

Вопросы ихтиологии, 2019, T. 59, № 6, стр. 724-731
Активный выход молоди горбуши Oncorhynchus gorbusha (Salmonidae) в поток для пассивной покатной миграции
Д. С. Павлов 1, Е. А. Кириллова 1, *, П. И. Кириллов 1
1 Институт проблем экологии и эволюции РАН − ИПЭЭ
Москва, Россия
* E-mail: ekirillova@sevin.ru
Поступила в редакцию 13.06.2019
После доработки 13.06.2019
Принята к публикации 13.06.2019
Аннотация
По результатам полевых экспериментов и натурных наблюдений показано, что пассивной покатной миграции молоди горбуши Oncorhynchus gorbuscha предшествует её активный выход в речной поток. Он определяется врождёнными поведенческими реакциями (реореакция, оптомоторная реакция, фото- и тигмореакция) в ответ на изменения внешней среды.
Покатная миграция – первый и неотъемлемый этап миграционного цикла у большинства рыб, места нереста и нагула которых разобщены в пространстве. Многие лососёвые рыбы (Salmonidae) нерестятся в реках, а их молодь мигрирует в нагульный водоём (море или озеро). У анадромных видов длительность пресноводного периода жизни видоспецифична: может составлять от нескольких суток до нескольких лет (Гриценко, 2002; Quinn, 2005; Шунтов, Темных, 2008; Павлов и др., 2010, 2011). В зависимости от способа перемещения различают три формы покатной миграции – пассивную, активную и активно-пассивную. В первом случае перемещение представляет собой снос рыб потоком в не ориентированном относительно направления течения положении; во втором – рыбы движутся вниз по течению, затрачивая энергию для достижения мест нагула; в третьем – рыбы ориентированы головой против потока и, слабо сопротивляясь ему, сносятся вниз по течению (Павлов, 1979; Павлов и др., 2007). У лососёвых пассивная и активно-пассивная формы покатной миграции свойственны ранней молоди, как правило, ещё не перешедшей или переходящей на экзогенное питание. Для более поздней молоди первого года жизни, задерживающейся в реке и перешедшей на экзогенное питание, а также для молоди видов с длительным пресноводным периодом жизни свойственны активная и активно-пассивная формы миграции (Павлов и др., 2010, 2011).
Возможность реализации покатной миграции и её видоспецифические закономерности определяются комплексом врождённых поведенческих реакций, среди которых особое место принадлежит положительной реореакции – врождённому движению рыб против течения (Павлов и др., 2007). Эта реакция имеет две составляющие – ориентационную и локомоторную. Отсутствие условий для проявления одной из них (ухудшение условий для ориентации, снижение локомоторной активности рыб или их попадание в зоны с критическими скоростями течения) приводит к “нейтрализации” реореакции и сносу рыб потоком (Павлов, 1979; Павлов и др., 2007).
Закономерностям покатной миграции молоди лососёвых посвящено много публикаций. Отметим несколько обобщающих результаты разных исследований работ: Грут и Марголис (Groot, Margolis, 1991), Карпенко (1998), Гриценко (2002), Куинн (Quinn, 2005), Шунтов и Темных (2008), Павлов с соавторами (2010, 2011). Среди многих видов лососёвых особый интерес представляет горбуша Oncorhynchus gorbusha – вид, имеющий самый короткий пресноводный период жизни. Известно, что молодь горбуши начинает миграцию в море вскоре после выхода из нерестовых гнёзд и достигает нагульных акваторий в основном благодаря пассивной покатной миграции (Pritchard, 1944; Neave, 1955; Павлов и др., 2010). Миграция имеет межрегиональные различия в сроках и динамике, которые определяются локальными особенностями условий среды – термическим и гидрологическим режимом водотока, длительностью светлого и тёмного периода суток (McDonald, 1960; Groot, Margolis, 1991; Карпенко, 1998; Гриценко, 2002; Quinn, 2005; Шунтов, Темных, 2008; Павлов и др. 2010). Как правило, миграция приурочена к тёмному периоду суток. Исключение составляет часть ареала горбуши, где миграция молоди проходит в условиях полярного дня (Бакштанский, 1970, 1977; Камышная, 1974). До начала покатной миграции ранняя молодь горбуши находится в грунте (Солдатов, 1912; Pritchard, 1944; Hoar, 1951; Neave, 1955; Hoar et al., 1957; Бакштанский, 1970; Godin, 1980), и остаётся неясным, каким образом она оказывается в потоке воды для совершения покатной миграции. Нам не удалось найти специальных исследований этого вопроса.
Ранее на молоди карповых (Cyprinidae) и окунёвых (Percidae) рыб было выявлено, что их личинки активно выходят в поток в период вечерних сумерек и совершают покатную миграцию (Павлов и др., 1981; Павлов и др., 2007). В связи с этим было выдвинуто предположение о том, что и у молоди горбуши выход в поток происходит активно.
Цель настоящего исследования – в условиях эксперимента выявить поведенческий механизм выхода молоди горбуши в поток для реализации пассивной покатной миграции.
МАТЕРИАЛ И МЕТОДИКА
Экспериментальные исследования и полевые наблюдения проводили в период массового ската горбуши в р. Малая Хузи, во II−III декаде июня 2016–2017 гг. Река Малая Хузи – типичный малый водоток горного типа длиной 32 км на северо-востоке Сахалина. Подробное описание процедуры отлова покатной молоди горбуши и результаты исследования закономерностей её покатной миграции представлены в наших предыдущих работах на этой реке (Павлов и др., 2015; Кириллова, Кириллов, 2017; Кириллов и др., 2018). Эксперименты дополнялись круглосуточными визуальными наблюдениями за поведением молоди рыб в прибрежье реки. Все работы сопровождались фото- и видеорегистрацией. В тёмное время суток использовали прибор ночного видения или кратковременную подсветку фонарём с рассеянным световым потоком.
Экспериментальная установка для изучения характера перемещений молоди горбуши и её отношения к течению является модификацией двухсекционной щелевой камеры (Pavlov et al., 1997). Установка (50 × 70 × 10 см) состоит из двух равных по площади секций (непроточной и проточной), имитирующих непроточное прибрежье и транзитный поток в естественных условиях; секции разделены сплошной перегородкой с отверстием шириной 3 см посередине; поверхность дна ровная гладкая (рис. 1).
Рис. 1.
Схема экспериментальной установки: 1 – непроточная (стартовая) секция, 2 – проточная секция, 3 – перегородка, 4 – отверстие, 5 – решётка, обеспечивающая приток и отток воды в проточной секции установки, (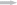 ) – направление и относительная скорость течения.
) – направление и относительная скорость течения.
Установку размещали непосредственно в реке, вблизи берега. Проточная секция располагалась дальше от берега, скорость течения в ней была 0.15−0.30 м/с; у разделяющей секции перегородки скорость течения была ниже, чем у противоположной стенки. Выбранные нами скорости течения согласуются с данными Хора (Hoar, 1951), который показал, что при скорости течения 0.03–0.45 м/с ранняя молодь горбуши демонстрирует устойчивую положительную реореакцию.
Опытных рыб до экспериментов содержали в течение 1.5 сут. в слабопроточных садках с размещёнными на дне камнями в качестве укрытий. Часть рыб во время адаптации находилась под камнями, часть – в потоке воды. Не менее чем за 20 мин до начала эксперимента рыб перемещали из садка в непроточную (стартовую) секцию установки (при закрытом отверстии в перегородке) для адаптации. Во время помещения рыб в установку освещённость составляла 50–100 лк. Отверстие открывали при освещённости 5–10 лк. Длительность каждого эксперимента составляла 7–8 ч, т.е. весь сумеречно-ночной период, от ранних вечерних сумерек до полного рассвета. В утренние часы при освещённости 1–5 лк отверстие в перегородке между секциями закрывали. Ежечасно регистрировали число рыб в секциях установки.
Длина по Смитту (FL) покатной молоди горбуши варьировала в пределах 28−35 (в среднем 31.6) мм, масса – 108−260 (199.2) мг. Всего проведено 12 экспериментов (по 20 рыб в каждом) с 240 рыбами ещё не перешедшими на экзогенное питание. Уровень освещённости с точностью до десятых долей люкса определяли при помощи люксметра PCE-174. Порядки освещённости в сотые и тысячные доли люкса оценивали с использованием справочных материалов (Кухлинг, 1985; Schlyter, Sweden, 2006). После завершения эксперимента всех рыб отлавливали и анестезировали с помощью гвоздичного масла (6 мл/л), действующего аналогично MS-222 (Keene et al., 1998; Руководство …, 2011). Затем рыб фиксировали в 4%-ном растворе формальдегида для последующего измерения длины и массы.
РЕЗУЛЬТАТЫ
До открытия выхода в проточную секцию молодь горбуши равномерно распределялась вдоль дальних стенок непроточной секции на расстоянии не более 5 см от них, избегая центральной наиболее открытой части и пространства у перегородки между секциями (рис. 2). Рыбы держались в толще воды в произвольном друг относительно друга положении. Потревоженные, например резким приближением наблюдателя, они объединялись в плотную стаю.
Рис. 2.
Распределение молоди горбуши Oncorhynchus gorbuscha в экспериментальной установке при разной освещённости, лк: а − > 0.5, б – 0.5−0.1,
в − < 0.1; (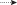 ) – направление перемещений рыб в установке; ост. обозначения см. на рис. 1.
) – направление перемещений рыб в установке; ост. обозначения см. на рис. 1.
После открытия отверстия между секциями в ранние сумерки (при освещённости 2−10 лк) рыбы сохраняли своё положение в непроточной секции и не совершали попыток приблизиться к выходу в проточную секцию (рис. 2а, 3). Когда освещённость снижалась до ~1 лк, отдельные особи бросковыми движениями подходили к отверстию, но так же быстро возвращались к стенкам. При последующем снижении освещённости до 0.5 лк рыбы начинали чаще приближаться к выходу, а единичные особи быстро выскакивали в проточную секцию (рис. 2б, 3). С наступлением темноты (< 0.1 лк) в течение 1 ч более половины особей поодиночке покидали непроточную секцию. Ко времени окончания эксперимента в стартовой секции оставались три−четыре рыбы. При временном увеличении ночной освещённости до десятых долей люкса (с восходом луны) часть рыб (10–30%) возвращалась в непроточную секцию. Значительная вариабельность минимальных значений числа рыб в проточной секции на диаграмме размаха (рис. 3) отражает временные уходы рыб в непроточную секцию. При заходе луны за кроны деревьев и снижении освещённости ушедшие обратно рыбы возвращались в проточную камеру.
Рис. 3.
Изменение числа рыб (—) в проточной секции установки с изменением освещённости (-
- -): (⚫) и (◼) – средние значения, (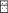 ) – 25–75% и медиана, (
) – 25–75% и медиана, ( ) – минимальное и максимальное значения (без выбросов).
) – минимальное и максимальное значения (без выбросов).
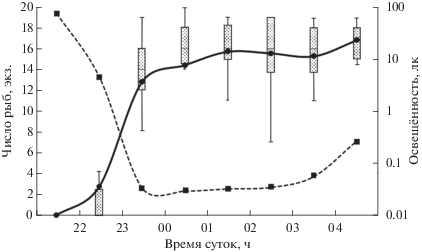
Молодь горбуши, вышедшая в проточную секцию, демонстрировала устойчивый положительный тип реореакции и располагалась на бо́льших скоростях течения (у противоположной от отверстия стенки секции) ближе к входу потока воды в установку (рис. 2в). Оставшиеся в стартовой секции единичные особи находились вдали от выхода в проточную секцию и почти не проявляли активности. После рассвета и закрытия выходного отверстия (освещённость 1−5 лк) рыбы в проточной секции сохраняли своё положение в потоке. Но при помещении на дно секции предмета, имитирующего укрытие (камня диаметром 10 см, взятого с речного дна), рыбы во время манипуляций отплывали к стенкам секции, а после их окончания стремились спрятаться под камень.
Визуальные наблюдения в реке и контрольные ловы ихтиопланктонной конусной сетью показали, что в светлое время суток в толще прозрачной воды молодь горбуши отсутствует. Все рыбы распределяются на дне реки под камнями, подмытыми корнями и в иных укрытиях (рис. 4а). Переворачивание камней позволяет обнаружить эту молодь. Заставить её выйти из укрытий в дневное время, передвигая и приподнимая камни, крайне затруднительно. Время от времени единичные особи поднимаются в толщу воды, но быстро (в течение 1–2 с) снова забиваются под камни. С наступлением глубоких сумерек (< 2 лк) горбуша начинает покидать укрытия и образовывать скопления в слабопроточном прибрежье или в гидравлической тени за крупными препятствиями со скоростью течения 0.01–0.10 м/с на небольшой глубине 0.1–0.3 м (рис. 4б). Наиболее массовые скопления отмечаются у крупных погружённых в воду предметов (коряг, затопленных кочек, пластов дёрна) или в небольших заводях. При испуге так же, как мы наблюдали в экспериментальной установке, молодь горбуши объединяется в плотные стаи. По мере дальнейшего снижения освещённости скопления в прибрежье рассредотачиваются и исчезают.


ОБСУЖДЕНИЕ
Результаты экспериментов показали, что молодь горбуши активно выходит в поток для последующей пассивной миграции. Механизм этого процесса заключается в реализации комплекса врождённых поведенческих реакций в ответ на изменения условий среды. Он состоит из нескольких этапов: выход из укрытий при снижении освещённости вечером и формирование скоплений в прибрежье; перемещение в реоградиенте в условиях снижающейся освещённости; вовлечение в поток и пассивный скат; прекращение миграции при повышении освещённости в утренние часы или её приостановка в течение ночи. Рассмотрим последовательно эти этапы.
Днём при высокой освещённости молодь горбуши избегает находиться в толще воды и распределяется на дне реки, прячась под камнями или в иных укрытиях. Этому способствует отрицательная фотореакция – неотъемлемый поведенческий маркер процесса смолтификации лососёвых (Смирнов, 1975; Hoar, 1976; Folmar, Dickhoff, 1980). В отличие от других видов тихоокеанских лососей горбуша смолтифицируется уже в нерестовых буграх, незадолго до выхода из грунта, на что указывают характерная для смолтов пелагическая окраска покатной молоди и её физиолого-биохимические показатели (Зуева, 1995). Во время нахождения в укрытиях у молоди не проявляется реореакция и, видимо, реализуется положительная тигмореакция.
В вечерние сумерки при снижении освещённости от 2.0 до 1.0–0.5 лк у молоди горбуши возрастает двигательная активность, она выходит из укрытий и формирует скопления в прибрежье. Эти скопления мы наблюдали незадолго до появления первых покатников (Павлов и др., 2015; Кириллов и др., 2018). В условиях эксперимента при близких значениях освещённости рыбы также начинали проявлять повышенную двигательную активность, а отдельные особи выходили в поток. В период выхода из грунта у молоди горбуши прекращают проявляться положительная тигмореакция и отрицательная фотореакция, пороговая величина последней составляет 0.5–1.0 лк (Hoar et al., 1957). При этом возросшая двигательная активность приводит к перемещению молоди на участки, где присутствует слабое течение, вызывающее положительную реореакцию. Пороговая скорость потока для проявления положительной реореакции молодью горбуши составляет 0.03 м/с (Hoar, 1951). Также в этот период у рыб происходят ретиномоторная реакция в сетчатке глаза и темновая адаптация зрения. По экспериментальным данным (Ali, Hoar, 1959), этот процесс идёт при снижении освещённости с 10.0 до 0.1 лк, и его продолжительность составляет ~45 мин. Пороговые величины освещённости для проявления оптомоторной реакции у ранней молоди костистых рыб в зависимости от их размера составляют десятые–сотые доли люкса (Павлов, 1970, 1979). Поэтому при предпороговом уровне освещённости рыбы сохраняют ориентацию в потоке воды и проявляют реореакцию, что мы и наблюдали в экспериментальной установке.
При дальнейшем снижении освещённости (<0.5 лк) отмечается резкое увеличение интенсивности покатной миграции горбуши в реке (Павлов и др., 2015; Кириллов и др., 2018), что указывает на выход в транзитный поток всё бо́льшего числа рыб в этот период. Этот выход осуществляется за счёт избирания рыбами участков с более высокими скоростями течения и активного направленного движения в зону наибольшей скорости. В эксперименте рыбы, воспринимая, по-видимому, турбулентные завихрения в отверстии, совершали бросковые движения в его направлении, а затем также бросковым движением выходили в проточную секцию (рис. 2б, 3). В проточной секции они двигались по вектору скоростей течения от меньших скоростей к бо́льшим. Всё это подтверждает направленный выход в поток. Распределение и положение рыб в проточной секции экспериментальной установки (рис. 2в) подтверждает наличие у них положительной реореакции и выбор ими зоны наибольших скоростей.
По-видимому, сходным образом осуществляется выход молоди горбуши в транзитный поток в естественных условиях в реке. В период снижения освещённости рыбы выходят из укрытий и прибрежья и активно перемещаются в реоградиенте речного потока в направлении более высоких скоростей течения. Когда зрительная ориентация молоди становится невозможна (при освещённости ниже пороговой для оптомоторной реакции – <0.1 лк), реореакция нейтрализуется и молодь подхватывается течением (рис. 4в). Критическая скорость течения, при которой происходит нейтрализация реореакции и снос рыб потоком, находится в обратной зависимости от освещённости (Павлов, 1970, 1979). Направленное перемещение в зону потока, где скорость течения превышает критическую, – универсальный механизм нейтрализации реореакции и начала пассивной покатной миграции у молоди рыб (Pavlov, 1994; Павлов и др., 2007; Pavlov et al., 2008). Дополнительным фактором, способствующим нейтрализации реореакции, выступает турбулентность потока: при её увеличении происходит снижение критической скорости течения для рыб (Штаф и др., 1983).
Концентрация покатников в стрежневой части потока (Каев, 2010; Каев, Игнатьев, 2015; Павлов и др., 2015; Кириллов и др., 2018) подтверждает наличие избирательности в отношении скорости течения. В прибрежной части русла с невысокой скоростью течения молодь горбуши скатывается единично либо вовсе отсутствует (Павлов и др., 2015; Кириллова, Кириллов, 2017; Кириллов и др. 2018). Ранее на карповых рыбах было экспериментально показано, что покатная молодь в реоградиентных условиях концентрируется в зоне наибольших скоростей (Павлов, Штаф, 1981; Павлов, Тюрюков, 1986). Следует отметить, что термин пассивная миграция отражает только форму перемещения. Однако это не означает, что рыбы полностью пассивны и не проявляют никаких реакций. Их распределение резко отличается от распределения мёртвых рыб и поплавков с нейтральной плавучестью (Павлов и др., 2007). Покатники поддерживают дорсовентральное положение тела, проявляют фотореакцию и компенсаторную гидростатическую реакцию, реагируют на изменение скорости течения (Павлов, Штаф, 1981) и на градиент скоростей (Павлов, 1979), на турбулентность потока (Павлов, Лупандин, 1998) и на различного рода гидродинамические стимулы (Павлов, Тюрюков, 1986). В последних случаях молодь на некоторое время даже может восстанавливать свою ориентацию против течения и проявлять реореакцию.
При прекращении пассивной покатной миграции с утренним повышением освещённости и выходе из потока ключевую роль играет восстановление реореакции и её ориентационной компоненты – оптомоторной реакции. Мигрирующая молодь начинает направленно двигаться к укрытиям. В этот период наблюдается утренний спад интенсивности покатной миграции (Павлов и др., 2015; Кириллов и др., 2018). Проявление отрицательной фотореакции и положительной тигмореакции интенсифицируют процесс ухода рыб в укрытия. В экспериментальной установке при помещении в неё камня молодь горбуши забивается под него. Восстановление реореакции мигрирующей молоди горбуши при попадании в ярко освещённый обширный участок русла и её последующие погружение на дно реки и укрытие в грунте среди камней описаны Нивом (Neave, 1955). Отмеченная последовательность завершения пассивного сноса и перемещения в укрытия согласуется с суточной динамикой миграции молоди горбуши – плавное снижение интенсивности и резкое завершение (Павлов и др., 2010, 2015; Кириллов и др., 2018). Как отмечено выше, в светлое время суток горбуша практически не покидает укрытий. Также мы неоднократно наблюдали в течение одной ночи приостановку миграции при появлении луны в просветах облаков или значительную интенсификацию ската при уходе луны с небосклона. Интенсивность покатной миграции молоди горбуши в течение ночи может увеличиваться в два раза после захода луны за горизонт (Павлов и др., 2015). Уходы части молоди в непроточную секцию во время повышения естественной освещённости подтверждают возможность снижения интенсивности ската горбуши. Покатная миграция молоди горбуши проходит в приповерхностном слое воды, где освещённость выше, чем в толще и у дна (Neave, 1955; наши данные). Соответственно, мигрирующие рыбы имеют возможность быстро реагировать на изменение естественной освещённости.
На основе полученных результатов, а также с учётом данных литературы можно предложить следующую схему реализации молодью горбуши покатной миграции. Пассивная покатная миграция у горбуши начинается в вечерние сумерки благодаря выходу молоди из укрытий и направленному активному перемещению в русловой поток. Начало и завершение покатной миграции регулируется комплексом врождённых поведенческих реакций – реореакцией, оптомоторной реакцией, фото- и тигмореакцией. Отрицательная фотореакция в совокупности с положительной тигмореакцией определяют нахождение молоди в светлое время суток (при освещённости >2 лк) в укрытиях. Активный массовый выход молоди из укрытий происходит в вечерние сумерки при освещённости <1 лк, когда она перестаёт избегать света. Реореакция определяет ориентацию рыб против течения вплоть до снижения освещённости ниже пороговой для зрительного механизма ориентации в потоке – оптомоторной реакции. При предпороговых значениях освещённости наблюдается увеличение двигательной активности рыб и их векторизованное движение в реоградиенте в направлении более высоких скоростей течения – вплоть до нейтрализации реореакции и сноса потоком. После активного выхода в транзитный поток молодь распределяется в его стрежневой части, где отмечается её наибольшая концентрация. Процесс перераспределения покатников в речном потоке определяется как его структурой, так и наличием активных движений, совершаемых рыбами для поддержания своего положения в реоградиенте. Во время утреннего повышения освещённости восстанавливается положительная реореакция и начинает проявляться отрицательная фотореакция. В это время мигрирующая молодь активно двигается к укрытиям и благодаря положительной тигмореакции прячется в них.
Список литературы
Бакштанский Э.Л. 1970. Скат молоди горбуши и кеты и причина его задержки в реках Кольского полуострова // Тр. ВНИРО. Т. 74. С. 129−143.
Бакштанский Э.Л. 1977. Изучение поведения покатников лососей в связи с проблемой воспроизводства // Управление поведением животных. М.: Наука. С. 17−19.
Гриценко О.Ф. 2002. Проходные рыбы острова Сахалин (систематика, экология, промысел). М.: Изд-во ВНИРО, 248 с.
Зуева К.Д. 1995. Наличие процесса смолтификации у горбуши Oncorhynchus gorbuscha (Walb.) при отсутствии стадии “parr” // Вопр. ихтиологии. Т. 5. № 2. С. 324–330.
Каев А.М. 2010. Методические аспекты количественного учета покатной молоди лососей в реках Сахалино-Курильского региона // Изв. ТИНРО. Т. 162. С. 194–206.
Каев А.М., Игнатьев Ю.И. 2015. О распределении покатной молоди горбуши в потоке небольшой сахалинской реки // Там же. Т. 180. С. 93–98. https://doi.org/10.26428/1606-9919-2015-180-93-98
Камышная M.С. 1974. Динамика ската и питание молоди горбуши в реках района акклиматизации // Биология промысловых рыб и беспозвоночных на ранних стадиях развития (в связи с вопросами динамики численности). Мурманск: Изд-во ПИНРО. С. 99−100.
Карпенко В.И. 1998. Ранний морской период жизни тихоокеанских лососей. М.: Изд-во ВНИРО, 165 с.
Кириллов П.И., Кириллова Е.А., Павлов Д.С. 2018. Закономерности покатной миграции молоди горбуши Oncorhynchus gorbuscha в р. Малая Хузи (Сахалинская область) // Вопр. ихтиологии. Т. 58. № 6. С. 710–723. https://doi.org/10.1134/S0042875218060139
Кириллова Е.А., Кириллов П.И. 2017. Особенности экологии и поведения молоди горбуши Oncorhynchus gorbuscha в период ската в некоторых реках острова Сахалин // Чтения памяти В.Я. Леванидова. Вып. 7. С. 67–75.
Кухлинг Х. 1985. Справочник по физике. М.: Мир, 520 с.
Павлов Д.С. 1970. Оптомоторная реакция и особенности ориентации рыб в потоке воды. М.: Наука, 148 с.
Павлов Д.С. 1979. Биологические основы управления поведением рыб в потоке воды. М.: Наука, 319 с.
Павлов Д.С., Лупандин А.И. 1998. Влияние освещённости на отношение личинок плотвы (Rutilus rutilus) к потокам с различной турбулентностью // Докл. РАН. Т. 358. № 5. С. 710–712.
Павлов Д.С., Тюрюков С.Н. 1986. Гидродинамический механизм ориентации рыб в потоке воды // Рыбоводство. № 1. С. 32–33.
Павлов Д.С., Штаф Л.Г. 1981. Распределение покатной молоди рыб в реоградиентном потоке // ДАН СССР. Т. 260. № 2. С.509–512.
Павлов Д.С., Нездолий В.К., Ходоревская Р.П. и др. 1981. Покатная миграция молоди рыб в реках Волга и Или. М.: Наука, 320 с.
Павлов Д.С., Лупандин А.И., Костин В.В. 2007. Механизмы покатной миграции молоди речных рыб. М.: Наука, 213 с.
Павлов Д.С., Кириллова Е.А., Кириллов П.И. 2010. Покатная миграция молоди лососевых рыб в р. Утхолок и ее притоках (северо-западная Камчатка). 1. Покатная миграция молоди первого года жизни // Изв. ТИНРО. Т. 163. С. 3–44.
Павлов Д.С., Кириллова Е.А., Кириллов П.И. 2011. Покатная миграция молоди лососевых рыб в р. Утхолок и её притоках (северо-западная Камчатка). 2. Покатная миграция молоди второго и последующих лет жизни // Там же. Т. 164. С. 27–73.
Павлов Д.С., Кириллов П.И., Кириллова Е.А., Черешкевич Ф.Г. 2015. Покатная миграция молоди горбуши Oncorhynchus gorbuscha (Walbaum) в реке Малая Хузи (северо-восток острова Сахалин) // Биология внутр. вод. № 4. С. 64–75. https://doi.org/10.7868/S0320965215040129
Руководство по применению анестетика “гвоздичное масло” в аквакультуре. 2011. М.: Изд-во ВНИРО, 58 с.
Смирнов А.И. 1975. Биология, размножение и развитие тихоокеанских лососей. М.: Изд-во МГУ, 336 с.
Солдатов В.К. 1912. Исследование биологии лососевых Амура // Рыбные промыслы Дальнего Востока. СПб.: Тип. В.О. Киршбаума, 223 с.
Штаф Л.Г, Павлов Д.С., Скоробогатов М.А., Барекян А.Ш. 1983. Влияние степени турбулизации потока на поведение рыб // Вопр. ихтиологии. Т. 23. Вып. 2. С. 307–317.
Шунтов В.П., Темных О.С. 2008. Тихоокеанские лососи в морских и океанических экосистемах. Т. 1. Владивосток: Изд-во ТИНРО-центр, 481 с.
Ali M.A., Hoar W.S. 1959. Retinal responses of pink salmon associated with its downstream migration // Nature. V. 184. № 4680. P. 106–107. https://doi.org/10.1038/184106a0
Folmar L.C., Dickhoff W.W. 1980. The parr-smolt transformation (smoltification) and seawater adaptation in salmonids a review of selected literature // Aquaculture. V. 21. № 1. P. 1−37. https://doi.org/10.1016/0044-8486(80)90123-4
Godin J.-G.J. 1980. Temporal aspects of juvenile pink salmon (Oncorhynchus gorbuscha Walbaum) emergence from a simulated gravel redd // Can. J. Zool. V. 58. P. 735–744. https://doi.org/10.1139/z80-105
Groot C., Margolis L. 1991. Pacific salmon life histories. Vancouver: Univ. Brit. Columbia Press, 564 p. https://doi.org/10.1016/0044-8486(93)90177-z
Hoar W.S. 1951. The behaviour of chum, pink and coho salmon in relation to their seaward migration // J. Fish. Res. Board Can. V. 8. P. 241–263. https://doi.org/10.1139/f50-015
Hoar W.S. 1976. Smolt transformation: evolution, behaviour and physiology // Ibid. V. 33. P. 1234–1252. https://doi.org/10.1139/f76-158
Hoar W.S., Keenleyside M.H.A., Goodall R.G. 1957. Reactions of juvenile Pacific salmon to light // Ibid. V. 14. P. 815–830. https://doi.org/10.1139/f57-036
Keene J.L., Noakes D.L., Moccia R.D., Soto C.G. 1998. The efficacy of clove oil as an anaesthetic for rainbow trout, Oncorhynchus mykiss (Walbaum) // Aquacult. Res. V. 9. № 2. P. 89–101. https://doi.org/10.1046/j.1365-2109.1998.00927.x
McDonald J. 1960. The behaviour of Pacific salmon fry during their downstream migration to freshwater and saltwater nursery areas // J. Fish. Res. Board Can. V. 17. P. 665–685. https://doi.org/10.1139/f60-051
Neave F. 1955. Notes on the seaward migration of pink and chum salmon fry // Ibid. V. 12. P. 369–374. https://doi.org/10.1139/f55-022
Pavlov D.S. 1994. The downstream migration of young fishes in river (mechanisms and distribution) // Folia Zool. V. 43. № 3. P. 193–208.
Pavlov D.S., Kostin V.V., Lupandin A.I. 1997. Development of downstream migration in roach larvae (Rutilus rutilus L.): the role of locomotor activity // Rus. J. Aquat. Ecol. V. 7. № 1. P. 75–84.
Pavlov D.S., Mikheev V.N., Lupandin A.I., Skorobogatov M.A. 2008. Ecological and behavioural influences on juvenile fish migrations in regulated rivers: a review of experimental and field studies // Hydrobiologia. № 609. P. 125–138. https://doi.org/10.1007/s10750-008-9396-y
Pritchard A.L. 1944. Physical characteristics and behaviour of pink salmon fry at McClinton Creek. B.C. // J. Fish. Res. Board Can. V. 6. P. 217–227. https://doi.org/10.1139/f42-026
Quinn T.P. 2005. The behavior and ecology of pacific salmon and trout. Seatle; Bethesda, Maryland: Univ. Wash. Press; Amer. Fish. Soc., 378 p.
Schlyter P., Sweden S. 2006. Radiometry and photometry in astronomy (http://stjarnhimlen.se/comp/radfaq.html#10).
Дополнительные материалы отсутствуют.
Инструменты
Вопросы ихтиологии