941), но данное обстоятельство интересно тем, что солёность воды в этом районе колебалась в пределах 3.89–11.98‰, что существенно меньше 15‰ − минимального значения, зарегистрированного ранее для эвригалинных ликодов в море Лаптевых.
Распределение по типу донных грунтов. Состав грунтов в море Лаптевых, как и в целом в арктических морях, зависит от предшествующих и современных условий их формирования. В более глубоководных районах дно покрыто илами, а на мелководьях донные осадки состоят из ила и песка с включениями гальки и валунов (Суховей, 1986).
Анализ сведений о характере донных осадков в местах вылова ликодов в 2014 г. показал, что они обитают на грунтах четырёх типов (рис. 4). Так, в местах нахождения L. jugoricus, L. polaris и L. cf. palearis прослеживается от двух до четырёх типов грунтов, основными из которых являются илистый песок с серой глиной и серо-коричневые песчаные илы. Встречаемость данных видов на этих грунтах составляла 67–100%. Часть особей была обнаружена также в районах с коричневым илом, включающим коричнево-серую глину и песчаный ил с галькой. Местонахождение ликодов, выловленных на верхнем участке свала глубин (L. pallidus, L. reticulatus, L. sagittarius), характеризуется наличием всего одного типа грунта – коричневого ила, включающего коричнево-серую глину. В данном случае следует учитывать небольшое число пойманных особей, что и отразилось на величине встречаемости (100%).
Рис. 4.
Встречаемость особей шести видов рода Lycodes на грунтах разных типов в море Лаптевых, сентябрь−октябрь 2014 г.: ( ) – илистый песок и серая глина; (
) – илистый песок и серая глина; ( ) – серые или коричневые песчанистые илы, серая глина; (
) – серые или коричневые песчанистые илы, серая глина; ( ) – коричневый ил, коричневая или серая глина; (◼) – песчанистый ил c галькой.
) – коричневый ил, коричневая или серая глина; (◼) – песчанистый ил c галькой.
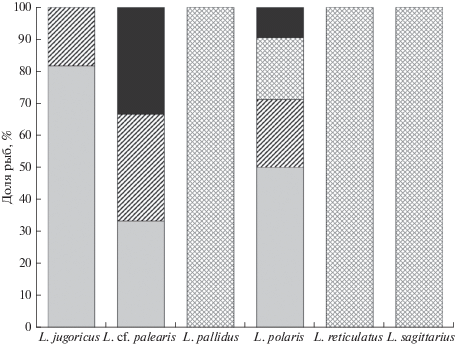
В более ранних исследованиях также отмечалось, что, например, бледный ликод обитает на участках с илистыми грунтами, но без детального уточнения их состава (Андрияшев, 1954). Подобные описания встречаются и для других видов, когда приводятся сведения только для основных фракций грунта – песка или ила (Андрияшев, 1948, 1954; Balushkin et al., 2011), что затрудняет анализ предшествующих данных и полученных в 2014 г.
Численность и биомасса. Наиболее плотные скопления по численности отмечены у полярного ликода – 58.3 экз/км2, а по биомассе у югорского – 12.25 кг/км2 (табл. 2). Относительная численность и биомасса разных видов ликодов в море Лаптевых кажутся низкими. Но полученные величины существенно выше опубликованных ранее данных: например, биомасса югорского ликода в прибрежье Новосибирских о-вов оценивалась на уровне 1.2 кг/км2 (Чернова, 2015), а пяти представителей семейства Zoarcidae, четырёх видов ликодов (L. eudipleurostictus, L. pallidus, L. polaris, L. reticulatus) и гимнелюса Книповича Gymnelus knipowitschi составляла 5.33 кг/км2 (Глебов и др., 2016). Следует отметить, что относительная биомасса только трёх из четырёх упомянутых выше ликодов (L. pallidus, L. polaris, L.reticulatus), которые также встречались в наших уловах, была существенно больше – 14.18 кг/км2.
Некоторые черты биологии. Длина исследованных особей даёт только предварительное представление о размерном составе рыб каждого вида в море Лаптевых, поскольку выборки, за исключением югорского и полярного ликодов, ограничены.
Длина югорского ликода варьировала в пределах 190–460 мм (табл. 2), причём наибольшая величина достаточно близка к зарегистрированному максимальному размеру данного вида – 510 мм (Mecklenburg et al., 2016). Наличие в уловах крупных особей отразилось на средних показателях длины и массы, которые составили соответственно 305.0 мм и 257.8 г, хотя половина рыб была заметно мельче (225–285 мм, 68–156 г). До настоящего времени длина югорского ликода в море Лаптевых была известна только по 3 экз. – 61, 155 и 260 мм (Андрияшев, 1948, 1954). Аппроксимация степенной функцией соотношения длина–масса особей югорского ликода в нашей выборке указывает на наличие положительной аллометрии роста (рис. 5а).
Рис. 5.
Соотношение между длиной (TL) и массой тела двух видов рода Lycodes в море Лаптевых: а − югорский ликод L. jugoricus, у = 0.0007х3.66, R2 = 0.9918; б − полярный ликод L. polaris, у = 0.0045х3.05, R2 = 0.9508.
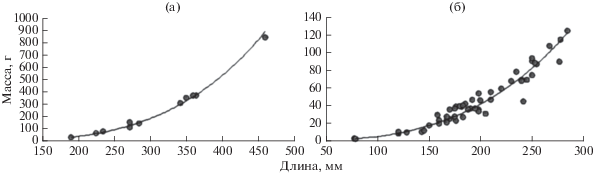
Длина полярного ликода составляла от 77 до 285 мм и, так же, как у югорского ликода, наибольший размер особей был достаточно близок к максимальной зарегистрированной длине данного вида – 333 мм (Mecklenburg et al., 2016). Средние длина и масса составили соответственно 194.4 мм и 45.2 г (табл. 2). Соотношение длины и массы полярного ликода показывает, что у данного вида в море Лаптевых наблюдается практически изометрический рост (рис. 5б).
Соотношение самцов и самок в выборке полярного ликода было почти равным – 1.0 : 1.1. Две самки TL 176.0 и 200.0 мм имели практически созревшие и, по-видимому, готовые к вымету икринки диаметром 5.0–5.5 мм; в их гонадах присутствовали также икринки меньшего размера, что может указывать на порционный нерест. У нескольких других самок диаметр икры был немного меньше (3.5–4.0 мм). Наши данные указывают на позднеосенний нерест этого вида, что предполагалось и ранее (Андрияшев, 1954).
Присутствие в наших выборках рыб разного размера, а также половозрелых особей полярного ликода указывает на наличие процесса воспроизводства и пополнения популяций ликодов в данном регионе. Что касается гребенчатого ликода, то его биология практически не изучена, но, учитывая максимальную длину этого вида (TL 62 см) (Mecklenburg et al., 2016) и размеры молодых экземпляров L. cf. palearis, пойманных в 2014 г. (TL 11.6–19.4 см), можно предположить, что на мелководье о-ва Котельный могут находиться выростные участки этой формы.
Сведений о питании ликодов в море Лаптевых, за исключением данных для 1 экз. L. jugoricus (Андрияшев, 1954), практически нет. Наши данные, характеризующие питание югорского, полярного и сетчатого ликодов, представлены в табл. 5.
Таблица 5.
Состав пищевых объектов и частота их встречаемости у особей трёх видов рода Lycodes в море Лаптевых, сентябрь–октябрь 2014 г. , %
| Компонент пищи и другие показатели | Вид и размерная группа (TL), мм | ||||
|---|---|---|---|---|---|
| L. jugoricus | L. polaris | L. reticulatus | |||
| 360–364 | 460 | 160–192 | 205–285 | 315 | |
| Рыба | 100 | – | 50.0 | 42.8 | 100 |
| Isopoda, род Saduria | 100 | 100 | 25.0 | – | – |
| Hyperiidea | – | – | – | 14.3 | – |
| Amphipoda | – | – | – | 42.8 | – |
| Cumacea, сем. Diastylidae | – | – | 25.0 | – | – |
| Bivalvia, род Yoldia | 50.0 | – | – | – | – |
| Holothuroidea, род Myriotrochus | – | – | – | 14.3 | – |
| Число рыб, экз. | 2 | 1 | 7 | 7 | 1 |
| Доля питавшихся особей, % | 100 | 100 | 57 | 100 | 100 |
| Средний балл наполнения желудков | 3 | 1 | 0.6 | 2 | 2 |
В желудках югорского ликода обнаружены равноногие раки рода Saduria, двустворчатые моллюски рода Yoldia и переваренная рыба; в желудке самой крупной особи – изоподы. Однако из-за ограниченного числа просмотренных рыб делать какие-либо выводы о различиях спектра питания рыб разного размера пока преждевременно. В отличие от исследованных нами особей желудок единственного проанализированного экземпляра, пойманного в 1913 г. у входа в б. Нарвик, прилегающую к Хатангскому заливу, содержал много прибрежных Amphipoda и остатки Polychaeta (Андрияшев, 1954).
В составе пищи полярного ликода отмечены шесть объектов, что заметно разнообразнее, чем у L. jugoricus. В желудках обнаружены изоподы рода Saduria, Hyperiidae, Amphipoda, кумовые раки семейства Diastylidae, голотурия рода Myriotrochus и гомогенные остатки рыб, среди которых был идентифицирован достаточно сохранившийся ильный люмпен Anisarchus medius. Состав пищи у особей разного размера незначительно различался как по количеству объектов, так и по таксономическому составу ракообразных. Информацию о питании полярного ликода в море Лаптевых в доступных источниках литературы мы не обнаружили. В сопредельном Карском и далее в Баренцевом морях этот вид характеризуется как типичный бентофаг, в питании которого доминируют полихеты, креветки и амфиподы (Брискина, 1939; Долгов, 2014; Долгов и др., 2014).
В желудке единственного просмотренного экземпляра сетчатого ликода обнаружена только полупереваренная рыба. По-видимому, рыба является основным источником энергии для взрослых особей сетчатого ликода, поскольку, например, при исследовании питания непромысловых видов рыб в северной части Баренцева моря было установлено, что в пище сетчатого ликода доминируют Boreogadus saida, Mallotus villosus, представители семейств Liparidae, Cottidae, Stichaeidae, молодь Hippoglossoides platessoides и Lycodes sp. (Долгов, 2014). В Карском море в качестве основной пищи сетчатого ликода зарегистрированы B. saida и непромысловые виды рыб – Liparis fabricii, представители Cottidae и молодь Lycodes sp. (Долгов и др., 2014).
ВЫВОДЫ
1. В сентябре–октябре 2014г. в море Лаптевых зарегистрировано шесть видов рыб рода Lycodes; из которых наиболее многочисленным является полярный ликод: частота встречаемости 24.7 против 1.2−6.2% остальных пяти видов.
2. Батиметрическое распределение, температура и солёность воды в местах поимок ликодов в общем сходны с наблюдаемыми в других частях их ареалов. Уточнены диапазоны этих параметров среды для моря Лаптевых: батиметрический диапазон обитания югорского (12–37 м) и сетчатого ликодов (54–277 м), а также температура, при которой встречались стрельчатый (–0.76 … 1.12°С) и сетчатый (–1.6 … 1.1°С) ликоды. Югорский ликод отмечен в водах с большей солёностью (33.8‰), а бледный ликод – при более низкой (30.80‰), чем было известно ранее, L. cf. palearis обитает на глубине 20–43 м при температуре –1.72 … 2.73°С и солёности воды 25.25−33.85‰.
3. Получены первые данные по питанию полярного и сетчатого ликодов в море Лаптевых. Спектр питания полярного ликода включает Isopoda, Hyperiidae, Amphipoda, Cumacea и рыбу; сетчатый ликод питается только рыбой. Дополнены сведения по составу пищи югорского ликода: в 2014 г. он потреблял Isopoda, Bivalvia и рыбу.
Список литературы
Аксютина З.М. 1968. Элементы математической оценки результатов наблюдений в биологических и рыбохозяйственных исследованиях. М.: Пищ. пром-сть, 289 с.
Андрияшев А.П. 1939. Новые данные по экологии и распространению рыб моря Лаптевых // ДАН СССР. Т. 23. № 7. С. 728–731.
Андрияшев А.П. 1948. К познанию рыб моря Лаптевых // Тр. ЗИН АН СССР. Т. VII. № 3. С. 76–100.
Андрияшев А.П. 1954. Рыбы северных морей СССР. М.; Л.: Изд-во АН СССР, 556 с.
Боркин И.В., Васильев А.В., Четыркина О.Ю. 2008. Ихтиофауна // Экосистема Карского моря. Мурманск: Изд-во ПИНРО. С. 130–206.
Брискина М.М. 1939. Питание непромысловых рыб Баренцева моря // Тр. ВНИРО. Т. IV. С. 339−354.
Глебов И.И., Надточий В.А., Савин А.Б. и др. 2016. Результаты комплексных биологических исследований в море Лаптевых в августе−сентябре 2015 г. // Изв. ТИНРО. Т. 187. С. 72−88.
Добровольский А.Д., Залогин Б.С. 1982. Моря СССР. М.: Изд-во МГУ, 192 с.
Долгов А.В. 2014. Питание непромысловых рыб в северной части Баренцева моря // Особенности формирования биопродуктивности северных районов Баренцева моря в период потепления Арктики. Мурманск: Изд-во ПИНРО. С. 155−185.
Долгов А.В., Бензик А.Н., Четыркина О.Ю. 2014. Питание непромысловых рыб и их роль в продуктивности экосистемы Карского моря // Тр. ВНИРО. Т. 152. С. 190–208.
Есипов В.К. 1940. Ихтиофауна моря братьев Лаптевых // Зоол. журн. Т. 19. Вып. 1. С. 139–142.
Инструкции и методические рекомендации по сбору и обработке биологической информации в районах исследований ПИНРО. 2001. Мурманск: Изд-во ПИНРО, 291 с.
Карамушко О.В. 2008. Видовой состав и структура ихтиофауны Баренцева моря // Вопр. ихтиологиии. Т. 48. № 3. С. 293–308.
Клёнова М. В. 1960. Геология Баренцова моря. М.: Изд-во АН СССР, 366 с.
Методическое пособие по изучению питания и пищевых отношений рыб в естественных условиях. 1974. М.: Наука, 254 с.
Михин В.С. 1941. Рыбы и рыбный промысел реки Хатанги и Хатангского залива // Тр. НИИ поляр. земледелия, животноводства и промысл. хоз-ва. Вып. 16. С. 37–72.
Парин Н.В., Евсеенко С.А., Васильева Е.Д. 2014. Рыбы морей России: аннотированный каталог. М.: Т-во науч. изд. КМК, 733 с.
Суховей В.Ф. 1986. Моря мирового океана. Л.: Гидрометеоиздат, 288 с.
Черешнев И.А., Кириллов А.Ф. 2007. Рыбообразные и рыбы морских и пресных вод бассейнов морей Лаптевых и Восточно-Сибирского // Вест. СВНЦ ДВО РАН. № 2. С. 95–106.
Чернова Н.В. 2015. Ихтиофауна морских вод Новосибирских островов (охранная зона заповедника “Усть-Ленский” // Науч. тр. гос. природ. заповедника “Присурский”. Т. 30. Вып. 1. С. 271–276.
Anderson M.E., Fedorov V.V. 2004. Family Zoarcidae Swainson 1839 – eelpouts // Calif. Acad. Sci. Annot. Checklists of Fish. № 34. P. 1–58.
Andriashev A.P. 1986. Zoarcidae // Fishes of the North-eastern Atlantic and the Mediterranean / Eds. Whitehead P.J.P. et al. Paris: UNESCO. P. 1130–1150.
Balushkin A.V., Sheiko B.A., Fedorov V.V. 2011. Catalog of the archival collection of the Zoological Institute, Russian Academy of Sciences: class Osteichthyes (bony fishes), order Perciformes, family Zoarcidae // J. Ichthyol. V. 51. № 10. P. 950–1034. https://doi.org/10.1134/S0032945211100031
Bauch D., Dmitrenko I.A., Wegner C. et al. 2009. Exchange of Laptev Sea and Arctic Ocean halocline waters in response to atmospheric forcing // J. Geophys. Res. V. 114. P. CO5008.
Chernova N.V., Neyelov A.V. 1995. Fish caught in the Laptev Sea during the cruise of RV Polarstern in 1993 // Ber. Polarforsch. № 176. P. 222–227.
Dolgov A.V. 2013. Annotated list of fish-like vertebrates and fish of the Kara Sea // J. Ichthyol. V. 53. № 11. P. 914–922. https://doi.org/10.1134/S0032945213110039
Eschmeyer W.N., Fricke R., Van der Laan R. 2018. Catalog of fishes (http://researcharchive.calacademy.org/research/ ichthyology/catalog/fishcatmain.asp. Version 01/2018).
Mecklenburg C.W., Steinke D. 2015. Ichthyofaunal baselines in the Pacific Arctic region and RUSALCA study area // Oceanography. V. 28. № 3. P. 158–189.
Mecklenburg C.W., Møller P.R., Steinke D. 2011. Biodiversity of Arctic marine fishes: taxonomy and zoogeography // Mar. Biodiv. V. 41. № 1. P. 109–140. https://doi.org/10.1007/s12526-010-0070-z
Mecklenburg C.W., Byrkjedal I., Christiansen J.S. et al. 2013. List of marine species of the Arctic region annotated with common names and zoogeographic characterizations. Akureyri, Iceland: CAFF, 35 p.
Mecklenburg C.W., Mecklenburg T.A., Sheiko B.A., Steinke D. 2016. Pacific Arctic marine fishes. Akureyri, Iceland: CAFF, 398 p.
Mecklenburg C.W., Lynghammar A., Johannesen E. et al. 2018. Marine fishes of the Arctic region. V. I. Akureyri, Iceland: CAFF, 342 p.
Wienerroither R., Johannesen E., Dolgov A. et al. 2011. Atlas of the Barents Sea fishes // IMR−PINRO Joint Report Series 1−2011. 273 p.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии