

Вопросы ихтиологии, 2019, T. 59, № 5, стр. 523-530
Морфологическая изменчивость пятнистого круглопёра Eumicrotremus pacificus (Cottoidei, Cyclopteridae)
О. С. Воскобойникова 1, *, А. А. Баланов 2
1 Зоологический институт РАН – ЗИН
Санкт-Петербург, Россия
2 Национальный научный центр морской биологии Дальневосточного отделения РАН –ННЦМБ ДВО РАН
Владивосток, Россия
* E-mail: vosk@zin.ru
Поступила в редакцию 29.10.2018
После доработки 15.01.2019
Принята к публикации 15.01.2019
Аннотация
Исследована изменчивость морфометрических признаков, строения сейсмосенсорной системы и наружного скелета у самок, не участвующих в нересте самцов и нерестовых самцов пятнистого круглопёра Eumicrotremus pacificus. Самки отличаются от самцов бóльшим антеанальным расстоянием и стабильностью строения наружного скелета на протяжении всего взрослого периода жизни. Нерестовые самцы характеризуются существенным изменением ряда пластических признаков и строения наружного скелета, по-видимому, связанным с особенностями биологии нереста видов Eumicrotremus. В строении осевого скелета, скелета непарных плавников и сейсмосенсорной системы головы E. pacificus заметная изменчивость не выявлена. Подтверждена возможность использования признаков внешней морфологии и наружного скелета самок и не вступивших в нерест самцов в систематике круглопёрых рыб.
В последнее время появилось значительное число работ, посвящённых систематике круглопёрых рыб семейства Cyclopteridae, выполненных с использованием морфологических и молекулярных методов исследования (Byrkjedal et al., 2007; Kai et al., 2015; Lee et al., 2015, 2017; Stevenson et al., 2017). На основании сходства в последовательностях митохондриальных генов cyt b и COI отдельные виды родов Cyclopteropsis, Lethotremus и Eumicrotremus сведены в этих работах в синонимию, а также высказано предположение о том, что и остальные виды рода Cyclopteropsis представляют собой самцов разных видов Eumicrotremus и являются их младшими синонимами. Расхождения сделанных выводов с морфологическими данными объясняются в этих работах проявлением полового диморфизма или существенной морфологической изменчивостью ряда признаков. В первую очередь к ним относятся степень развития наружного скелета и форма 1-го спинного плавника (Kai et al., 2015; Stevenson et al., 2017).
Цель нашего исследования – установить пределы изменчивости морфологических признаков у всех взрослых особей и отдельно у самцов и самок пятнистого круглопёра Eumicrotremus pacificus Schmidt, 1904. Выбор этого вида продиктован выразительностью его диагностических признаков, комплекс которых чётко обособляет его от других видов рода Eumicrotremus (Шмидт, 1904; Линдберг, Легеза, 1955; Ueno, 1970).
МАТЕРИАЛ И МЕТОДИКА
В исследовании использованы материалы из фондовых коллекций ЗИН РАН и ННЦМБ ДВО РАН.
ЗИН № 12921 – лектотип Eumicrotremus pacificus SL 62.6 мм, Охотское море, зал. Анива, трал 24, 28–30 м, 28.08.1901 г., коллектор П.Ю. Шмидт. ЗИН № 52863 – паралектотипы, 3 экз. SL 19.2–29.0 мм, собраны вместе с лектотипом. ЗИН № 12922 – паралектотипы, личинки и мальки, 16 экз. SL 15.5–29.7 мм, собраны вместе с лектотипом. ЗИН № 25374 – 1 экз. SL 38.6 мм, зал. Петра Великого, март 1927 г., коллектор Миловидова. ЗИН № 26047 – 1 экз. SL 55.4 мм, открытое море к югу от м. Островной, 42°44′40″ c.ш. 133°43′40'' в.д., 18.09.1934 г., экспедиция ЗИН АН СССР, коллектор Г.У. Линдберг. ЗИН № 26482 – 3 экз. SL 49.4–60.2 мм, траулер “Аскольд”, Японское море у м. Ефстафия, 43°26′3'' с.ш. 135°03′15'' в.д., 29.06.1932 г., коллектор М. Кривобок. ЗИН № 33640 – 1 экз. SL 52.7 мм, НИС “Топорок”, ст. 78, бимтрал, Охотское море, зал. Терпения, 08.09.1947 г., коллектор Г.У. Линдберг. ЗИН № 33693 – 3 экз. SL 50.5–57.5 мм, Охотское море, зал. Анива, глубина 23 м, заросли красных водорослей, 15.07.1947 г., коллектор З. Петрова. ЗИН № 33694 – 11 экз. SL 15.7–38.8 мм, Охотское море, зал. Анива, глубина 23 м, заросли красных водорослей, 15.07.1947 г. ЗИН № 33703 – 4 экз. SL 40.3–46.2 мм, НИС “Топорок”, ст. 44, Охотское море, зал. Анива, глубина 25 м, 20.09.1947 г., коллектор З.И. Петрова. ЗИН № 33705 – 6 экз. SL 40.9–53.6 мм, НИС “Топорок”, ст. 123–124, оттертрал 35, Охотское море, б. Цитоси, глубина 19–27 м, 23.09.1947 г., коллектор Г.У. Линдберг. ЗИН № 33709 – 11 экз. SL 36.0–48.7 мм, НИС “Топорок”, ст. 35/36, оттертрал 19, море Неморо, глубина 87 м, 04.09.1948 г., коллектор Г.У. Линдберг. ЗИН № 33712 – 9 экз. SL 34.4–51.0 мм, НИС “Топорок”, ст. 34–35, оттертрал 16, море Неморо, глубина 78–85 м, 04.09.1948 г., коллектор Г.Б. Семенова. ЗИН № 33716 – 2 экз. SL 42.3 и 45.3 мм, НИС “Топорок”, ст. 48, оттертрал 26, море Неморо, глубина 100 м, 07.09.1948 г., коллектор Г.Б. Семенова. ЗИН № 34778 – 2 экз. SL 37.2 и 72.7 мм, э/с “Гидрограф”, Японское море, 42°51′3″ с.ш. 132°00′в.д., глубина 77 м, 16.11.1925 г. ЗИН № 41700 – 2 экз. SL 486 и 55.4 мм, Японское море, зал. Петра Великого у о-ва Попова, глубина 70–80 м, 05.07.1973 г., коллектор М.Б. Ильина. ЗИН № 42505 – 1 экз. SL 67.7 мм, МРС 254, Японское море, зал. Петра Великого, 15.08.1958, коллекторы Е.А. Дорофеева, М.И. Легеза. ЗИН № 42506 – 2 экз. SL 31.3 и 64.3 мм, МРС 254, зал. Петра Великого, у м. Гамова, 29.07.1958 г., коллекторы Е.А. Дорофеева, М.И. Легеза. ЗИН № 47927 – 2 экз. SL 57.3 и 80.7 мм, Японское море, зал. Петра Великого, у о-ва Большой Пелис, глубина 60 м, 18.06.1979 г., коллектор А. Маркевич. ЗИН № 53916 – 1 экз. SL 59 мм, НИС “Профессор Кагановский”, трал 32, донный трал ДТ69.0, 43°28′ с.ш. 135°04′ в.д., глубина 130 м, 30.05.1996 г., коллектор Е.Н. Ильинский. ЗИН № 53917 – 1 экз. SL 70.0 мм, НИС “Профессор Кагановский”, трал 33, Японское море, Приморье, 43°27' с.ш. 135°06′ в.д., глубина 69 м, 30.05.1996 г., коллектор Е. Ильинский. ЗИН № 55982 – 1 экз. SL 48.3 мм, РТ “Милоградово”, трал 180, Японское море, 45°36′ с.ш. 137°35′ в.д., глубина 40 м, 24.06.1970 г., коллектор В.В. Федоров.
ННЦМБ № Ep1-10 – 1 экз., РТ “Бухоро”, Японское море, Северное Приморье, трал 107, 46.4° с.ш. 137.2° в.д., глубина 65 м, 04.11.2010 г., коллекторы Д.В. Антоненко, В. Панченко. ННЦМБ №№ Ep5-11, Ep6-11, Ep7-11 – 3 экз., РТ “Профессор Пробатов”, Охотское море, зал. Анива, трал 46, 46.3° с.ш. 143.0° в.д., глубина 25 м, 10.06.2011 г., коллектор А.А. Баланов. ННЦМБ № Epa5-11 – 1 экз., РТ “Профессор Пробатов”, Охотское море, зал. Анива, трал 7, 49.0° с.ш. 144.0° в.д., глубина 46 м, 20.07.2011 г., коллектор А.А. Баланов. ННЦМБ № Epa 8-11 – 1 экз., РТ “Профессор Пробатов”, Охотское море, зал. Анива, трал 7, 49.0° с.ш. 143.4° в.д., глубина 40 м, 21.07.2011 г., коллектор А.А. Баланов.
Всего исследовано 75 экз. молоди, самцов (нерестовых и не вступивших в нерест) и самок E. pacificus. Нерестовыми самцами мы называем тех, которые были собраны во время охраны гнезда и отличаются от остальных самцов заметной редукцией наружного скелета. К ним относятся материалы из коллекции ННЦМБ ДВО РАН, а также ЗИН № 33640. Преднерестовые самцы также характеризуются частичной редукцией наружного скелета. Причина частичной утраты наружного скелета, по-видимому, заключается в избегании самцами определённых неудобств при уходе за кладкой, находящейся в пустой раковине моллюсков рода Neptunea. Один из таких самцов (ЗИН № 33640, SL 52.7 мм) был идентифицирован Линдбергом и Легезой (1955) как C. bergi Popov, 1929, причём авторы указывали на его промежуточное положение между этим видом и E. pacificus. Более подробное изучение этого экземпляра показало, что он имеет характерные диагностические признаки E. pacificus, и он был нами переопределён как E. pacificus. В нашем исследовании самцы, находящиеся в разном состоянии (не вступившие в нерест (далее – самцы), нерестовые и лектотип E. pacificus), рассматриваются отдельно.
Измерения проводили штангенциркулем с точностью до 0.1 мм по ранее разработанной методике для круглопёрых (Воскобойникова, Назаркин, 2015). Измерения приведены в процентах стандартной длины (SL). Число костных бляшек подсчитывали в надглазничном, межглазничном, заглазничном, окологрудном, зажаберном и срединном затылочном рядах по методике Линдберга и Легезы (1955). Для определения степени редукции наружного скелета определяли наличие или отсутствие костных бляшек на голове, в хвостовом отделе туловища и на хвостовом стебле. Отдельно была исследована последовательность появления костных бляшек у личинок и молоди. Число плавниковых лучей подсчитывали по рентгенограммам. Изготовлены и исследованы рентгенограммы 21 экз. E. pacificus (ЗИН № 33693, 33705, 33709, 41700, 42927), а также рентгенограммы 6 экз. из коллекции ННЦМБ ДВО РАН. В тексте используются следующие сокращения: SL – стандартная длина тела, с – длина головы; D, A, P, V, C – соответственно спинной, анальный, грудной, брюшной и хвостовой плавники; каналы сейсмосенсорной системы: CSO – надглазничный, CIO – окологлазничный, CT – темпоральный, CPM – предкрышечно-нижнечелюстной.
Пределы изменчивости, средние значения и ошибка среднего установлены с использованием программы Microsoft Excel. Нормальность распределения признаков проверена по критерию Колмогорова–Смирнова. Расчёт достоверности различий признаков выполнен по t-критерию Стъюдента в программе Statistica (версия 10).
РЕЗУЛЬТАТЫ
Измерения. Пластические признаки 50 экз. E. pacificus приведены в табл. 1. В наиболее широких пределах варьируют: расстояние от переднего конца рыла до основания P (20.9–50.1%), антеанальное расстояние (61.6–82.5%), первое (24.6–52.1%) и второе (48.5–79.7%) антедорсальные расстояния, высота D1 (17.3–46.3%), максимальная ширина тела (28.5–59.8%). Наибольшие расхождения по средним значениям между самками, самцами, лектотипом, с одной стороны, и нерестовыми самцами – с другой стороны выявлены в расстоянии от конца рыла до переднего края диска (29.6–33.5 против 39.3%). Среднее значение антеанального расстояния оказалось наибольшим у самок.
Таблица 1.
Пластические признаки Eumicrotremus pacificus
| Признак | Самки (15 экз.) | Самцы | Все экземпляры (50 экз.) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| не вступившие в нерест (27 экз.) | нерестовые (8 экз.) | лектотип | |||||||||||
| M | m | min–max | M | m | min–max | M | m | min–max | M | m | min–max | ||
| SL, мм | 39.0–80.7 | 36.0–57.5 | 52.7–63.2 | 62.6 | 36.0–80.7 | ||||||||
| В % SL | |||||||||||||
| aP | 39.0 | 8.83 | 20.9–50.1 | 41.4 | 4.58 | 24–47.7 | 39.5 | 4.11 | 31.2–44.1 | 41.1 | 40.4 | 6.72 | 20.9–50.1 |
| a–disk | 32.1 | 5.94 | 24.7–44.0 | 29.6 | 3.74 | 22.6–39.9 | 39.3 | 3.19 | 32.2–42.8 | 33.5 | 31.8 | 4.33 | 22.6–44.0 |
| l disk | 31.2 | 5.89 | 20.1–45.1 | 33.5 | 6.34 | 20.3–44.4 | * | 28.1 | 32.6 | 6.18 | 20.1–45.1 | ||
| disk–anus | 20.0 | 3.90 | 14.4–27.6 | 19.0 | 3.48 | 10.3–27.3 | 20.8 | 3.59 | 14.3–25.0 | 16.8 | 19.5 | 3.62 | 10.3–27.6 |
| anus–A | 15.7 | 1.70 | 12.1–18.6 | 13.9 | 2.13 | 10.5–17.3 | 16.1 | 2.61 | 11.4–21.1 | 17.6 | 14.8 | 2.06 | 10.5–21.1 |
| aA | 79.6 | 4.46 | 72.5–82.5 | 73.4 | 5.04 | 61.6–79.9 | 75.8 | 1.39 | 74.0–77.9 | 71.9 | 75.6 | 4.34 | 61.6–82.5 |
| aD1 | 41.0 | 3.36 | 36.1–47.5 | 40.4 | 6.41 | 24.6–52.1 | 39.7 | 3.04 | 34.1–44.8 | 39.6 | 40.5 | 4.99 | 24.6–52.1 |
| aD2 | 69.6 | 6.06 | 53.1–79.7 | 67.5 | 6.23 | 48.5–74.4 | 70.8 | 3.44 | 66.1–76.4 | 69.6 | 68.6 | 5.78 | 48.5–79.7 |
| c | 39.8 | 1.40 | 39.7–42.3 | 40.7 | 2.94 | 32.6–46.0 | 40.9 | 1.41 | 38.2–42.1 | 40.6 | 40.5 | 2.25 | 32.6–46.0 |
| ao | 13.7 | 1.75 | 9.7–15.9 | 14.6 | 2.19 | 10.3–18.9 | 15.6 | 1.48 | 12.9–17.4 | 16.8 | 14.5 | 1.95 | 9.7–18.9 |
| o | 11.8 | 1.30 | 9.5–14.4 | 12.4 | 1.54 | 10.1–16.1 | 13.3 | 1.21 | 12.1–15.8 | 12.0 | 12.3 | 1.42 | 9.5–16.1 |
| po | 18.1 | 1.23 | 16.5–20.3 | 17.6 | 2.12 | 13.6–22.8 | 18.8 | 4.10 | 16.1–29.4 | 16.9 | 17.9 | 2.13 | 13.6–29.4 |
| lP | 24.9 | 2.75 | 19.5–28.5 | 23.4 | 3.23 | 17.9–36.5 | 26.9 | 2.35 | 22.4–30.9 | 22.4 | 23.6 | 2.96 | 17.9–36.5 |
| lC | 22.4 | 1.65 | 19.9–24.3 | 22.6 | 2.19 | 18.5–27.7 | 22.1 | 1.98 | 19.9–25.9 | – | 22.5 | 1.99 | 18.5–27.7 |
| hD1 | 29.3 | 6.07 | 20.6–37.4 | 30.1 | 6.68 | 17.3–46.3 | 25.9 | 2.11 | 24.3–31.1 | 27.1 | 29.2 | 5.84 | 17.3–46.3 |
| hD2 | 18.3 | 2.86 | 13.0–22.8 | 18.0 | 2.56 | 14.5–27.2 | 18.2 | 1.92 | 14.6–21.0 | 16.8 | 18.1 | 2.56 | 13.0–27.2 |
| hA | 17.9 | 2.37 | 14.2–22.8 | 17.2 | 3.30 | 11.8–26.1 | 18.4 | 2.06 | 14.6–21.2 | 15.2 | 17.5 | 2.84 | 11.8–26.1 |
| Hc | 39.1 | 2.67 | 35.0–43.0 | 38.3 | 3.68 | 30.2–45.1 | 42.6 | 2.81 | 38.5–45.8 | 42.0 | 39.2 | 3.35 | 30.2–45.8 |
| H | 50.5 | 2.72 | 46.5–56.9 | 47.6 | 4.48 | 37.3–55.4 | 52.5 | 3.50 | 48.6–58.1 | 55.9 | 49.3 | 3.80 | 37.3–58.1 |
| HA | 34.7 | 3.15 | 28.3–40.2 | 33.5 | 3.38 | 25.4–39.5 | 38.6 | 1.89 | 35.9–41.2 | 38.6 | 34.7 | 3.10 | 25.4–41.2 |
| hpc | 9.2 | 0.98 | 6.8–10.3 | 9.6 | 1.18 | 7.5–11.9 | 11.4 | 0.72 | 9.8–12.2 | 10.1 | 9.7 | 0.98 | 6.8–12.2 |
| wc | 53.6 | 3.80 | 42.1–57.5 | 52.6 | 4.32 | 42.4–60.0 | 53.1 | 4.73 | 46.2–61.7 | 54.6 | 53.0 | 4.2 | 42.1–61.7 |
| W | 47.9 | 8.22 | 28.5–59.8 | 46.9 | 4.63 | 36.7–56.7 | 44.3 | 4.88 | 35.4–53.7 | 52.7 | 46.9 | 6.28 | 28.5–59.8 |
| io | 34.7 | 3.15 | 20.8–31.9 | 25.6 | 3.34 | 20.7–32.9 | 25.6 | 1.94 | 22.5–29.0 | 25.5 | 28.3 | 2.17 | 20.8–32.9 |
| lmx | 17.2 | 1.21 | 15.4–19.2 | 18.2 | 1.85 | 14.8–23.3 | 19.7 | 1.34 | 18.1–21.6 | 16.6 | 18.1 | 1.58 | 15.4–23.3 |
| В % c | |||||||||||||
| ao | 34.5 | 4 | 25.6–40.6 | 35.9 | 5.1 | 25.8–47.1 | 38.1 | 4.6 | 30.8–45.5 | 41.4 | 35.9 | 4.7 | 25.6–45.5 |
| o | 29.6 | 2.77 | 28.9–34.7 | 30.6 | 3.8 | 25.7–42 | 32.4 | 2.6 | 29.2–37.5 | 29.6 | 30.6 | 3.3 | 25.7–42 |
| po | 45.6 | 1.19 | 44–50.6 | 43.3 | 4.3 | 37.7–51.9 | 42.8 | 2.2 | 41.2–46.1 | 41.6 | 43.9 | 3.03 | 37.7–51.9 |
| io | 64.7 | 7.26 | 55.3–81.4 | 63.1 | 7.4 | 53–75.6 | 62.8 | 4.9 | 53.7–68.9 | 62.8 | 63.5 | 6.96 | 53.7–81.4 |
Примечание. Здесь и в табл. 2: SL – стандартная длина, aP – расстояние от переднего конца рыла до основания грудного плавника, a–disk – от переднего конца рыла до переднего края внутреннего диска, l disk – длина наружного диска, disk–anus – расстояние от заднего края внутреннего диска до ануса, anus–A – от ануса до начала анального плавника, aA – антеанальное расстояние, aD1 и aD2 – 1-е и 2-е антедорсальное расстояние, c – длина головы, ao – длина рыла, o – диаметр глаза, po – заглазничное расстояние, lP – длина грудного плавника, lC – длина хвостового плавника, hD1 – высота D1, hD2 – высота D2, hA – высота анального плавника, Hс – высота головы, H – максимальная высота тела (на уровне середины диска), HA – высота тела на уровне начала анального плавника, hpc – высота хвостового стебля, wc – ширина головы, W – максимальная ширина тела, io – межглазничное пространство, lmx – длина верхней челюсти; *данные только по внутреннему диску. Здесь и в табл. 3: M, m – среднее значение и его ошибка, min–max – пределы варьирования признака.
Самки и самцы достоверно различаются по трём признакам: расстоянию от ануса до основания A, антеанальному расстоянию и по максимальной высоте тела (табл. 2). Между самками и нерестовыми самцами выявлены достоверные различия по семи признакам: расстоянию между передним концом рыла и передним краем диска, антеанальному расстоянию, расстоянию между передним концом рыла и передним краем глаза, по высоте головы, высоте на уровне начала A, высоте хвостового стебля, длине maxillare. Самцы и нерестовые самцы достоверно различаются по восьми признакам, из которых пять общие с самками, – это расстояние между передним концом рыла и передним краем диска, высота головы, высота на уровне начала A, высота хвостового стебля, длина maxillare; три признака, отличающие нерестовых самцов от не вступивших в нерест, – расстояние от ануса до переднего конца A, длина P, максимальная высота тела.
Таблица 2.
Достоверность различий средних значений признаков самок, самцов и нерестовых самцов Eumicrotremus pacificus
| Признак | Самки–самцы | Самки–нерестовые самцы | Самцы–нерестовые самцы | |||
|---|---|---|---|---|---|---|
| t-value | p | t-value | p | t-value | p | |
| В % SL | ||||||
| a–disk | 1.685 | 0.100 | –3.093** | 0.006 | –6.593** | 0.000 |
| anus–A | 2.745** | 0.009 | –0.352 | 0.729 | 2.391* | 0.023 |
| aA | 3.768** | 0.001 | 2.324 | 0.031 | –1.300 | 0.203 |
| ao | –1.180 | 0.245 | –2.374* | 0.029 | –1.177 | 0.247 |
| o | –1.216 | 0.231 | –2.444* | 0.024 | –1.374 | 0.179 |
| lP | 1.042 | 0.304 | –1.887 | 0.075 | –2.755** | 0.009 |
| Hc | 0.685 | 0.497 | –2.832* | 0.011 | –2.867* | 0.007 |
| H | 2.154* | 0.038 | –1.368 | 0.187 | –2.776* | 0.009 |
| HA | 1.026 | 0.311 | –3.171** | 0.005 | –4.045** | 0.000 |
| hpc | –1.025 | 0.312 | –5.233** | 0.000 | –3.930** | 0.000 |
| lmx | –1.763 | 0.086 | –4.319** | 0.000 | –2.124* | 0.041 |
| В % c | ||||||
| o | –0.782 | 0.439 | –2.195* | 0.041 | –1.256 | 0.218 |
| po | 1.764 | 0.086 | 2.674* | 0.015 | 0.293 | 0.771 |
Рентгенограмма (объединённые данные, в круглых скобках – среднее, в квадратных скобках – данные лектотипа). D VI–VIII (6.9) [VII], I 8–I 10 (I 8.7) [I 8], A 9–11 (9.9) [10 ] , P 24–26 (24.7) [25 ] , C 1 + 5–4 + 4 + 1 [1 + 5 + 4 + 1], vert. 10–12 + + 16–17 = 26–28 (11.0 + 16.8 = 27.3) [10 + 17 = 27].
Эпиплевральных рёбер 4–11 пар, от 1–3–4-го позвонков. Первый луч D1 и 1-й луч A сверхштатные. У всех изученных особей лучи D1 гибкие колючие, не утолщены. Первый птеригиофор D1 подходит к невральному отростку 2-го позвонка. Между D1 и D2 два свободных птеригиофора. За исключением 1-го колючего луча остальные лучи D2 членистые, ветвистые. В хвостовом отделе 6–8 свободных невральных и 6–7 гемальных отростков. Верхний краевой луч C крепится на единственной эпуралии, 4–5 членистых ветвистых лучей – на эпаксиальной гипуральной пластинке, 4 луча – на гипаксиальной пластинке, нижний краевой луч – на гемальном отростке 1-го преурального центра.
Исследованные группы особей (самцы + самки, нерестовые самцы, лектотип) не различаются по числу плавниковых лучей и позвонков, а также по числу свободных от птеригиофоров невральных и гемальных отростков (табл. 3). У нерестовых самцов по сравнению с самками и не вступившими в нерест самцами лучи D1 несколько утолщены.
Таблица 3.
Счётные признаки по рентгенограммам самок и самцов, нерестовых самцов и лектотипа Eumicrotremus pacificus
| Признак | Самки и самцы (21 экз.) | Нерестовые самцы (4 экз.) | Лектотип | |||
|---|---|---|---|---|---|---|
| M | m | min–max | M | min–max | ||
| SL, мм | 40.3–87.0 | 52.7–63.2 | 62.6 | |||
| Число позвонков: | ||||||
| – туловищных | 11.0 | 0.32 | 10–12 | 10.0 | 10 | 10 |
| – общее | 27.3 | 0.64 | 26–28 | 27.0 | 27 | 27 |
| D1 | 6.9 | 0.50 | 6–8 | 7.0 | 7 | 7 |
| D2 (членистые) | 9.7 | 0.57 | 9–11 | 10.0 | 10 | 9 |
| A | 9.9 | 0.44 | 9–11 | 10.0 | 9–11 | 10 |
| C (основные) | 8.9 | 0.30 | 8–9 | 9.0 | 9 | 9 |
| Число свободных позвонков: | ||||||
| – от конца D2 | 6.9 | 0.44 | 6–8 | 6.7 | 6–7 | 7 |
| – от конца A | 6.5 | 0.60 | 6–7 | 6.7 | 6–7 | 6 |
Сейсмосенсорная система. По числу пор сейсмосенсорной системы на голове и по их форме существенная изменчивость не выявлена. Поры каналов и свободные невромасты расположены на коротких кожных трубочках. Имеются две поры CSO (1-я – перед передней ноздрей, 2-я – перед задней ноздрей); непарная пора межглазничной комиссуры (позади задних ноздрей); две поры CT (1-я – за глазом, 2-я – за верхним концом жаберного отверстия) и две поры CIO (под и за глазом). В CPM четыре–пять пор: передние две–три расположены на нижней челюсти, две задние – на praeoperculum: снизу и в нижнем заднем углу. Нет поры у верхнего конца praeoperculum. У личинок SL 15.3 и 15.7 мм межглазничная комиссура разорвана, формируя две поры.
Наружный скелет. Костные бляшки впервые отмечены у личинки SL 15.3 мм: верхняя и нижняя – в окологрудном ряду, нижняя – в зажаберном ряду. У личинки SL 15.7 мм есть задняя бляшка в надглазничном, передняя бляшка в заглазничном, четыре бляшки в окологрудном и одна в зажаберном ряду. У личинки SL 19.5 мм в срединном затылочном ряду отмечена одна бляшка, а у молоди SL 31.0 мм – две бляшки. У этой же особи имеются четыре бляшки в межглазничном ряду и три – в зажаберном. Максимальное число бляшек у взрослых особей в среднем затылочном ряду шесть, в межглазничном – семь, в надглазничном – шесть, в заглазничном – десять, в окологрудном – пять, в зажаберном – четыре. У самок степень развития костных бляшек на хвостовом стебле и на хвостовом отделе (без хвостового стебля) в среднем выше, чем у самцов, и существенно выше, чем у преднерестовых самцов (рис. 1, 2; табл. 4). Размеры бляшек у самок также несколько больше, чем у самцов, и существенно больше, чем у нерестовых самцов. У представителей обоих полов в окологрудном ряду бляшки крупные, уплощённые, граничат друг с другом за исключением окологрудного ряда у двух крупных нерестовых самцов, у которых мелкие бляшки с высокими изогнутыми назад вершинами расположены далеко друг от друга (рис. 2).
Рис. 1.
Фотография лектотипа Eumicrotremus pacificus Schmidt, 1904 – самец SL 62.6 мм, ЗИН № 12921. Ряды костных бляшек: cp – окологрудной, pb – зажаберный, po – посторбитальный, pro – предкрышечный, so – надглазничный.
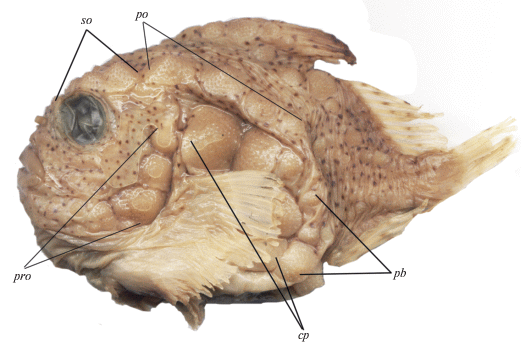
Рис. 2.
Рисунок нерестового самца Eumicrotremus pacificus – SL 55.2 мм, ННЦМБ № Ep6-11; обозначения см. на рис. 1.
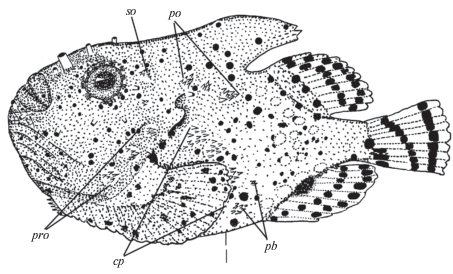
Таблица 4.
Распределение костных бляшек наружного скелета у самок, самцов и нерестовых самцов Eumicrotremus pacificus (доля особей), %
| Особь | n, экз. | SL, мм | Ряды костных бляшек | |||
|---|---|---|---|---|---|---|
| межглаз-ничные | в туловищном отделе | в хвостовом отделе (без хвостового стебля) | в хвостовом стебле | |||
| Самки | 15 | 39.0–80.7 | 100 | 100 | 100 | 67 |
| Самцы: | ||||||
| – не вступившие в нерест | 37 | 31.0–57.5 | 100 | 100 | 89 | 23 |
| – нерестовые | 8 | 52.7–63.2 | 0 | 100 | 43 | 0 |
Окраска в спирте. У всех личинок и ювенильных особей голова и тело равномерно тёмные; D1 тёмный, почти чёрный на конце; остальные плавники светлые. У взрослых особей окраска варьирует. Часть мелких особей SL ≤ 50 мм имеет светло-коричневую окраску головы, туловища и плавников, из которых тёмный лишь D1; у остальных на светло-коричневом туловище встречаются чёрные пятнышки от мелких до средних размеров; на D2, A и C также имеются более или менее выраженные чёрные пятнышки, приуроченные обычно к 1-му и 2-му лучу D2 и A и к верхним лучам C. У особей SL > 50 мм пятнышки на теле обычно хорошо выражены, наиболее крупные у особей SL > 60 мм. На плавниках пятнышки также крупнее и образуют по две продольные полосы вдоль плавниковых лучей D2 и A и две поперечные полосы на C. Наиболее заметны пятна на теле и непарных плавниках у нерестовых самцов.
ОБСУЖДЕНИЕ
Сравнительный анализ строения и окраски самок, самцов, нерестовых самцов и лектотипа E. pacificus выявил ряд различий между этими группами. Наиболее заметные различия наблюдаются по пластическим признакам и степени развития наружного скелета. Наибольшее число достоверных различий по пластическим признакам (11) установлено между нерестовыми самцами и самками и не достигшими нерестового состояния самцами. Очевидно, эти признаки характеризуют преобразования нерестовых самцов перед нерестом: удлинение верхней и нижней челюстей, увеличение высоты головы и тела. Самки и самцы E. pacificus также достоверно различаются по некоторым пластическим признакам, наиболее значимо – по величине антеанального расстояния, которое больше у самок. Причина этого различия, очевидно, связана с увеличением объёма полости тела в процессе развития гонад у самок.
Степень развития наружного скелета E. pacificus характеризуется некоторой изменчивостью в пределах каждой из рассмотренных групп, что соответствует представлениям других исследователей (Шмидт, 1904; Линдберг, Легеза, 1955; Ueno, 1970). Наибольшей степенью развития скелета отличаются самки, лишь у части которых (33%) костные бляшки отсутствуют на хвостовом стебле. Самцы отличаются от самок меньшей степенью развития костных бляшек на хвостовом стебле: они отсутствуют у 77% особей и в хвостовом отделе туловища (без хвостового стебля) – у 11% особей. Нерестовые самцы существенно отличаются от остальных представителей E. pacificus полным отсутствием костных бляшек на хвостовом стебле и в межглазничных рядах, заметным сокращением их числа в хвостовом отделе (без хвостового стебля) у 57% особей. Размеры костных бляшек у нерестовых самцов также заметно редуцированы, что особенно заметно у 2 экз. (ННЦМБ Ep6-11 и ЗИН № 303640). При этом у всех этих самцов сохраняется диагностический признак вида – четыре–пять костных бляшек в окологрудном ряду.
Нерестовые самцы E. pacificus характеризуются также максимальным развитием нерестовой окраски в виде чёрных пятнышек мелких и средних размеров на теле. На D2, A и C также имеются более или менее выраженные ряды чёрных пятнышек, обычно на 1-м и 2-м лучах D2 и A и верхних лучах C.
Таким образом, самки и не достигшие нерестового состояния самцы E. pacificus демонстрируют половой диморфизм по трём пластическим признакам, из которых наиболее заметным является величина антеанального расстояния, а также по большей степени развития наружного скелета в хвостовом отделе самок. Нерестовые самцы существенно отличаются не только от самок, но и от самцов, не участвующих в нересте.
Биркьедал с соавторами (Byrkjedal et al., 2007) на основании морфологического и молекулярного исследований E. eggvinii Koefoed, 1956 и E. spinosus (Fabricius, 1776) из Баренцева моря, Гренландии и банок Ньюфаундленда установили полное сходство двух видов по последовательностям изученных ядерного и двух митохондриальных (COI, COII) генов и лишь небольшие расхождения по митохондриальному гену цитохрома b. Вместе с тем морфологические различия между особями этих видов оказались значительными: установлены достоверные различия по 22 морфологическим признакам. В частности, у E. eggvinii достоверно большие, чем у E. spinosus, средние значения следующих признаков: относительная длина и ширина головы, ширина рта, длина рыла, длина основания D1 и А, длина основания Р, длина и ширина диска, расстояние от рыла до ануса. Биркьедал с соавторами (Byrkjedal et al., 2007) не использовали традиционный подход к оценке диагностических признаков наружного скелета, но они подсчитали общее число костных бляшек на левой стороне рыбы, что дало им возможность обнаружить значительное расхождение по этому признаку E. spinosus и E. eggvinii. На основании результатов молекулярного анализа авторы сделали вывод о том, что особи E. eggvinii представляют собой самцов E. spinosus, а значительные различия между самцами и самками E. spinosus по общему числу костных бляшек привели их к заключению о существенной изменчивости в степени развития наружного скелета и у остальных видов круглопёрых. В последующих фаунистических списках (Mecklenburg et al., 2011, 2018; Парин и др., 2014) E. eggvinii рассматривается как синоним E. spinosus. Недавно Воскобойникова и Чернова (2016) обнаружили в материалах фондовой коллекции ЗИН РАН самцов E. spinosus, сходных по строению внешнего скелета и ряду других морфологических признаков с самками этого вида. На этом основании был восстановлен видовой статус E. eggvinii. В настоящем исследовании между самками и нерестовыми самцами E. pacificus отмечены различия по длине maxillare и длине рыла, сходные с различиями между E. spinosus и E. eggvinii (Byrkjedal et al., 2007). Не достигшие нерестового состояния самцы и нерестовые самцы E. pacificus так же отличаются между собой по длине maxillare, как и E. spinosus и E. eggvinii. Кроме того, можно отметить сходство в редукции наружного скелета E. eggvinii и нерестовых самцов E. pacificus. В результате настоящего исследования морфологической изменчивости E. pacificus и сопоставления структуры полов E. spinosus и E. pacificus можно предположить, что E. eggvinii, возможно, являются нерестовыми самцами E. spinosus.
Таким образом, в результате изучения морфологической изменчивости E. pacificus установлены пределы изменчивости морфометрических признаков и строения наружного скелета в рамках одного вида круглопёров. Выявлен половой диморфизм E. pacificus. Показано, что самки характеризуются стабильностью строения наружного скелета на протяжении всего взрослого периода жизни. У нерестовых самцов происходит существенная редукция наружного скелета и изменение величин ряда пластических признаков, по-видимому, связанные с особенностями нереста видов Eumicrotremus (Воскобойникова и др., 2017). Можно предположить, что при внимательном изучении других видов рода будет обнаружена сходная дифференциация самок, самцов и нерестовых самцов по морфологическим признакам.
Список литературы
Воскобойникова О.С., Назаркин М.В. 2015. Переописание колючего круглопера Андрияшева Eumicrotremus andriashevi c выделением нового подвида Eumicrotremus andriashevi aculeatus ssp. nov. (Cyclopteridae) // Вопр. ихтиологии. Т. 55. № 2. С. 139–145.
Воскобойникова О.С., Чернова Н.В. 2016. Восстановление валидности круглопера Эггвина Eumicrotremus eggvini (Cyclopteridae) и его новая находка у Земли Франца-Иосифа (Баренцево море) // Там же. Т. 56. № 1. С. 34–40.
Воскобойникова О.С., Баланов А.А., Кудрявцева О.Ю. и др. 2017. Актуальные проблемы систематики круглоперых рыб семейства Cyclopteridae // Матер. юбилей. отчет. сессии, посвящ. 185-летию ЗИН РАН. СПБ.: Изд-во ЗИН РАН. С. 27–30.
Линдберг Г.У., Легеза М.И. 1955. Обзор родов и видов рыб подсемейства Cyclopterinae (Pisces) // Тр. ЗИН АН СССР. Т. 18. С. 389–458.
Парин Н.В., Евсеенко С.А., Васильева Е.Д. 2014. Рыбы морей России: аннотированный каталог. М.: Т-во науч. изд. КМК, 733 с.
Шмидт П.Ю. 1904. Рыбы восточных морей Российской империи. Научные результаты Корейско-Сахалинской экспедиции Императорского Русского Географического Общества. СПб.: Изд-во Императ. рус. геогр. о-ва, 466 с.
Byrkjedal I., Rees D.J., Willassen E. 2007. Lumping lumpsuckers: molecular and morphological insights into taxonomic status of Eumicrotremus spinosus (Fabricius, 1776) and Eumicrotremus eggvinii Koefoed, 1956 (Teleostei: Cyclopteridae) // J. Fish. Biol. V. 71. P. 111–131.
Kai Y., Stevenson D.E., Ueda Y. et al. 2015. Molecular insights into geographic and morphological variation within the Eumicrotremus asperrimus species complex (Cottoidei: Cyclopteridae) // Ichthyol. Res. V. 62. P. 396–408.
Lee S.J., Kim Y.-K., Moon D.Y.K., Kai J.-K. 2015. Taxonomic review of the Korean lumpsucker “Do-chi” reported previously as Eumicrotremus orbis (Pisces: Cyclopteridae) based on morphological and molecular characters // Fish. Aquat. Sci. V. 18. № 4. P. 405–410.
Lee S.J., Kim J.-K., Ikeguchi S., Nakabo T. 2017. Taxonomic review of dwarf species of Eumicrotremus (Actinopterrygii: Cottoidei: Cyclopteridae) with descriptions of two new species from the western North Pacific // Zootaxa. V. 4282. № 2. P. 337–349.
Mecklenburg C.W., Miller P.R., Steinke D. 2011. Biodiversity of the Arctic marine fishes: taxonomy and zoogeography // Mar. Biodiversity. V. 41. № 1. P. 109–140.
Mecklenburg C.W., Lynghammar A., Johannesen E. et al. 2018. Marine fishes of the Arctic Region. V. I. Conservation of Arctic Flora and Fauna. Akureyri, Iceland: CAFF, 454 p.
Stevenson D., Mecklenburg K.W., Kai Y. 2017. Taxonomic clarification of the Eumicrotremus asperrimus species complex (Teleostei: Cyclopteridae) in the eastern North Pacific // Zootaxa. V. 4294. № 4. P. 419–435.
Ueno T. 1970. Fauna Japonica: Cyclopteridae (Pisces). Tokyo: Acad. Press Jpn., 233 p.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии