

Вопросы ихтиологии, 2019, T. 59, № 5, стр. 567-577
Некоторые результаты изучения динамики численности горбуши Oncorhynchus gorbuscha и кеты O. keta на северо-западном побережье острова Сахалин
1 Сахалинский научно-исследовательский институт рыбного хозяйства и океанографии − СахНИРО
Южно-Сахалинск, Россия
2 Сахалинский государственный университет − СахГУ
Южно-Сахалинск, Росссия
* E-mail: kaev@outlook.com
Поступила в редакцию 04.09.2018
После доработки 26.12.2018
Принята к публикации 16.01.2019
Аннотация
Проанализированы данные, характеризующие численность отдельных поколений горбуши Oncorhynchus gorbuscha и кеты O. keta на северо-западном побережье Сахалина (объём вылова, захода производителей в реки и последующего ската молоди). По динамике уловов и биологическим показателям рыб показано, что промысел базируется на подходах разных группировок этих видов рыб. Если для кеты наличие группировок связано с ходом рыб разных рас (летняя и осенняя) и разного происхождения (из Амура и небольших рек северо-западного побережья Сахалина и сопредельного материкового побережья), то для горбуши – в основном за счёт подхода рыб местного происхождения, но разных темпоральных форм.
Основу промысла тихоокеанских лососей рода Oncorhynchus в Сахалино-Курильском регионе составляет горбуша O. gorbuscha – около 80% уловов всех лососей в регионе. По результатам экологических и морфологических исследований было показано наличие в регионе локальных стад этого вида (Воловик, 1967а; Иванков, 1967). Это группировки рыб, воспроизводящихся в реках юго-западного, северо-западного, северо-восточного и юго-восточного побережья Сахалина, побережья заливов Терпения и Анива, островов Итуруп и Кунашир (рис. 1). Результаты последующих исследований позволили сформулировать положение о локальном стаде для лососей с коротким пресноводным периодом жизни, в соответствии с которым горбуша, размножающаяся в реках того или иного гидрогеологического массива, образует единую популяционную систему (локальное стадо), состоящую из группировок (популяций) рыб ряда рек. Эти группировки сходны между собой по основным биологическим характеристикам и типу динамики стада и отличаются от таких группировок из других районов (Иванков, 1993, 2011). Наличие между этими районами на Сахалине выступающих мысов, на которых нет рек для нереста, или широких проливов между островами способствует географической изоляции данных стад (Гриценко, 1990). В этих же районах воспроизводятся хорошо различающиеся между собой по морфобиологическим показателям группировки кеты O. keta (Иванков, 1970, 1972; Иванкова и др., 2000), однако её нерестилища в отличие от горбуши сосредоточены в основном лишь в бассейнах некоторых рек в местах массированного выхода грунтовых вод и, как следствие, наблюдается чёткая локализация её популяций (Каев, 2003; Макоедов и др., 2009).
Рис. 1.
Районы воспроизводства локальных стад горбуши Oncorhynchus gorbuscha и кеты O. keta на Сахалине и южных Курильских островах.
В текущем столетии на юго-западном, северо-западном, северо-восточном и юго-восточном побережьях Сахалина ежегодно вылавливают в среднем соответственно 954, 2258, 17 056 и 29 108 т горбуши, в заливах Терпения и Анива – 19 622 и 17 623 т, на островах Итуруп и Кунашир – 22 421 и 1699 т. В северных районах Сахалина промысловый запас горбуши формируется почти полностью за счёт естественного воспроизводства, тогда как в южных районах определённую роль играет её заводское разведение. Так, на Итурупе соотношение скатившейся с нерестилищ молоди и выпущенной с лососёвых рыборазводных заводов составляет 63 : 37, в зал. Анива − 59 : 41, на юго-восточном побережье Сахалина − 71 : 29, в зал. Терпения − 87 : 13. Промысловый запас кеты в регионе формируется в основном с помощью заводского разведения (Каев, Игнатьев, 2015); его пополнение за счёт естественного воспроизводства сохранилось в регионе только на Курильских о-вах, в зал. Терпения и на северо-западном побережье Сахалина.
Если для оценки состояния запасов заводских популяций горбуши и кеты используются данные по объёму выпуска молоди, то в отношении диких популяций проводятся специальные учёты производителей этих видов на нерестилищах ряда рек и молоди, скатывающейся с нерестилищ в контрольных реках. В связи с низким уровнем естественного воспроизводства кеты мониторинг ориентирован преимущественно на оценку состояния воспроизводства горбуши. При этом плотность наблюдений (доля ежегодно обследуемых (заход производителей) и контрольных (учёт покатной молоди) рек по отношению к общему числу рек) существенно выше в районах с наибольшими уловами на единицу протяжённости побережья. Именно для этих районов (охотоморское побережье Итурупа, юго-восточное побережье Сахалина и зал. Анива) в начале XXI в. были созданы базы данных по горбуше на основе унификации расчётов численности заходов производителей в реки и последующего ската молоди для всех лет наблюдений, а также биологических показателей рыб для характеристики поколений, на основе которых были изучены особенности воспроизводства горбуши этих стад (Kaev et al., 2007). В первой половине 1990-х гг. по стандартной схеме начался ежегодный сбор материалов на Кунашире, что также позволило обобщить многолетние данные по воспроизводству местных стад горбуши и кеты (Каев, Ромасенко, 2017). Воспроизводство лососей в других районах изучено в меньшей степени из-за слабой обеспеченности исходными данными. Для рационального использования запасов необходимы знания об особенностях динамики каждого из стад. К таким районам, несмотря на многолетнюю историю промысла, относится и северо-западное побережье Сахалина.
Цель исследования − установить особенности динамики стад горбуши и кеты в данном районе на основе обобщения имеющихся материалов по единой для всех лет наблюдений схеме.
МАТЕРИАЛ И МЕТОДИКА
При выборе исходных данных по численности лососей приходится учитывать их существенные расхождения в разных источниках. Так, например, по кете величина уловов при сравнении данных отечественного издания (Уловы …, 1989) и Северотихоокеанской комиссии по анадромным рыбам (Irvine et al., 2012) различается в некоторые годы до 1.4 раз. Сотрудники Охинской контрольно-наблюдательной станции (КНС) определяли численность производителей горбуши и кеты на нерестилищах визуальным подсчётом рыб при пешем обходе отдельных участков некоторых рек, а также с 1991 г. вели учёт покатной молоди этих видов в р. Лангры методом выборочных обловов (Воловик, 1967б). Регулярные исследования состояния запасов лососей в этом районе не проводили, а исходя из соотношения численности производителей в реках и последующего ската молоди не всегда понятно, как эти данные рассчитывали. В этой связи в качестве исходного материала для анализа использованы данные по вылову горбуши и кеты в районе (промысловая статистика в настоящее время представляется Сахалинским территориальным управлением Росрыболовства), заходам производителей этих видов в Лангры и скату молоди с нерестилищ этой реки (данные ихтиологической службы бассейнового управления Сахалинрыбвод), которые передаются для подготовки ежегодных статистических документов Северотихоокеанской комиссии по анадромным рыбам (NPAFC, 2018). До 2009 г. величину заходов горбуши и кеты во все реки района принимали по данным NPAFC, начиная с 2010 г. для расчёта суммарной величины заходов рыб в реки района использован принцип реперных рек, в соответствии с которым все реки подразделены на три группы. Первую группу составляют регулярно обследуемые реки, вторую – эпизодически обследуемые, третью – реки, в которых обследования не проводили. Рассчитанная плотность скоплений рыб (отношение подсчитанного числа рыб в реке к площади нерестилищ) в реках первой группы принималась за действительную. Для рек второй группы плотность скоплений рыб рассчитана как среднее значение между наблюдённой в данной реке и определённой для рек первой группы. Суммарные результаты наблюдений в реках первой и второй группы экстраполировали на реки третьей группы (Каев и др., 2004; Kaev, Klovach, 2014).
Для биологической характеристики рыб (длина по Смитту (FL), абсолютная индивидуальная плодовитость (АИП), коэффициент упитанности по Фультону (Кф)) использованы фондовые материалы СахНИРО, собранные при полевых наблюдениях в данном районе, объём проб составлял, как правило, 100 рыб. В тексте использованы следующие символы: M – среднее значение, SD – среднее квадратичное отклонение, r – коэффициент корреляции Пирсона, p – уровень значимости нуль-гипотезы, n – число поколений. Достоверность различия между средними значениями оценена по критерию Фишера (F).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
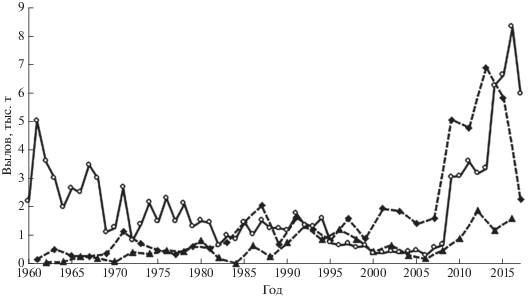
Северо-западный Сахалин является традиционным районом промысла горбуши и кеты, причём уловы кеты чаще были выше, чем горбуши (рис. 2). Статистика уловов горбуши, видимо, не в полной мере отражает межгодовые изменения её численности в силу того, что до середины 1990-х гг., когда начал набирать силу коммерческий промысел, предпочтение отдавалось лову кеты. Однако официальная статистика вылова кеты также не отражает реальное состояние её запасов, так как с начала 1980-х по середину 2000-х гг. вводились ограничения на её промысел, исходя из представлений об амурском происхождении лососей, облавливаемых вдоль сахалинского побережья Амурского лимана. Ведь только в бассейне Амура в 1950-е гг. насчитывалось 16 млн м2 нерестилищ преимущественно горбуши, летней и осенней кеты, хотя к концу века их площадь сократилось предположительно вдвое (Рослый, 2002).
Рис. 2.
Динамика уловов горбуши Oncorhynchus gorbuscha поколений нечётных (- -◆- -) и чётных (- -▲- -) лет и кеты O. keta (—⚪—) на северо-западном побережье Сахалина в 1960–2017 гг.
В реках северо-западного побережья Сахалина по современным оценкам имеется 1244 тыс. м2 нерестилищ горбуши (Каев, 2011) и 256 тыс. м2 – кеты (Каев, 2003, с уточнением). Всего от м. Погиби до м. Елизаветы в районе насчитывается 39 рек (не считая мелких водотоков и безымянных ручьёв), в которые отмечены заходы лососей, однако примерно для трети из них нет данных о площади нерестилищ. Нерестилища горбуши равномернее рассредоточены по разным рекам в соответствии с их размерами, в то время как у кеты их основная часть (66%) сосредоточена в р. Лангры. Нерестилища кеты в крупных реках расположены преимущественно в верхнем и в меньшей степени в среднем течении основного русла и впадающих притоков. Большое значение для нереста кеты принадлежит многочисленным ключевым ручьям и чашам, которые образуют порой целые системы небольших озёр, соединённых протоками. По наблюдениям в августе 1990 г., температура воды в таких ключевых ручьях составляла 8–9 против 16–18°С в речном потоке. В марте 1991 г. (зимняя межень) температура воды в районе выхода ключей колебалась в пределах 3−7°С (Каев, 2003).
В 2010–2017 гг. численность горбуши ежегодно учитывали только в р. Лангры, почти регулярно (исключая один−два года) – в реках Пильво и Большая, ещё в 13 реках наблюдения были эпизодическими. Ежегодные учёты кеты проводили в реках Лангры, Большая, Чингай и эпизодически ещё в семи реках. Для рек Северо-Сахалинской равнины характерно пониженное колебание годового стока за счёт регулирующей деятельности почво-грунтов, озёр и болот (Атлас …, 1967), вследствие чего вода имеет коричневатый (гумусовый) цвет и сильно затрудняет подсчёт рыб. Кроме того, в силу слаборазвитой инфраструктуры (мало населённых пунктов и дорог) и заболоченности местности большинство участков рек труднодоступны для наблюдателей. По этим причинам вкупе с приблизительной оценкой нерестового фонда получаемые данные по заходам рыб носят ориентировочный характер, однако по ним всё же можно судить о межгодовой изменчивости заходов производителей, так как их численность рассчитывали по плотности скоплений рыб на обследованных участках. Анализируются данные начиная с 1998 г., так как в 1996−1997 гг. сотрудники Охинской КНС провели ревизию нерестилищ и изменили методику подсчёта числа производителей в реках.
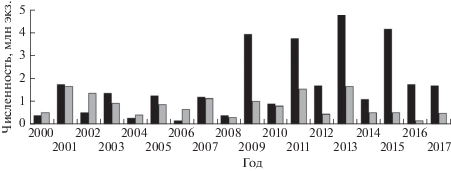
В 1998–2013 гг. в р. Лангры заходило от 70.6 до 361.0 тыс. производителей горбуши, от нереста которых скатывалось 1252−10191 тыс. мальков (рис. 3). Судя по соотношению этих величин, среднее число покатников от одного производителя у поколений чётных лет (M = 25.6, SD = 13.77, n = 8) в 1.5 раза превосходило таковое у поколений нечётных лет (M = 17.2, SD = 7.66, n = 8), но это различие статистически незначимое (F = 2.3). Не отмечено снижения численности покатников при увеличении заходов рыб в реку как следствия возможного переполнения нерестилищ. Анализ этой связи выполнен на примере заходов горбуши в 1993–2013 гг. в р. Лангры и последующего ската молоди от её нереста. С учётом неопределённостей, связанных с изменением методики учёта производителей, для характеристики их численности использована плотность скопления рыб, определяемая на обследуемых контрольных участках (рис. 4). В 2000–2017 гг. вылавливали в основном в прибрежных морских водах 158.9−4777.3 тыс. рыб, а в реки заходило 131.4−1643.5 тыс. производителей, при этом наблюдалась, хотя и статистически незначимая из-за короткого ряда наблюдений, но всё же положительная связь средней силы (r = 0.43, p > 0.05, n = 16) между уловами горбуши и её заходами в реки (рис. 5).
Рис. 3.
Динамика численности производителей (◼) горбуши Oncorhynchus gorbuscha в р. Лангры в 1998–2013 гг. и покатной молоди (—◻—) соответствующих поколений.
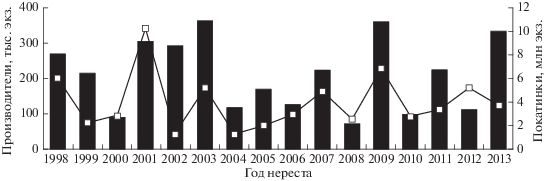
Рис. 4.
Соотношение между плотностью производителей и последующим скатом молоди на нерестилищах р. Лангры в 1993–2013 гг.: а − горбуша Oncorhynchus gorbuscha (R2 = 0.2591), б − кета O. keta (R2 = 0.1023).
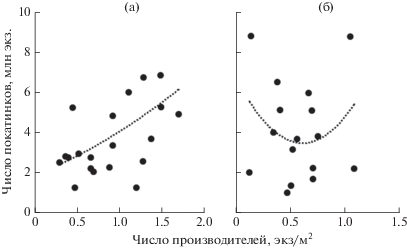
Рис. 5.
Численность горбуши Oncorhynchus gorbuscha в уловах (◼) и в заходах ( ) производителей в реки на северо-западном побережье Сахалина в 2000–2017 гг.
) производителей в реки на северо-западном побережье Сахалина в 2000–2017 гг.
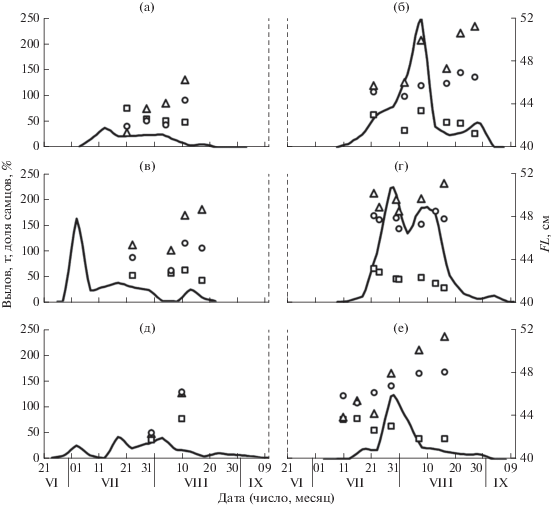
Судя по сезонной динамике уловов и биологическим показателям рыб, к северо-западному побережью Сахалина подходят разные группировки горбуши (рис. 6). В чётные годы (рис. 6а, 6в, 6д) её ход начинался раньше, первые уловы − в начале июля и даже в конце июня, причём в некоторые годы величина вылова в это время была довольно существенной, особенно в 2012 г. Во II декаде июля подходит горбуша с небольшими размерами тела, самки обычно крупнее самцов. В конце июля–начале августа появляется новая группировка, представленная сравнительно крупными рыбами, особенно самцами. Если численность рыб в этой группировке высока, то тенденция постепенного снижения доли самцов по мере хода нарушается: их доля при подходе этой группировки увеличивается, а затем вновь следует её постепенное сокращение, как это наблюдалось в 2011 и 2017 г. (рис. 6б, 6е); в 2013 г. почти весь ход горбуши был представлен рыбами этой поздней группировки (рис. 6г).
Рис. 6.
Динамика уловов и размерно-полового состава горбуши Oncorhynchus gorbuscha у северо-западного побережья Сахалина: а – 2010 г., б – 2011 г., в – 2012 г., г – 2013 г., д – 2014 г., е – 2017 г.; (——) – уловы, (◻) – доля самцов, (△) – длина (FL) самцов, (⚪) – FL самок.
Изменения численности кеты рассмотрим начиная с 2005 г., так как для периода 1998–2004 гг. отсутствуют статистические данные NPAFC для двух лет по заходам производителей в Лангры и для четырёх лет – во все реки района, соответственно, данные по скату молоди из рек побережья для этих лет имеют экспертный характер. Кроме того, с 2009 г. отменили основные ограничения на промысел кеты, в результате чего резко возросла величина уловов (рис. 2), которая стала определяться величиной подходов рыб в зону промысла. В 2005–2017 гг. в р. Лангры заходило 18.8−173.0 тыс. производителей, от нереста которых скатывалось 1683−8790 тыс. мальков, или 13−106 экз. в расчёте на одного производителя (M = 47.5, SD = 31.16). По значениям этого показателя рассчитана суммарная величина ската молоди из рек района (таблица). Расчёт выживаемости отдельных поколений в течение морского периода жизни невозможен из-за отсутствия данных по возрастному составу кеты. Судя по эпизодическим наблюдениям, в среднем 71.0% рыб возвращается на нерест на 4-м году жизни (Каев, 2003), т.е. по величине вылова и захода рыб в реки на 4-й год после нереста можно ориентировочно оценить выживаемость рыб в морских водах. Как видим, величина уловов взрослых рыб в некоторые годы вполне сопоставима с численностью покатной молоди (особенно для поколения 2011 г. рождения). С этим вполне согласуется также неадекватность уловов кеты по отношению к её заходам в реки, межгодовые изменения этих величин характеризуются довольно высоким отрицательным значением коэффициента корреляции (r = –0.61, p > 0.05, n = 9), статистическая незначимость которого обусловлена коротким периодом наблюдений. Отрицательная корреляция между этими параметрами отмечалась и в предыдущие годы (r = –0.41, p > 0.05, n = 9), хотя в этом случае её можно рассматривать и как следствие вводимых в те годы ограничений на промысел. Между численностью производителей на нерестилищах и последующим скатом молоди у кеты, в отличие от горбуши, наблюдалась весьма слабая корреляция (рис. 4б).
Таблица 1.
Показатели воспроизводства кеты Oncorhynchus keta в реках и численность её подходов к северо-западному побережью Сахалина в годы возврата рыб соответствующих поколений в возрасте 3+
| Годы нереста/возврата поколения в возрасте 3+ | Р. Лангры | Все реки | Подход рыб в год возврата поколения в возрасте 3+, тыс. экз. | ||||
|---|---|---|---|---|---|---|---|
| Заход, тыс. экз. | Скат, тыс. экз. | Скат/заход | Заход, тыс. экз. | Скат, тыс. экз. | Вылов | Заход в реки | |
| 2005/2009 | 173.0 | 2198 | 12.7 | 275.5 | 3137 | 1118.0 | 265.8 |
| 2006/2010 | 112.5 | 2220 | 19.7 | 179.2 | 3535 | 1169.9 | 160.3 |
| 2007/2011 | 82.0 | 3152 | 38.4 | 130.5 | 5017 | 1265.1 | 181.2 |
| 2008/2012 | 18.8 | 2000 | 106.4 | 29.9 | 3183 | 1016.0 | 100.3 |
| 2009/2013 | 167.0 | 8790 | 52.6 | 265.8 | 13990 | 1069.5 | 174.7 |
| 2010/2014 | 106.0 | 5977 | 56.4 | 160.3 | 9040 | 1974.0 | 180.9 |
| 2011/2015 | 112.4 | 1683 | 15.0 | 181.2 | 2714 | 2420.7 | 97.1 |
| 2012/2016 | 63.7 | 5111 | 80.2 | 100.3 | 8049 | 2714.5 | 62.8 |
| 2013/2017 | 110.8 | 5093 | 46.0 | 174.7 | 8033 | 1930.5 | 63.1 |
| 2014 | 120.5 | н.д. | н.д. | 180.9 | н.д. | – | – |
| 2015 | 57.8 | н.д. | н.д. | 97.1 | н.д. | – | – |
| 2016 | 36.9 | н.д. | н.д. | 62.8 | н.д. | – | – |
| 2017 | 41.8 | н.д. | н.д. | 63.1 | н.д. | – | – |
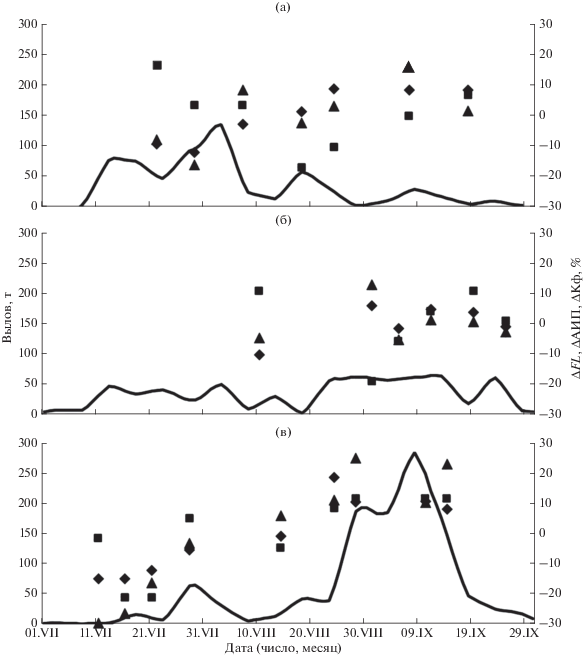
Неустойчивый характер сезонной динамики уловов кеты отчасти связан с погодными условиями. В то же время при анализе хода промысла в годы, когда проводили серии биологических анализов рыб, обращает на себя внимание то, что до наблюдавшегося во все годы снижения уловов в середине августа средняя длина рыб и плодовитость самок были меньше, чем в последующие дни августа и в сентябре. Кроме того, во все годы в пробах, собранных во второй половине августа или в начале сентября, значения КФ рыб были ниже, чем в собранных позже пробах. На рис. 7 эти показатели представлены в виде отклонений от средних значений текущего года (∆, %). В связи с существенными различиями диапазонов варьирования как средних значений анализируемых показателей, так и их дельт (FL − 54.6−65.0 см, ∆FL − –7.5−9.4%; АИП – 1743−3236 икринок, −29.8−25.1%; КФ − 1.41−1.55, −4.3−3.3%) для удобства восприятия данных при расчёте отклонений значения длины умножали на 2, а значения КФ − на 5, что позволило уравнять амплитуды изменения значений разных показателей. Наличие тенденций в их изменениях указывает на подход разных группировок кеты в течение её промысла.
Рис. 7.
Динамика уловов и биологических показателей кеты Oncorhynchus keta у северо-западного побережья Сахалина: а – 2010 г., б – 2012 г., в – 2017 г.; (——) – уловы; отклонения биологических показателей от среднего значения текущего года: (◆) – длина (FL), (◼) – коэффициент упитанности по Фультону (КФ), (▲) – абсолютная индивидуальная плодовитость (АИП).
Как и в других районах Сахалина, на северо-западном побережье острова горбуша широко расселена по рекам, а нерестовый ареал кеты сравнительно локализован. Однако этот район отличается от прочих весьма низкой эффективностью нереста обоих видов, о которой можно судить по отношению числа покатников к числу производителей в реках, т.е. по числу скатывающихся мальков в расчёте на одного производителя. По многолетним данным, на северо-восточном побережье Сахалина в реках Мелкая и Даги скатывалось в среднем по 89 и 67 мальков (Каев, Геращенко, 2008), в реках Дудинка, Бахура, Кура и Лютога – 72, 66, 69 и 77 мальков (Каев и др., 2004), а на Итурупе в р. Рыбацкая – 160 мальков горбуши (Kaev et al., 2006). Аналогичных наблюдений за результатами нереста кеты значительно меньше: в реках Тымь (северо-восточное побережье Сахалина), Илюшина (Кунашир) и Рыбацкая скатывалось соответственно по 156, 377 и 318 мальков (Каев, 2003; Каев, Ромасенко, 2017). Это в 3–8 раз больше в сравнении с р. Лангры в отношении кеты (по 47 мальков) и горбуши (по 21 мальку). Имеющиеся данные по эффективности воспроизводства обоих видов в речных водах не позволяют оценить оптимум пропуска производителей на нерестилища. У горбуши не наблюдались столь большие заходы производителей, в результате нереста которых стабилизировалось или даже уменьшалось количество покатников как следствие действия факторов, зависимых от плотности. У кеты также мал диапазон варьирования захода производителей. При этом у обоих видов отмечается большая изменчивость численности покатников, приходящихся на одно и то же число производителей, что и определяет слабую связь этих показателей, особенно у кеты (рис. 4).
В целом низкий уровень эффективности воспроизводства горбуши и кеты в реках северо-западного побережья связан, видимо, с заболоченностью местности. На первый взгляд, иначе обстоит ситуация в реках на п-ове Шмидта (слабо гористая прибрежная местность), что и послужило основанием для заключения о высокой эффективности нереста горбуши в этом районе без приведения каких-либо фактов (Живоглядов и др., 2017). Отчасти, видимо, это так, хотя и здесь основная часть русла наиболее крупных рек, таких как Тумь и особенно Пильво (соответственно 65 и 295 тыс. м2 нерестилищ), протекает по заболоченной местами долине, простирающейся в меридианном направлении по срединной части всего полуострова. По данным Сахалинрыбвода, при синхронных наблюдениях в 2010–2015 гг. плотность заполнения нерестилищ в р. Пильво была несколько выше, чем в р. Лангры (0.78 против 0.65 экз./м2), однако обследование бассейна Лангры значительно сложнее ввиду огромной площади её бассейна (1190 против 250 км2). Не вносит ясность в данный вопрос и сравнение имеющихся данных по малым водотокам, плотность заполнения нерестилищ в которых в одни годы была выше на п-ове Шмидта, в другие – в остальных районах побережья.
Статистика уловов лососей также не указывает на повышенную концентрацию рыб у п-ова Шмидта, вдоль которого, например, в 2016–2017 гг. было выловлено в среднем 1% кеты и 16% горбуши от улова этих рыб на северо-западном побережье острова, в то время как в реках полуострова сосредоточено примерно 45% нерестилищ горбуши. Вдоль остального островного побережья Сахалинского залива, в основном в его западной части, прилегающей к мысу, разделяющему Сахалинский залив и Амурский лиман, вылавливали 22% горбуши и 15% кеты. На островном побережье Амурского лимана севернее впадения р. Лангры – 59% горбуши и 82% кеты, оставшуюся часть (3% горбуши и 2% кеты) – в основном в районе р. Лангры. Столь большие уловы вблизи указанного мыса являются следствием вылова здесь лососей, мигрирующих через Амурский лиман в реки сахалинского и материкового побережья. В первую очередь это относится к кете, ведь большинство её нерестилищ сосредоточено в р. Лангры. Причём, судя по тесной связи местных уловов с уловами этого вида в Амуре (r = 0.90, p < 0.001, n = 9) в сочетании с показанной выше отрицательной корреляцией уловов с заходами производителей в реки, основу вылова составляют рыбы, мигрирующие в реки материкового побережья. Судя по небольшим размерам рыб и малой плодовитости самок (рис. 7а−7в), что характерно для амурской летней кеты (Рослый, 2002), эти рыбы формируют основу уловов примерно до середины августа (к настоящему времени нет свидетельств о массовом нересте летней расы кеты в реках рассматриваемого побережья Сахалина). Затем начинается ход более крупной и плодовитой осенней кеты. По результатам мечения рыб из промысловых уловов в прибрежье (несколько севернее впадения р. Лангры) установлено, что сначала в подходах представлены рыбы с наличием внешних брачных изменений, происходящие в основном из рек сахалинского (33−44%) и материкового побережья Амурского лимана, а с конца I декады сентября по численности доминировала уже собственно амурская осенняя кета (86−100%). Эти рыбы отличались не только серебристым окрасом, но и высокой скоростью передвижения, судя по местам и срокам их вторичной поимки в русле Амура (Каев, Рослый, 1987). По биологическим показателям начало хода этих сравнительно высокотелых рыб тестируется повышенными значениями коэффициента упитанности, в соответствии с которыми их доминирование в подходах в 2017 г. наблюдалось уже с конца августа (рис. 7в), что связано, видимо, не с более ранними подходами, а с их сравнительно высокой численностью в этом году.
В отношении горбуши не приведены данные по соотношению между численностью покатников и последующим возвратом взрослых рыб соответствующих поколений вследствие их чрезмерно высоких значений в сравнении с таковыми в других районах региона. Так, выживаемость в течение морского периода жизни у горбуши о-ва Итуруп в среднем составляет 4.8% (от 3.4 до 8.0% для отдельных поколений), зал. Анива – 4.4 (1.8−9.6)%, юго-восточного побережья Сахалина – 6.2 (1.8−13.1)% (Kaev, Irvine, 2016). Тем не менее важно подчеркнуть, что соотношение суммарного количества выловленных и зашедших в реки рыб, а также покатной молоди статистически значимо (F = 10.3, p < 0.01) выше у поколений нечётных лет (M = 21.1, SD = 8.02, n = 8) в сравнении с поколениями чётных лет (M = 10.2, SD = 5.28, n = = 8). Это косвенно может свидетельствовать о лучшей выживаемости рыб поколений нечётных лет нереста, вполне согласующейся с более высокими уловами в нечётные годы. На первый взгляд, столь высокие значения данного соотношения свидетельствуют о массовом прилове у северо-западного побережья Сахалина горбуши, происходящей из рек сопредельного материкового побережья. Однако из сопоставления уловов горбуши и кеты по разным участкам побережья следует, что доля прилова горбуши, происходящей из рек сопредельного материкового побережья, существенно ниже, так как её сравнительно большие уловы у побережья Сахалинского залива в районе впадения многих нерестовых рек позволяют полагать о местном происхождении облавливаемых рыб. Такое предположение было сделано уже при первом обобщении имеющихся данных по воспроизводству горбуши в этом районе (Иванов и др., 2001).
Весомым аргументом в пользу преимущественно местного происхождения горбуши в уловах является несовпадение их межгодовых колебаний на разных побережьях: по статистике NPAFC в 2003–2017 гг. в Амуре в нечётные годы вылавливали в среднем 1.78 (0.70−3.38) тыс. т, в чётные − 8.39 (1.38−23.10) тыс. т, тогда как на северо-западном побережье Сахалина доминантными по уровню уловов являются нечётные годы (рис. 2). То есть горбуша амурского происхождения может оказывать сколь-нибудь заметное влияние на величину уловов в рассматриваемом районе Сахалина лишь в чётные годы, но такое влияние невелико, судя по величине и соотношению уловов этого вида здесь, в чётные и нечётные годы. Не исключено, что на островном побережье Сахалинского залива может облавливаться какая-то часть горбуши, мигрирующей к охотоморскому побережью Хабаровского края. Для этой горбуши, как и на Сахалине, характерна более высокая численность в нечётные годы, однако плотность её миграционного потока невелика, если судить по её общей численности (в нечётные и чётные 2001–2016 гг. соответственно 9.6 и 1.3 млн экз.) и огромной протяжённости данного побережья Хабаровского края. Таким образом, необычно большое соотношение между численностью покатной молоди и взрослых рыб у горбуши северо-западного побережья Сахалина связано, видимо, с пробелами в сборе первичных материалов. Учитываемый скат молоди в Лангры (крупнейшая река в районе) экстраполируется на более малые реки и ручьи согласно имеющимся данным по площади нерестилищ и заходам производителей; к тому же есть ещё ряд малых водотоков, для которых такие данные отсутствуют. Это может иметь существенное значение, судя по тому, что в других районах региона в небольших водотоках наблюдается более высокая плотность заполнения нерестилищ производителями горбуши и даже более высокая эффективность их нереста (Каев и др., 2010; Каев, Ромасенко, 2017).
Изменения в сезонной динамике уловов горбуши связаны с ходом её разных группировок. Первой в Амурский лиман подходит с юга из Татарского пролива япономорская горбуша (Енютина, 1972), уловы которой (примерно до начала II декады июля) у сахалинского побережья малы и носят случайный характер, так как не связаны с численностью подходов этой группировки. Массовый заход этих рыб в Амурский лиман начался с 2012 г., когда резко возросла численность япономорской горбуши в чётные годы, судя по её уловам на материковом и сахалинском побережье Татарского пролива. Именно с этого года обозначились высокие уловы этих рыб в устье Амура в июне 2012, 2014, 2016 гг. − соответственно 1491, 4499 и 6372 т (16.1, 30.2 и 27.6% суммарных уловов в эти годы в данном районе). В эти же годы возросли уловы в конце июня и в начале июля у побережья Сахалина, но их динамика была иной (соответственно 915, 160 и 203 т). Нет достоверных свидетельств о заходе япономорской горбуши в реки сахалинского побережья лимана, так как под приводимыми данными о её заходе вплоть до начала августа понимаются рыбы, мигрирующие в лиман с севера (Иванов и др., 2001). Судя по биологическим показателям, две волны подхода горбуши с севера связаны с последовательной миграцией её ранней и поздней темпоральных форм охотоморской группировки, различающихся по длине тела самцов и самок; подход поздней формы тестируется по увеличению длины рыб, особенно самцов, которые становятся крупнее самок (Каев, 2012). Степень выраженности в это время увеличения доли самцов зависит от соотношения численности рыб этих форм.
Вопрос, почему в уловах лососей у сахалинского побережья транзитные рыбы в основном представлены кетой, а не горбушей, может быть объяснён так. В лимане имеются два протяжённых русла (прямое продолжение рукавов Амура), называемых Сахалинским и Северным фарватерами, по которым в основном передвигаются амурские лососи (Рослый, 2002). Кета, мигрируя большей частью в Амур, использует Сахалинский фарватер, который вскоре отклоняется от побережья Сахалина к центральной части лимана, а горбуша, видимо, использует в основном Северный фарватер, проходящий вдоль материкового побережья лимана, в массе мигрируя в реки этого побережья.
Таким образом, судя по динамике уловов и биологическим показателям рыб, промысел горбуши и кеты базируется на подходах разных группировок этих видов. Если у кеты наличие группировок связано с ходом рыб разных рас (летняя и осенняя) и разного происхождения (из Амура и небольших рек северо-западного побережья Сахалина и сопредельного материкового побережья), то у горбуши – в основном за счёт подхода рыб местного происхождения, но разных темпоральных форм. Невысокая эффективность воспроизводства горбуши и кеты в местных реках, судя по соотношению численности производителей на нерестилищах и последующего ската молоди, вполне соответствует небольшим уловам рыб местных стад этих видов лососей. Выживаемость поколений в течение морского периода жизни адекватно оценить на данном этапе исследований не представляется возможным вследствие массового присутствия в уловах транзитных рыб (кета) и наличия неопределённостей при расчётах суммарной численности молоди, скатывающейся из всех рек побережья (горбуша).
Список литературы
Атлас Сахалинской области. 1967. М.: ГУ геодезии и картографии при Совмине СССР, 135 с.
Воловик С.П. 1967а. Структура нерестовых стад и эффективность естественного воспроизводства горбуши на Южном Сахалине: Автореф. дис. … канд. биол. наук. Калининград: КТИРПиХ, 25 с.
Воловик С.П. 1967б. Методы учета и некоторые особенности поведения покатной молоди горбуши в реках Сахалина // Изв. ТИНРО. Т. 61. С. 104–117.
Гриценко О.Ф. 1990. Популяционная структура сахалинской горбуши Oncorhynchus gorbuscha // Вопр. ихтиологии. Т. 30. Вып. 5. С. 825–835.
Енютина Р.И. 1972. Амурская горбуша (промыслово-биологический очерк) // Изв. ТИНРО. Т. 77. С. 3–126.
Живоглядов А.А., Игнатьев Ю.И., Живоглядова Л.А. 2017. Динамика численности и условия воспроизводства массовых видов тихоокеанских лососей (Oncorhynchus) северо-западного побережья Сахалина // Вопр. ихтиологии. Т. 57. № 4. С. 435–444.
Иванков В.Н. 1967. Локальные стада горбуши Курильских островов // Гидробиол. журн. Т. 3. № 1. С. 62–67.
Иванков В.Н. 1970. Изменчивость и внутривидовая дифференциация кеты // Там же. Т. 6. № 2. С. 106–112.
Иванков В.Н. 1972. Особенности экологии и структура популяций осенней кеты различных районов Сахалина // Уч. зап. ДВГУ. Т. 60. Фауна и рыбохозяйственное значение прибрежных вод северо-западной части Тихого океана. С. 27–35.
Иванков В.Н. 1993. Популяционная организация у тихоокеанских лососей с коротким пресноводным периодом жизни // Вопр. ихтиологии. Т. 33. Вып. 1. С. 78–83.
Иванков В.Н. 2011. Внутривидовая дифференциация и популяционная организация горбуши Oncorhynchus gorbuscha в различных частях ареала // Изв. ТИНРО. Т. 167. С. 64–76.
Иванкова Е.В., Борисовец Е.Э., Карпенко А.И., Хоревин Л.Д. 2000. Популяционная структура кеты Oncorhynchus keta острова Сахалин // Вопр. ихтиологии. Т. 40. № 4. С. 467–476.
Иванов А.Н., Шершнев А.П., Иванова Л.В. 2001. О функциональной структуре горбуши северо-западного Сахалина // Чт. памяти В.Я. Леванидова. Вып. 1. С. 310–322.
Каев А.М. 2003. Особенности воспроизводства кеты в связи с ее размерно-возрастной структурой. Южно-Сахалинск: Изд-во СахНИРО, 288 с.
Каев А.М. 2011. Оценка эффективности прогнозирования и управления промыслом горбуши в Сахалино-Курильском регионе // Изв. ТИНРО. Т. 167. С. 32–53.
Каев А.М. 2012. Темпоральная структура и некоторые вопросы динамики стада горбуши Oncorhynchus gorbuscha (Salmonidae) // Вопр. ихтиологии. Т. 52. № 1. С. 62–71.
Каев А.М., Геращенко Г.В. 2008. Показатели воспроизводства горбуши северо-восточного побережья острова Сахалин // Реализация «Концепции дальневосточной бассейновой программы изучения тихоокеанских лососей». Бюл. № 3. С. 102–106.
Каев А.М., Игнатьев Ю.И. 2015. Развитие заводского разведения тихоокеанских лососей в Сахалино-Курильском регионе и его значение для промысла // Тр. ВНИРО. Т. 153. С. 95–104.
Каев А.М., Ромасенко Л.В. 2017. Горбуша и кета острова Кунашир (структура популяций, воспроизводство, промысел). Южно-Сахалинск: Изд-во СахГУ, 124 с.
Каев А.М., Рослый Ю.С. 1987. Мечение осенней кеты в лимане реки Амур // Рыб. хоз-во. № 2. С. 21–23.
Каев А.М., Антонов А.А., Ким Хе Юн, Руднев В.А. 2004. Показатели воспроизводства горбуши Oncorhynchus gorbuscha южной части острова Сахалин // Тр. СахНИРО. Т. 6. С. 3–38.
Каев A.M., Сафронов С.Н., Никитин В.Д. и др. 2010. Подходы к созданию лососевых рыбохозяйственных заповедных зон в Сахалинской области // Лососевые рыбохозяйственные заповедные зоны на Дальнем Востоке России. М.: Изд-во ВНИРО. С. 51–59.
Макоедов А.Н., Коротаев Ю.А., Антонов Н.П. 2009. Азиатская кета. Петропавловск-Камчатский: Изд-во КамчатНИРО, 356 с.
Рослый Ю.С. 2002. Динамика популяций и воспроизводство тихоокеанских лососей в бассейне Амура. Хабаровск: Хабар. книж. изд-во, 210 с.
Уловы тихоокеанских лососей, 1900–1986 гг. 1989. М.: Изд-во ВНИРО, 213 с.
Irvine J.R., Tompkins A., Saito T. et al. 2012. Pacific salmon status and abundance trends − 2012 update // NPAFC Doc. № 1422. Rev. 2. 90 p. (Available at http://www.npafc.org)
Kaev A.M., Irvine J.R. 2016. Population dynamics of pink salmon in the Sakhalin-Kuril Region, Russia // Bull. NPAFC. № 6. P. 297–305.
Kaev A.M., Klovach N.V. 2014. Revision of data on pink salmon abundance in East Sakhalin and Kuril Islands // NPAFC Doc. № 1501. 11 p.
Kaev A.M., Chupakhin V.M., Kruchinin M.Y. 2006. Reproduction indices of the Iturup Island pink salmon (Kuril Islands) // NPAFC Doc. № 977. 18 p.
Kaev A.V., Antonov A.A., Chupakhin V.M., Rudnev V.A. 2007. Possible causes and effects of shifts in trends of abundance in pink salmon of Southern Sakhalin and Iturup Islands // Bull. NPAFC. № 4. P. 223–233.
NPAFC. 2018. North Pacific Anadromous Fish Commission. (http://www.npafc.org/new/pub_documents.html)
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии