

Вопросы ихтиологии, 2019, T. 59, № 6, стр. 672-680
Некоторые результаты изучения динамики численности горбуши Oncorhynchus gorbuscha на северо-восточном побережье острова Сахалин
1 Сахалинский филиал Всероссийского научно-исследовательского института рыбного хозяйства
и океанографии − СахНИРО
Южно-Сахалинск, Россия
2 Сахалинский государственный университет − СахГУ
Южно-Сахалинск, Россия
* E-mail: kaev@outlook.com
Поступила в редакцию 02.10.2018
После доработки 18.02.2019
Принята к публикации 04.03.2019
Аннотация
Проанализированы данные, характеризующие численность отдельных поколений горбуши Oncorhynchus gorbuscha (объём вылова, захода производителей в реки и последующего ската молоди) в северном и южном районах северо-восточного побережья Сахалина. Судя по соотношению численности производителей на нерестилищах и покатной молоди, эффективность воспроизводства в горных реках горбушёвого типа южной части побережья выше, чем в сравнительно крупных равнинных реках северной части. Показано, что наблюдавшийся в последние годы рост численности горбуши был обусловлен в основном увеличением выживаемости поколений в течение морского периода жизни.
Протяжённость северо-восточного побережья Сахалина − от м. Терпения на юге до м. Елизаветы (северная оконечность острова) − составляет ~600 км. Нерест горбуши Oncorhynchus gorbuscha зарегистрирован в 111 реках. В 57 из них площадь нерестилищ оценивается на уровне 6.02 млн м2; остальные реки, как правило, небольшие, местами заболоченные и не имеют существенного значения в воспроизводстве горбуши. При столь огромном нерестовом фонде ежегодные уловы горбуши в 1961–1990 гг. составляли в среднем 0.61 тыс. т, что существенно ниже, чем в расположенных южнее заливах Терпения и Анива и юго-восточном побережье острова, где в эти же годы вылавливали в среднем 2.57, 2.84 и 9.43 тыс. т при площади нерестилищ в реках соответственно 6.89, 1.67 и 1.49 млн м2 (Каев, 2011а). Столь малые уловы были связаны не только с низким уровнем воспроизводства горбуши, но и со слабым развитием инфраструктуры в северо-восточной части острова. Горбушу в те годы промышляли в основном в приустьевой акватории р. Тымь (зал. Ныйский), а в остальных районах лов иногда вели выездные бригады рыбаков. Малой промысловой значимостью северо-восточного побережья Сахалина было обусловлено и небольшое число публикаций по изучению воспроизводства местной горбуши (Чуриков, 1975; Шершнев, Жульков, 1979; Гриценко и др., 1982, 1987; Шершнев и др., 1985). Для научного сопровождения промысла, география которого в этом районе стала расширяться с конца 1990-х гг., были систематизированы имеющиеся данные путём унификации расчётов численности заходов производителей в реки всех лет наблюдений и последующего ската молоди, а также биологических показателей рыб для характеристики поколений (Kaev, Geraschenko, 2008). Однако в дальнейшем публикацию данных по показателям воспроизводства этого стада горбуши прекратили, так как их оценка зачастую стала носить экспертный характер из-за отсутствия данных по численности покатной молоди.
В настоящее время назрела необходимость обобщить результаты предыдущих наблюдений как некой основы для наступающего нового этапа, характеризующегося расширением исследований в условиях возросшей промысловой значимости данного района.
МАТЕРИАЛ И МЕТОДИКА
Межгодовая и сезонная динамика уловов представлена по данным официальной промысловой статистики (Сахалино-Курильское территориальное управление Росрыболовства). Промысел почти исключительно вели ставными неводами (пассивное орудие лова), поэтому изменения в величине уловов отражают динамику нерестовых подходов горбуши к побережью. Число выловленных рыб за тот или иной период промысла рассчитывали путём деления биомассы уловов на среднюю массу тела особей, определяемую по биологическим анализам рыб (данные СахНИРО). Для характеристики численности производителей на нерестилищах использованы данные бассейнового управления Сахалинрыбвод, полученные методом визуального учёта рыб при пеших обходах ряда рек (Шевляков и др., 2013). Данные проанализированы с 1985 г., когда началась ежегодная промысловая деятельность в южной части побережья. За исследуемый период ежегодно путем подсчёта производителей на отдельных нерестилищах с дальнейшей экстраполяцией на всю нерестовую площадь водоёма обследовали от 3 до 24 рек, в среднем по 12 рек. Особое место в этом списке занимает вторая по величине на Сахалине р. Тымь с протяжённостью основного русла 330 км и площадью водосбора 7850 км2 (Ресурсы …, 1964), имеющая 49 притоков 1-го порядка, соизмеримых по величине со многими реками побережья. Понятно, что о численности заходов горбуши в эту реку по результатам 17-летних наблюдений приходилось судить по эпизодическим обследованиям отдельных нерестилищ в верхней части основного русла и в 2–15 притоках разного порядка.
Следует отметить принципиальную разницу между реками южной и северной части побережья. В южной части реки в среднем короче (наиболее крупная из них Лангери с длиной основного русла 101 км) и имеют горный характер, стекая с простирающихся вдоль побережья Восточно-Сахалинских гор. Севернее 50°50′ с.ш. горы отступают в глубь острова. В этой части побережья долины рек обширны и заболочены, часть рек (в том числе все крупные) впадает в заливы лагунного типа. Численность горбуши оценивали раздельно для южной и северной части побережья (рис. 1). В южных реках горбушёвого типа сосредоточено 1626 тыс. м2 нерестилищ, в северных – 4396 тыс. м2 (Kaev, Geraschenko, 2008). Для расчёта суммарной величины заходов рыб в реки каждого из районов использован принцип реперных водоёмов, в соответствии с которым все реки подразделены на три группы. Первую группу составляют регулярно обследуемые реки, вторую – эпизодически обследуемые, третью – реки, в которых обследования не проводили. В первую группу вошли реки Мелкая (32 года наблюдений), Богатая (30 лет) и Даги (26 лет). Рассчитанная плотность скоплений рыб (отношение подсчитанного числа рыб в реке к площади нерестилищ) в реках первой группы принималась за действительную. В годы отсутствия данных по этим рекам использовали данные наблюдений в ближайших реках-аналогах. Для рек второй группы плотность скоплений рыб рассчитана как среднее значение между наблюдённой в данной реке и определённой для рек из первой группы. Суммарные результаты наблюдений в реках первой и второй группы экстраполировали на реки третьей группы (Kaev, Geraschenko, 2008).
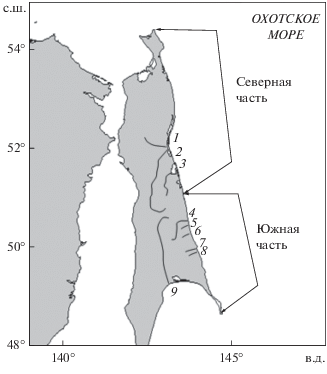
Рис. 1.
Схема разделения северо-восточного побережья Сахалина на северную и южную части, реки: 1 – Даги, 2 – Тымь, 3 – Набиль, 4 – Лангери, 5 – Малая Хузи, 6 – Хой, 7 – Богатая, 8 – Мелкая, 9 − Поронай.
Численность покатной молоди оценивали по данным её учёта в реках Мелкая и Даги, выполненного соответственно сотрудниками СахНИРО и Сахалинрыбвода методом выборочных обловов (Воловик, 1967; Каев, 2011б). В других реках объём ската рассчитывали путём умножения численности производителей в них на коэффициент ската (Кс − число покатников, приходящихся на одного условного производителя), установленный в контрольных реках. Показателем выживаемости горбуши в морской период служит доля (%) вернувшихся рыб от численности покатной молоди соответствующего поколения. Данные по численности рыб соответствуют данным, использованным при официальном представлении СахНИРО материалов для подготовки соответствующих статистических документов Северотихоокеанской комиссии по анадромным рыбам (North Pacific anadromous fish commission ̶ NPAFC: http://www.npafc.org/new/ pub_documents.html).
Биологическая характеристика рыб (длина по Смитту (FL), масса, абсолютная индивидуальная плодовитость (АИП)) основана на фондовых материалах СахНИРО, собранных при полевых наблюдениях в данном районе (17544 рыб в 197 пробах). При расчёте значений биологических показателей для поколения данным каждой пробы придавалась доля, соответствующая доле промыслового вылова рыб в периоды их сбора. Результаты представлены в виде среднего значения и среднего квадратичного отклонения (M ± SD), достоверность различия между средними значениями оценена по критерию Фишера (F). В тексте использованы следующие символы: r – коэффициент корреляции Пирсона, p – уровень значимости нуль-гипотезы, n – число поколений.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
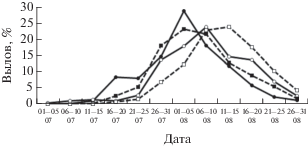
Уловы горбуши в 1990-х гг. в северной и южной части побережья мало различались, несмотря на начавшийся в дополнение к традиционным местам лова в районе впадения Тыми и соседних рек регулярный промысел в южных районах. Однако в начале XXI в., когда наметился существенный рост запасов горбуши в регионе, наиболее заметное увеличение уловов произошло в южной части побережья (рис. 2). В 2001–2018 гг. здесь стали добывать в нечётные годы в среднем 66%, а в чётные 78% суммарных уловов этого вида на северо-восточном побережье Сахалина. При этом чётко обозначился более высокий уровень уловов по нечётным годам, лишь в последние годы это соотношение нарушилось в результате резкого сокращения численности горбуши в 2017 г. на фоне роста её запасов в чётные годы. Судя по датам достижения половины уловов, в южной части побережья массовые подходы горбуши наблюдались несколько раньше, чем в северной, как по линии нечётных (3 августа против 6 августа), так и чётных лет (8 августа против 12 августа). Если сравнивать эти даты в каждом из районов по чётным и нечётным годам, то расхождение между датами увеличивается до 5 и 6 сут., т.е. хорошо заметно смещение подходов горбуши по чётным годам на более поздние даты (рис. 3).
Рис. 2.
Межгодовая динамика уловов горбуши Oncorhynchus gorbuscha в нечётные (⚫, ◼) и чётные (⚪, ◻) годы в южной (—) и северной (- -) части северо-восточного побережья Сахалина в 1990–2018 гг.
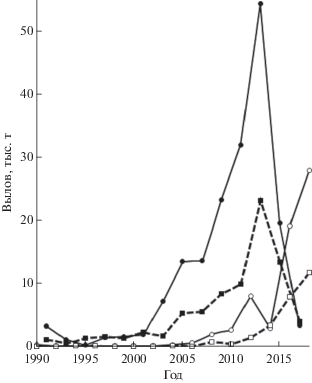
Рис. 3.
Сезонная динамика уловов горбуши Oncorhynchus gorbuscha в нечётные и чётные годы в южной и северной части северо-восточного побережья Сахалина, средние значения за период 2007–2018 гг.; обозначения см. на рис. 2.
В реки южной части побережья в 1985–2016 гг. ежегодно заходило от 15 до 8220 (в среднем 1993) тыс. производителей (табл. 1). После их нереста в соответствии со значениями Кс с нерестилищ мигрировало от 2.3 до 696.2 (170.6) млн мальков. Последующий возврат взрослых рыб составлял 29−43 718 (7786) тыс. экз. При промысле вылавливали от 0 до 40 007 (5967) тыс. рыб, т.е. коэффициент промыслового изъятия находился на уровне 46%, естественно, что его наибольшие значения (до 96%) приходились на годы с наибольшими подходами горбуши (исключая поколение 2015 г. рождения).
Таблица 1.
Численность поколений горбуши Oncorhynchus gorbuscha на разных этапах онтогенеза в южной и северной частях северо-восточного побережья Сахалина
| Год нереста | Южная часть | Северная часть | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Заход, тыс. экз. | Скат, млн экз. | Кс | Возврат | Заход, тыс. экз. | Скат, млн экз. | Кс | Возврат | |||
| тыс. экз. | % ската | тыс. экз. | % ската | |||||||
| 1985 | 8220 | 87.1 | 10.6 | 1662 | 1.91 | 3327 | 98.6 | 29.6 | 2675 | 2.71 |
| 1986 | 15 | 2.3 | 148.3 | 29 | 1.28 | 72 | 4.5 | 62.5 | 86 | 1.92 |
| 1987 | 1234 | 138.2 | 112.0 | 1644 | 1.19 | 1832 | 51.9 | 28.3 | 1947 | 3.75 |
| 1988 | 29 | 4.4 | 150.1 | 361 | 8.22 | 86 | 6.6 | 76.5 | 276 | 4.18 |
| 1989 | 1522 | 160.0 | 105.1 | 6400 | 4.00 | 1743 | 116.6 | 66.9 | 2919 | 2.50 |
| 1990 | 184 | 35.2 | 191.4 | 279 | 0.79 | 269 | 21.7 | 80.7 | 110 | 0.51 |
| 1991 | 3694 | 179.4 | 48.6 | 3230 | 1.80 | 1999 | 73.1 | − | 1232 | − |
| 1992 | 246 | 21.4 | 87.1 | 600 | 2.80 | 110 | 7.2 | − | 299 | − |
| 1993 | 2370 | 146.4 | 61.8 | 2752 | 1.88 | 833 | 38.8 | − | 3609 | − |
| 1994 | 536 | 11.6 | 21.6 | 510 | 4.40 | 251 | 4.1 | − | 291 | − |
| 1995 | 2600 | 143.5 | 55.2 | 5588 | 3.89 | 2683 | 111.5 | − | 5022 | − |
| 1996 | 443 | 13.8 | 31.2 | 845 | 6.12 | 257 | 17.8 | 69.3 | 290 | 1.63 |
| 1997 | 4600 | 76.9 | 16.7 | 2107 | 2.74 | 3881 | 71.7 | 18.5 | 3572 | 4.98 |
| 1998 | 797 | 48.4 | 60.7 | 377 | 0.78 | 285 | 22.6 | 79.2 | 278 | 1.23 |
| 1999 | 895 | 100.4 | 112.1 | 3824 | 3.81 | 2592 | 324.3 | 125.1 | 4341 | 1.34 |
| 2000 | 327 | 20.9 | 64.0 | 695 | 3.32 | 255 | 27.0 | 106.0 | 173 | 0.64 |
| 2001 | 2223 | 141.3 | 63.5 | 9226 | 6.53 | 2411 | 141.3 | 58.6 | 3323 | 2.35 |
| 2002 | 640 | 86.9 | − | 650 | − | 169 | 17.2 | 102.3 | 166 | 0.96 |
| 2003 | 3661 | 278.2 | − | 16162 | − | 2053 | 352.5 | 171.7 | 8486 | 2.39 |
| 2004 | 488 | 69.0 | − | 821 | − | 136 | 14.5 | 106.3 | 180 | 1.24 |
| 2005 | 3944 | 95.0 | − | 13315 | − | 3710 | 67.3 | 18.1 | 7249 | 10.77 |
| 2006 | 439 | 85.8 | − | 3286 | − | 152 | 22.4 | 147.5 | 578 | 2.50 |
| 2007 | 2933 | 343.7 | − | 19050 | − | 3009 | 265.4 | 88.2 | 8587 | 3.23 |
| 2008 | 1882 | 317.2 | − | 3714 | − | 133 | 16.9 | 126.9 | 392 | 2.32 |
| 2009 | 3194 | 170.7 | − | 28143 | − | 3074 | 123.8 | 40.3 | 10 300 | 8.18 |
| 2010 | 1845 | 429.0 | − | 8947 | − | 163 | 28.5 | 175.0 | 1412 | 4.96 |
| 2011 | 2354 | 191.2 | − | 43718 | − | 2354 | 143.9 | 61.1 | 19 223 | 13.25 |
| 2012 | 2388 | − | − | 3035 | − | 261 | − | − | 2684 | − |
| 2013 | 3712 | − | − | 17340 | − | 2211 | − | − | 11 481 | − |
| 2014 | 1119 | − | − | 18393 | − | 390 | − | − | 7160 | − |
| 2015 | 2700 | − | − | 2366 | − | 1848 | − | − | 2975 | − |
| 2016 | 2549 | 693.6 | 272.1 | 30070 | 4.34 | 584 | 119.7 | − | 13 153 | − |
В реки северной части побережья в эти же годы ежегодно заходило 72−3881 (1348) тыс. производителей. В соответствии со значениями Кс после их нереста скатывалось 4.1−352.5 (82.6) млн мальков. В дополнение к этому с двух лососёвых рыборазводных заводов (Адо-Тымовский и Тымовское, бассейн р. Тымь) в 1992, 2004, 2007, 2008, 2010, 2012 и 2014–2017 гг. выпускали соответственно 0.5, 2.9, 0.7, 0.4, 2.2, 1.1, 1.3, 0.4, 3.5 и 0.7 млн мальков. Ввиду малочисленности и нерегулярности выпусков эти данные не представлены в табл. 1, однако коэффициент возврата рассчитан от суммарного ската покатников дикого и заводского происхождения. Возврат взрослых рыб рассматриваемых поколений варьировал в пределах 86−19 223 (3890) тыс. экз. При промысле вылавливали от 0 до 17 011 (2589) тыс. рыб, коэффициент промыслового изъятия составлял в среднем 42% (максимально до 92%).
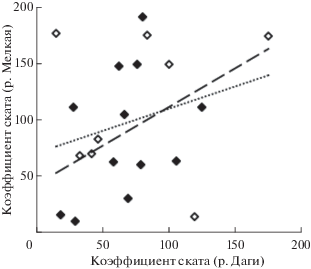
В табл. 1 для ряда поколений не рассчитаны значения коэффициентов ската и возврата, несмотря на наличие данных в прочих столбцах, что связано с отсутствием учёта покатной молоди этих поколений в данном районе. В таких случаях численность покатников рассчитывали на основе значений Кс, полученных по результатам учёта в контрольной реке другого района. Правомерность такого подхода обусловлена положительной корреляцией между значениями коэффициента в реках Даги и Мелкая (рис. 4). За все годы синхронных учётов молоди в этих реках (от нереста поколений в 1977–1990, 1996–2001 гг.) связь слабее, чем за период, начиная с поколения от нереста в 1985 г., что может быть связано с особенностями заполнения нерестилищ в р. Мелкая в условиях слабого промысла в ранние годы. За последний период (с 1985 г.) на одного условного производителя в р. Даги приходилось в среднем 67 покатников, а в р. Мелкая – 89 покатников, т.е. в 1.33 раза больше. В соответствии с этим для поколений горбуши от нереста в 1991–1995 гг. в северной части побережья при расчёте численности молоди применяли уменьшенное в 1.33 раза значение Кс, установленное для р. Мелкая, а для поколений горбуши от нереста в 2002–2011 гг. в южной части побережья – увеличенное в 1.33 раза значение Кс по р. Даги. Полученные таким образом значения численности покатной молоди отражают вероятные тенденции её изменений, но они не использованы для оценки коэффициентов ската и выживаемости поколений в морской период жизни вследствие возможных отклонений в расчётах (r = 0.38, p > 0.05). Для поколений от нереста в 2012–2015 гг. при экспертной оценке численности молоди в реках уже всего побережья использовали данные по эффективности нереста в бассейне р. Поронай (но эти данные не приведены в табл. 1 вследствие больших неопределённостей в расчётах). Для поколения от нереста 2016 г. расчёт проведён по результатам учёта покатников в р. Хой.
Рис. 4.
Соотношение значений коэффициента ската (Кс) при нересте горбуши Oncorhynchus gorbuscha в реках Мелкая и Даги в 1977–1984 (◇), 1985−1990 и 1996–2001 гг. (◆) и тренды: (···) – в целом весь период наблюдений, R2 = 0.0715; (− −) − 1985–2001 гг., R2 = = 0.1412.
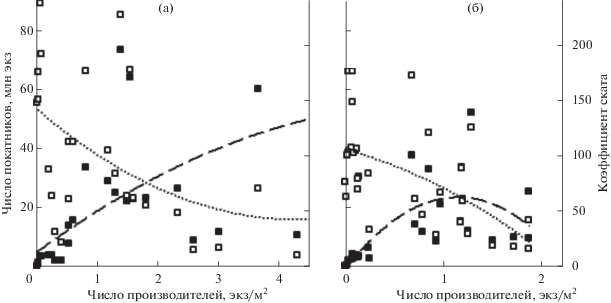
Воспроизводство горбуши в реках Мелкая и Даги различается не только числом покатников, приходящихся на одного условного производителя. На фоне общего для обеих рек снижения Кс с увеличением заходов горбуши в реки абсолютная численность покатников в р. Мелкая характеризовалась восходящим трендом (рис. 5а), в то время как в р. Даги при достижении плотности скоплений производителей на нерестилищах 1.2 экз/м2 наблюдалась смена направления тренда с увеличения на снижение (рис. 5б), в обоих случаях тренд представлен полиноминальной кривой 2-й степени.
Рис. 5.
Зависимость численности покатников (◼) и коэффициента ската (Кс) (◻) от плотности заполнения нерестилищ производителями горбуши Oncorhynchus gorbuscha в реках: а – Мелкая, б – Даги; (− −) – тренд численности покатников, (···) – тренд значений Кс.
Пробы для биологического анализа горбуши в 1979–2002 гг. отбирали из исследовательских уловов в р. Мелкая. Помимо этого с 1997 г. начат их сбор из промысловых уловов ставных неводов, сначала (по 2003 г.) в зал. Ныйский, в который впадают реки Тымь и Даги, затем добавились сборы в южной части побережья, которые с 2007 г. стали доминировать по числу проб, а с 2011 г. их сбор осуществлялся уже только в пределах южной части побережья, заметно доминировавшей по уловам. Естественно, возникает вопрос о правомочности использования проб, собранных в пределах одного из участков, для биологической характеристики горбуши в целом на побережье. Сборы из ставных неводов в 2009 г. позволяют сопоставить длину самцов и самок на разных участках побережья (рис. 6). При попарном сравнении проб в каждой из серий (по 5-дневкам) статистически значимые различия по длине рыб одинакового пола выявлены только в 6 из 24 сравниваемых пар (табл. 2). Эти различия могут быть связаны с ходом разных темпоральных группировок, описанных ранее применительно к восточному побережью Сахалина как япономорская и две охотоморские − ранняя и самая малочисленная поздняя (Гриценко, 1981). Позже было показано, что основу промысловых подходов горбуши к юго-восточной части Сахалина и к Итурупу составляют рыбы ранней (принимаемой ранее за япономорскую) и более многочисленной поздней темпоральной формы, мигрирующие из Тихого океана в Охотское море через проливы Курильской гряды. Смена в подходах этих форм тестируется, в частности, укрупнением рыб в уловах, особенно самцов, которые становятся крупнее самок (Каев, 2012). Судя по динамике уловов и размерному составу рыб, ход этих форм характерен и для более северных районов Сахалина, в том числе и на его северо-восточном побережье (Каев, Сидоренко, 2015). Наличие в этом районе разных темпоральных форм горбуши, исходя из изменений размеров тела, соотношения полов и степени развития брачного наряда в течение нерестового хода в 2014–2016 гг., предполагается в её заходах в реки Лангери и Малая Хузи (Кириллова и др., 2018).
Рис. 6.
Динамика уловов и длины (FL) самцов и самок горбуши Oncorhynchus gorbuscha в южной и северной частях северо-восточного побережья Сахалина в июле−августе 2009 г.:, уловы в южной (—) и северной (- - -) частях, FL самцов в южной (△) и северной (▲) частях, (⚪), (⚫) – то же самок.
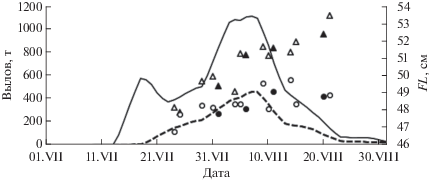
Таблица 2.
Значения критерия Фишера при попарном сравнении длины самцов и самок горбуши Oncorhynchus gorbuscha, отобранных из промысловых уловов ставных неводов в зал. Ныйский и в районах впадения рек Лангери и Мелкая в 2009 г.
| Дата сбора | Сравниваемые пробы | |||||
|---|---|---|---|---|---|---|
| Мелкая–Лангери | Мелкая–Ныйский | Лангери–Ныйский | ||||
| Cамцы | Cамки | Cамцы | Cамки | Cамцы | Cамки | |
| 24.VII | 0.2 | 7.9** | − | − | − | − |
| 29.VII–01.VIII | 0.2 | 0.1 | 0.4 | 1.9 | 1.0 | 0.8 |
| 04–06.VIII | 11.2** | < 0.1 | 10.8** | 0.9 | < 0.1 | 0.8 |
| 09–11.VIII | 0.3 | 11.7*** | < 0.1 | 2.0 | 0.2 | 7.7** |
| 14–15.VIII | 0.3 | 5.2* | − | − | − | − |
| 20.VIII | − | − | − | − | 2.1 | 0.1 |
В наших сборах самцы практически во всех случаях были крупнее самок (рис. 6), за исключением пробы в районе р. Лангери (24 июля), что было связано, видимо, со сменой в это время в подходах ранней темпоральной формы на позднюю. К концу хода половой диморфизм по длине тела становится более выраженным, поэтому появление статистически значимых различий в период 9−15 августа могло быть связано с разным темпом нарастания таких различий на разных участках побережья. Единственным исключением из этой схемы являются неожиданно сравнительно мелкие самцы в пробе, взятой 4 августа в районе р. Мелкая, что и обусловило их статистически значимые отличия от самцов из других районов.
Таким образом, в большинстве случаев не выявлены статистически значимые различия длины рыб в пробах, синхронно собранных в разных районах, а обнаруженные немногочисленные такие различия находят своё объяснение с точки зрения последовательного подхода рыб разных темпоральных форм. Пробу за 4 августа в районе р. Мелкая можно рассматривать как исключение, которое изредка отмечается и в других районах и может быть связано, например, с доминированием в выборке особей какой-то отдельной стаи, т.е. с проявлением эффекта элементарных популяций (Лебедев, 1967). Эти результаты дают основания представлять пробы, собранные в той или иной части побережья, в едином массиве данных.
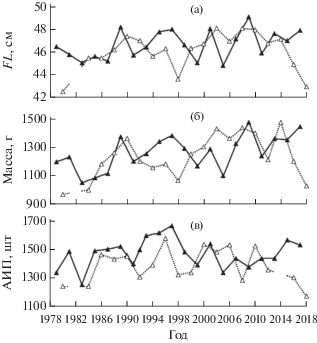
В нечётные годы длина рыб в среднем равна 46.7 ± 1.24 см, в чётные – 46.1 ± 1 .61 см, масса соответственно – 1262 ± 120 и 1235 ± 151 г, АИП – 1469 ± 102 и 1386 ± 118 икринок. В целом в чётные годы рыбы были несколько мельче по длине (F = = 1.6, p > 0.05) и массе тела (F = 0.4, p > 0.05), и именно в чётные годы трижды наблюдалось снижение размеров рыб до минимальных значений (рис. 7а, 7б). В отличие от длины и массы АИП в чётные годы была ниже статистически значимо (F = 5.4, p < 0.05). Считается, что плодовитость самок горбуши тесно связана с длиной тела, приближаясь у этого вида с наиболее упрощённой среди лососей возрастной структурой к прямолинейной зависимости (Костарев, 1965; Каев, 2003). Однако в данном случае корреляция плодовитости самок с их длиной (r = 0.56) и массой (r = 0.59) существенно слабее, чем между длиной и массой тела (r = 0.88). Разнонаправленность изменений в некоторые годы хорошо просматривается при сопоставлении динамики этих показателей по генеративным линиям нечётных и чётных лет (рис. 7в). Сравнение плодовитости самок в данном случае с размерами тела всех рыб вполне уместно, учитывая тесную связь длины (r = 0.95) и массы тела (r = 0.96) самок с суммарными значениями этих показателей для рыб обоих полов. Малые размеры горбуши и плодовитости самок в 2018 г., исходя из представлений о плотностно-зависимой регуляции роста (Гриценко и др., 1983; Карпенко и др., 2013), можно трактовать как следствие рекордно высокой численности рыб этого вида по линии чётных лет на северо-восточном побережье Сахалина и в целом в Охотском море (по предварительным данным, вылов ~400 тыс. т), учитывая смешение особей разных стад в районах совместного нагула сеголеток в Охотском море и зимовки в Тихом океане (Атлас …, 2002; Шунтов, Темных, 2008, 2011). Однако в 1980 и 1998 гг. малые размеры горбуши наблюдались при её сравнительно низкой численности на северо-восточном побережье Сахалина (Kaev, Geraschenko, 2008) и небольших уловах в бассейне Охотского моря (российский вылов соответственно − 73 и 179 тыс. т). Напротив, самая крупная горбуша на побережье отмечена в 2009 г. на фоне местного роста её численности (табл. 1) и больших уловов в Охотском море (Irvine et al., 2012). То есть совокупность данных по численности и размерам горбуши на северо-восточном побережье Сахалина не в полной мере соответствует концепции плотностно-зависимой регуляции её роста.
Рис. 7.
Динамика биологических показателей горбуши Oncorhynchus gorbuscha у северо-восточного побережья Сахалина в нечётные (–▲–) и чётные (··△··) годы в период 1979–2018 гг.: а − длина (FL), б – масса, в − абсолютная индивидуальная плодовитость (АИП).
Таким образом, на северо-восточном побережье Сахалина реки, протекающие в его северной и южной части, различаются по уровню и эффективности воспроизводства горбуши, что хорошо тестируется как величиной уловов по отношению к площади нерестилищ, так и числом покатной молоди, приходящейся на одного условного производителя. Кроме того, в северных реках при плотности скоплений производителей на нерестилищах выше 1.2 экз/м2 (соответствует суммарному заходу во все реки 5.3 млн рыб) отмечается снижение численности покатников (рис. 5б), что является одним из важнейших признаков превышения оптимальной численности производителей на нерестилищах (Гриценко и др., 1987). В южных реках такой перегиб кривой не наблюдается (рис. 5а), хотя значение Кс при росте численности производителей также снижается. Так, при плотности производителей >2.5 экз/м2 на одно сравнительно высокое значение численности покатников приходится три крайне низких их значения, исходя из чего 4.1 млн рыб (суммарный заход рыб в реки при данной плотности) можно принять за оптимальную величину пропуска производителей в реки южной части побережья.
В зимний период северная часть Сахалина, особенно по долинам рек в центральной части и по охотоморскому побережью, характеризуется суровым климатом. Так, в Тымь-Поронайской низменности сумма отрицательных значений температуры достигает 2600–2700°С, в то время как в Сусунайской низменности (южные районы острова) – 1350–1450°С (Атлас …, 1967). Тем не менее, коэффициент ската для поколений 1985–2001 гг. рождения в р. Мелкая (в среднем 79 экз.) находился в пределах варьирования данного показателя в реках южного и юго-восточного побережья Сахалина, таких как Быстрая (70), Кура (73), Дудинка (145) и Бахура (164 экз.). Вследствие более ранних сроков нерестового хода горбуши в реках северной части Сахалина её нерест совпадает по срокам с летней меженью, что в значительной мере в дальнейшем предохраняет нерестовые гнёзда горбуши от промерзания, так как при зимнем падении уровня подавляющее их большинство остаётся под водой (Гриценко и др., 1987). Следовательно, в среднем на треть более низкие значения Кс в одни и те же годы в р. Даги в сравнении с р. Мелкая вполне можно рассматривать как последствия нереста в равнинных северных реках, менее пригодных для воспроизводства горбуши.
По наблюдённым данным, выживаемость в течение морского периода жизни для поколений горбуши 1985–1990, 1996–2001 гг. рождения из рек северной части побережья также была меньше, чем из рек южной части (в среднем 2.31 против 3.39%). Это существенно ниже, чем для поколений этих же лет рождения, например, у горбуши юго-восточного побережья Сахалина и о-ва Итуруп − соответственно 4.96 и 4.93% (Kaev, Irvine, 2016). Различия в выживаемости связаны, видимо, с неблагоприятными условиями обитания молоди в прибрежных водах северных районов острова вследствие холодного Восточно-Сахалинского течения как элемента крупномасштабной циклонической циркуляции вод в Охотском море, которое движется в южном направлении вдоль восточного побережья Сахалина с последующим выносом охотоморских вод через проливы южной части Курильской гряды (Чернявский, 1981). Так, в годы ската молоди рассматриваемых поколений температура воды в прибрежье в июне у северо-восточного побережья острова составляла 0.7−7.1 (3.2)°С, а у юго-восточного побережья – 6.3−9.4 (8.0)°С (Kaev, Geraschenko, 2008). По современным данным, интенсивность Восточно-Сахалинского течения летом сильно ослабевает вплоть до того, что южнее м. Терпения оно размывается и далее к югу не прослеживается (Пищальник и др., 2003). Возможно, поэтому выживаемость поколений горбуши южной части северо-восточного побережья Сахалина выше, чем северной части этого побережья.
При рассмотрении изменений выживаемости поколений горбуши в периоды её низкой и высокой численности (до и после 2003 г.) можно оперировать только наблюдёнными данными для поколений нечётных лет 1985–1989, 1997–2001 и 2003–2011 гг. рождения в северной части побережья, так как в южной части реальный скат молоди для периода высокой численности не известен. По чётным годам рост численности начался позже (рис. 2), вследствие чего реальный скат молоди для этого периода известен только для одного поколения в северной (2010 г. рождения) и одного (2016 г. рождения) в южной части побережья (табл. 1), что явно недостаточно для анализа. Из имеющихся данных по первой группе поколений следует, что в период высокой численности от нереста одного условного производителя скатывалось несколько больше молоди, чем в период низкой численности (соответственно: 191 ± 116, n = 5; 179 ± 98, n = 6; F = 0.03, p > 0.05). В то же время существенно различалась выживаемость этих поколений в течение морского периода жизни (соответственно: 7.56 ± 4.71, n = 5; 2.89 ± 1.26, n = 6; F = 5.55, p < 0.05). Данный результат, полученный на небольшом числе проанализированных поколений, соответствует таковому, установленному при анализе многолетних рядов данных, в соответствии с которым наиболее значимые тенденции изменения численности горбуши юго-восточного побережья Сахалина, зал. Анива и о-ва Итуруп определялись выживаемостью поколений преимущественно в ранний морской период жизни (Kaev, Irvine, 2016). Существенному повышению выживаемости молоди могло способствовать потепление прибрежных вод. Согласно базе спутниковых данных, используемой в СахНИРО для изучения поверхностной температуры Охотского моря (Цхай, Шевченко, 2013), на контрольном полигоне у северо-восточного побережья Сахалина (средние координаты 50.5° с.ш. и 144.5° в.д.) температура воды в июле−августе при нагуле молоди поколений нечётных 1997–2001 гг. рождения составляла в среднем 8.7°С, а 2003–2011 гг. рождения – 10.4°С.
Полученные результаты показали, что наряду с общими чертами изменения численности горбуши в регионе (рост запасов в XXI в. преимущественно за счёт увеличения выживаемости в течение морского периода жизни) динамике стада этого вида на северо-восточном побережье Сахалина присущи свои особенности. В частности, они связаны с разной эффективностью нереста в крупных равнинных реках северной части побережья и в горных реках горбушёвого типа в южной части побережья. Сравнительно низкая выживаемость горбуши северо-восточного побережья Сахалина в течение морского периода жизни в первую очередь обусловлена неблагоприятным температурным режимом прибрежных морских вод, так как последующий нагул сеголеток в центрально-южной части Охотского моря и зимовка в океанских водах протекают в районах совместного обитания с горбушей других стад региона.
Список литературы
Атлас распространения в море различных стад тихоокеанских лососей в период весенне-летнего нагула и преднерестовых миграций. 2002 / Под ред. Гриценко О.Ф. М.: Изд-во ВНИРО, 190 с.
Атлас Сахалинской области. 1967. М.: Изд-во ГУ геодезии и картографии при Совете Министров СССР, 135 с.
Воловик С.П. 1967. Методы учета и некоторые особенности поведения покатной молоди горбуши в реках Сахалина // Изв. ТИНРО. Т. 61. С. 104–117.
Гриценко О.Ф. 1981. О популяционной структуре горбуши Oncorhynchus gorbuscha (Walbaum) // Вопр. ихтиологии. Т. 21. Вып. 5. С. 787–799.
Гриценко О.Ф., Ковтун А.А., Косткин В.К. 1982. Особенности формирования численности поколений горбуши р. Тымь // Рыб. хоз-во. № 4. С. 33–35.
Гриценко О.Ф., Ковтун А.А., Косткин В.К. 1983. Долгопериодные изменения размеров горбуши // Там же. № 6. С. 30–31.
Гриценко О.Ф., Ковтун А.А., Косткин В.К. 1987. Экология и воспроизводство кеты и горбуши. М.: Агропромиздат, 166 с.
Каев А.М. 2003. Особенности воспроизводства кеты в связи с ее размерно-возрастной структурой. Южно-Сахалинск: Изд-во СахНИРО, 288 с.
Каев А.М. 2011а. Оценка эффективности прогнозирования и управления промыслом горбуши в Сахалино-Курильском регионе // Изв. ТИНРО. Т. 167. С. 32–53.
Каев А.М. 2011б. Методическое руководство по количественному учету покатной молоди горбуши и кеты в малых реках методом выборочных обловов. Южно-Сахалинск: Изд-во СахНИРО, 16 с.
Каев А.М. 2012. Темпоральная структура и некоторые вопросы динамики стада горбуши Oncorhynchus gorbuscha (Salmonidae) // Вопр. ихтиологии. Т. 52. № 1. С. 62–71.
Каев А.М., Сидоренко М.Е. 2015. Прогноз и фактическое развитие промысла горбуши в 2015 г. в основных районах ее воспроизводства в Сахалинской области // Изучение тихоокеанских лососей на Дальнем Востоке. Бюл. № 10. С. 35–40.
Карпенко В.И., Андриевская Л.Д., Коваль М.В. 2013. Питание и особенности роста тихоокеанских лососей в морских водах. Петропавловск-Камчатский: Изд-во КамчатНИРО, 304 с.
Кириллова Е.А., Кириллов П.И., Кузищин К.В. и др. 2018. Морфобиологическая разнокачественность и особенности воспроизводства горбуши Oncorhynchus gorbuscha из двух рек северо-востока острова Сахалин // Вопр. ихтиологии. Т. 58. № 6. С. 670–683.
Костарев В.Л. 1965. Плодовитость охотской горбуши // Изв. ТИНРО. Т. 59. С. 145–155.
Лебедев Н.В. 1967. Элементарные популяции рыб. М.: Пищ. пром-сть, 212 с.
Пищальник В.М., Архипкин В.С., Юрасов Г.И., Ермоленко С.С. 2003. Сезонные вариации циркуляции вод в прибрежных районах Сахалина // Метеорология и гидрология. № 5. С. 87–95.
Ресурсы поверхностных вод СССР. Гидрологическая изученность. 1964. Т. 18. Дальний Восток. Вып. 3. Сахалин и Курилы. Л.: Гидрометеоиздат, 124 с.
Цхай Ж.Р., Шевченко Г.В. 2013. Оценка температурных аномалий поверхности Охотского моря и прилегающих акваторий по спутниковым данным // Исследования Земли из космоса. № 2. С. 50–61.
Чернявский В.И. 1981. Циркуляционные системы Охотского моря // Изв. ТИНРО. Т. 105. С. 13–19.
Чуриков А.А. 1975. Особенности ската молоди лососей рода Oncorhynchus из рек северо-восточного побережья о-ва Сахалин // Вопр. ихтиологии. Т. 15. Вып. 6. С. 1078–1085.
Шевляков Е.А., Шубкин С.В., Дубынин В.А. и др. 2013. Методики учета производителей тихоокеанских лососей на нерестилищах и путях миграции к ним // Изучение тихоокеанских лососей на Дальнем Востоке. Бюл. № 8. С. 36–57.
Шершнев А.П., Жульков А.И. 1979. Особенности ската молоди и некоторые показатели эффективности воспроизводства горбуши Oncorhynchus gorbuscha (Walbaum) в р. Приторной // Вопр. ихтиологии. Т. 19. Вып. 1. С. 128–133.
Шершнев А.П., Руднев В.А., Белобржеский В.А. 1985. Некоторые особенности нерестовой миграции горбуши Oncorhynchus gorbuscha (Walbaum, Salmonidae) северо-восточного побережья Сахалина // Там же. Т. 25. Вып. 6. С. 951–956.
Шунтов В.П., Темных О.С. 2008. Тихоокеанские лососи в морских и океанических экосистемах. Т. 1. Владивосток: Изд-во ТИНРО-центра, 481 с.
Шунтов В.П., Темных О.С. 2011. Тихоокеанские лососи в морских и океанических экосистемах. Т. 2. Владивосток: Изд-во ТИНРО-центра, 473 с.
Irvine J.R., Tompkins A., Saito T. et al. 2012. Pacific salmon status and abundance trends − 2012 update // NPAFC Doc. № 1422. Rev. 2. 90 p.
Kaev A.M., Geraschenko G.V. 2008. Reproduction indices of the north-eastern Sakhalin pink salmon // Ibid. № 1124. 11 p.
Kaev A.M., Irvine J.R. 2016. Population dynamics of pink salmon in the Sakhalin-Kuril region, Russia // Bull. NPAFC. № 6. P. 297–305.
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии